
Nagdy F. Abdel-Baky
Department of Crop Production and Protection, College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah 51452, Kingdom of Saudi Arabia
M.A. Al-Deghairi
Department of Crop Production and Protection, College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah 51452, Kingdom of Saudi Arabia
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 6 | Page No.: 356-368







ABSTRACT
Impact of the host plant type on certain biological aspects and parasitism level of E. mundus under laboratory and semi-field conditions was evaluated. The parasitoid biological aspects were greatly differed within the host type. Parasitoid life cycle was shorter on squash, followed by common beans and sweet pepper, which lasted 27.6±1.9, 25.9±1.3 and 23.7±1.1 days, respectively. In contrary, female longevity was shorter on the sweet pepper (9.9±1.6 days), followed by the common bean (10.8±1.1 days) and was longer on squash (11.7±1.3 days). Additionally, E. mundus life span and female fecundity were also studied and varied among the studied hosts. Effect of host plants on both colonization of pest nymphs and parasitism percentages, were also evaluated. Greater numbers of young and old nymphs and higher parasitism rates were observed on squash followed by common beans, whereas, sweet pepper was last in this respect. Subsequently, the reproduction and biological characteristics of E. mundus have been shown obviously to be influenced by host plant. Efficiency of releasing parasitoids was greatly affected by host plant type as well as releasing rates. Releasing the parasitoid with constant numbers against various population densities of the pest achieved different pest control levels. Releasing rates of 1:5 and 1:10 (parasitoid:pest) gave good control measures, whereas, moderated control levels were fulfilled with ratios 1:20 and 1:30. Increasing pest densities negatively correlated with good control measures.
PDF Abstract XML References Citation
How to cite this article
Nagdy F. Abdel-Baky and M.A. Al-Deghairi, 2008. Role of Host Plants on the Biological Aspects and Parasitism Levels
of Eretmocerus mundus Mercet (Hymenoptera: Aphelinidae), a Parasitoid
of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Journal of Entomology, 5: 356-368.
DOI: 10.3923/je.2008.356.368
URL: https://scialert.net/abstract/?doi=je.2008.356.368
DOI: 10.3923/je.2008.356.368
URL: https://scialert.net/abstract/?doi=je.2008.356.368
INTRODUCTION
Currently, Bemisia tabaci species complex is considered to be one of the worst world`s top 100 invasive species, because of its serious damage to agricultural production and associated industries. Chemical control of Bemisia species complex provides only short-term solutions and the insects has provoked the development of resistance to those chemicals (Dittrich et al., 1990; Damásio et al., 2007). So far, several biological control strategies have been involved and evaluated for the pest management, as the use of hymenopteran parasitoids, either native or exotic (Goolsby et al., 1998; Ardeh et al., 2005) and could be an attractive management alternative for whiteflies (Gerling et al., 2001). At least 13 species B. tabaci parasitoids in the genus Eretmocerus have been identified from the New World (Zolnerowich and Rose, 1998) and many are important in biological control of B. tabaci (Gerling et al., 2001). The Eretmocerus spp., with at least two species (E. eremicus Rose and Zolnerowich and E. mundus Mercet) are commercially available (Zolnerowich and Rose, 1998; Hoddle and Van Driesche, 1999).
Whitefly parasitoids formed one of the important backbone of IPM programs. When IPM was practiced, E. mundus was found as the most predominant parasitoid species against whiteflies, particularly in the open field (Rodriguez-Rrodrigez et al., 1994). Biological characteristics of E. mundus have bean also studied under laboratory conditions (Sarhan 1976; Abdel-Baky and Ragab, 2005).
Eretmocerus mundus Mercet (Hymenoptera: Aphelinidae) has been recorded in many countries in the Mediterranean Basin (Mound and Halsey, 1978) and world-wide making it the most important, abundant and indigenous parasitoid attacking B. tabaci, a serious pest in greenhouses and outdoor crops (Roderiguez-Rrodrigez et al., 1994). E. mundus is standing out among whitefly potential biological control agents and considered as highly promising biological control agent of whiteflies (Goolsby et al., 1998). This may be due to many scientific facts such as; (a) no known Eretmocerus species exhibits auto-parasitism: a potentially deleterious trait to biological control (Van Lenteren et al., 1997) and b: its reproductive and competitive potential on its host (Greenberg et al., 2002; Urbaneja et al., 2007). E. mundus parasitizes all four nymphal instars of Bemisia tabaci, with 2nd instars being the preferred stage. E. mundus is a primary parasitoid that oviposites underneath the nymphal whitefly host (Gerling and Fried, 2000). To our knowledge, two Eretmocerus types used on the international scale, one species is arrhenotokous, but the other is thelytokous type, which known as the Australian strain (McAuslane and Nguyen, 1996). De Barro et al. (2000) considered thelytokous populations as the best candidates for biological control of whiteflies world-wide.
The suitability of whitefly nymphs as a host was recognized (Jones and Greenberg, 1998). Jervis et al. (2001) recognized two alternative life-history strategies in parasitoid life in respect of development mode. Kionobiont expresses when 2nd or 3rd instars parasitized by E. mundus, in which the parasitized larva continues to feed, grow and complete its development. Whereas, idiobiont expresses when B. tabaci 4th instars used as a parasitoid`s host, in which the parasitized larva evidently stop development and no life cycle occurred.
Releasing rates for natural enemies in greenhouses are generally expressed in numbers per unit area. Releasing rates of E. mundus were initially set to those recommended for E. eremicus of 1.5-3 wasps/m2/week as a preventative treatment and 3-9 wasps/m2/week as curative treatments (Stansly et al., 2005). These rates largely contrasted with those tested for control of B. tabaci by Hoddle et al. (2001) and Van Driesche et al. (2001).
Therefore, the objectives of the present study are to evaluate the impact of host plant type on some biological parameters of E. mundus and determine its potentiality as a bio-control agent for whitefly. All these measures were designed with objective to identify optimal release rates for whitefly control.
MATERIALS AND METHODS
Whiteflies and Parasitoids
Separated cultures from B. tabaci and its parasitoid E. mundus were originally initiated from individuals collected from squash, cucumber and pepper plants insecticides free at Qassim region, KSA. Whitefly colony was kept on common bean (Phaseolus vulgaris L.) under laboratory conditions at the room temperature (30±2°C) and photoperiod 12:12 (L:D). At least three generations of B. tabaci was gained under these conditions, to serve as host of the parasitoids. In respect to the parasitoid`s colony, nymphs of B. tabaci with recognizable parasitoid`s pupae and/or had a parasitization symptoms were removed and stored out at 25°C and kept until adult emergence and then collected in small tubes.
Three plant hosts namely; squash Cucurbita pepo L. (Cucurbitaceae), sweet pepper, Capsicum anuum L. (Solanaceae) and common bean, Phaseolus vulgaris (Fabaceae) planted into 5.4 cm2 peat moss. Plants were trimmed to 2 or 3 stems supported by strings attached to an overhead wire. These plant hosts served as a whitefly host to evaluate their influence on the biological aspects of E. mundus and its efficiency as a bio-control agent against B. tabaci under laboratory conditions.
Effect of the Plant Type on the Biological Aspects of E. mundus
Fifteen pots of each tested host plants were grown in screened cages until reaching a good vegetative development and free of whitefly infestation at 27±1°C and 72±2% RH. For each host plant, three full expanding leaves were selected from each pot, a couple of B. tabaci adults were introduced to each leaf under a clip cage and left for 24 h, then adults were removed. The numbers of deposited eggs were counted on each leaf of the three host plants. Since E. mundus prefers to parasitize on the 1st or 2nd instars of B. tabaci (Jones and Greenberg, 1998), once B. tabaci nymphs reached to the 2nd, the desire number of nymphs was left on the leaves and then one couple of E. mundus was introduced and kept for only 24 h. Whitefly infested leaves along with its parasitoids were left until emergence of whitefly adults or parasitoids. The life cycle, life span and female fecundity and longevity were recorded.
Parasitoid Longevity
Longevity of E. mundus, which fed on honey solution, was determined in air-conditioning controlling cabinet at 27±1°C, 72±2 RH and photoperiod of 12:12 (L:D). Fifty newly emerged parasitoids were individually placed in Petri-dishes (9 cm in diameter) with screened lids. The parasitoid`s adults were re-provisioned daily with honey smeared on the screened lids until they died. Adult longevity was determined.
Parasitoid Fecundity
Newly emerged females of E. mundus were singly isolated in glass chimney with the top sealed with screen cover. Common beans pots with leaflets infested by the 2nd instars (50-80 individuals) of B. tabaci were inserted inside the glass chimney. Bean plants were daily replaced with new similar pots until the death or escape of the parasitoid. Fecundity of E. mundus females were assessed by counting nymphs showing sign of parasitism and/or counting the adult numbers of the emerged parasitoid.
Efficiency of Eretmocerus mundus in Controlling Bemisia tabaci under Laboratory Conditions
Evaluations the Role of Host Plant Type on the Efficiency of E. mundus as a Bio-Control Agent
Impact of squash, common beans and sweet pepper on the efficiency of the parasitoid in controlling B. tabaci was evaluated. The evaluation was conducted as replicated releases of the parasitoids into semi-field cages. Cages were placed at the same time when the three host plants were planted and grown in small plastic pots. Each experiment was replicated five times with the total of 200 plants of each host plant. In the respect of each host plant, cage frames of 1.5 mL long on each side, were constructed of plastic PVC pipe and three-corner joints, with one cross joint in the middle of cage top. Cage covers were mesh screened with openings small enough to prevent passage by whitefly and parasitoids. Also, the cages were provided by zippers sewn in one side of the cage for easy access. Each cage spanned forty plastic pots cultivated by the plants. When the plants reached 20 cm height with at least 5 fully expanded leaves, B. tabaci adults were inoculated by introducing 100 pairs per cage and left for three weeks to reach its maximum population level. Average of whitefly populations/plant/cage/host was counted. Subsequently, 50 pairs of E. mundus parasitoid were introduced and let for two weeks before evaluating its role. Weekly random samples were taken by counting the whitefly populations and its parasitoids, thereon, the parasitism percentages were calculated. Immature parasitoids inside the pupae or older larvae of the whitefly host could be readily detected, so leaf samples were collected at that point reliably measured the parasitoid progeny. Leaves in cages were weekly inspected by taken 20 leaves in random and taken to the laboratory to record the number of whiteflies and parasitoids. Leaves were examined in the laboratory with stereoscopic microscopes to record the production of the parasitoid progeny for each host plant. The parasitized nymphs were counted on all harvested leaves; this included those individuals with displaced (asymmetrical) mycetomes that were indicative of the presence of mid-sized parasitoid larvae which indicated large, late instars nymphs, pupae of E. mundus and whitefly mummies with parasitoid emergence holes. Accordingly, comparisons among host plants were also involved.
Effect of Releasing Rates on the Parasitoid Efficiency
In this experiment, plastic pots cultivated with squash plants used as a whitefly host. Ten pairs of B. tabaci adults were introduced and kept under the screened cages for 48 h, then removed. The plants with B. tabaci eggs were incubated at 27±1°C until whitefly nymphs reached the desired age for parasitism (2nd or 3rd instars). Constant numbers of B. tabaci nymphs (200 individuals/plant) were determined. There were 15 replicates for each releasing ratio, each was covered with screened cage. After that, the parasitoid was released at six release rates (parasitoid:pest): 1:5, 1:10, 1:20, 1:30, 1:40 and 1:50/plant as well as, the check treatment with 0-parasitiod:200 B. tabaci nymphs. Care was taken by inspecting control cages first, followed by low release rate cages and finally high release rate cages to minimize risk of cross contamination by parasitoids.
Evaluation
Plants were weekly monitored for 10 successive weeks. In each cage, Plant leaves, infested with B. tabaci nymphs and its parasitoids at each release rate, were investigated. All whitefly stages and parasitized instars were counted on each leaf using a 10x-hand lens. Relative incidence of parasitized whiteflies was expressed as the number of parasitized nymphs/total number of nymphs/plant. Additionally, emerged whitefly adults were counted/plant of each release rate.
In all cases, healthy nymphal instars, exuviae from parasitized and unparasitized whiteflies and parasitized nymphs were counted. Early stages of parasitism in nymphal instars were recognized by displaced mycetomes and later stages by the presence of the parasitoid`s pupa.
Data Analysis
Obtained data were subjected to one-way analysis of variance with mean separation using LSD test in the event of a significant F (p<0.05) (Duncan, 1951). Degree of whitefly populations suppression obtained by parasitoid treatments was expressed using the formula 100x(1-treated/control).
RESULTS
Biological Aspects of E. mundus and Role of Host Plant Type
Eretmocerus mundus biological aspects were greatly differed based on the plant host type. The results in Table 1, indicated that faster development, shorter life cycle and higher female progeny were obviously observed with cucumber plants, followed by common bean, while the sweet pepper came in last. The life cycle of the parasitoid was shorter when reared on squash than on other two hosts. The life cycle lasted 27.6±1.9, 25.9±1.3 and 23.7±1.1 days on sweet pepper, common bean and squash plants, respectively. Female longevity was shorter when developed on sweet pepper which lived 9.9±1.6 days and increased on common bean (10.8±1.1), while, it was longer on the squash and recorded 11.7±1.3 days. Both of parasitoid life cycle and female longevity were significantly varied (p≥0.5) as shown in Table 1.
| Table 1: | Effect of host plants on certain biological characters of Eretmocerus mundus |
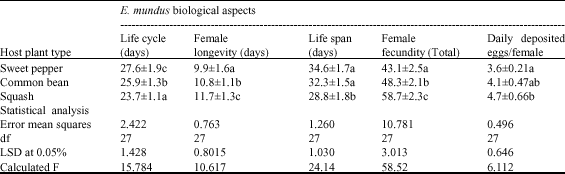 | |
| Means followed by the same letter within a column are not significantly different at (0.05%) (Duncan Multiple Rang test) | |
| Table 2: | Population of B. tabaci nymphs, colonization percentages, number of parasitized nymphs and emerged parasitoid of E. mundus |
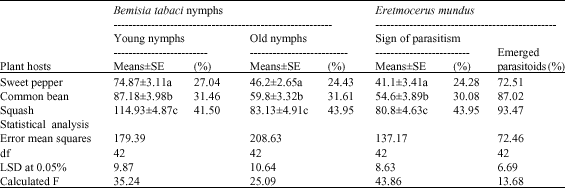 | |
| Means followed by the same letter within a column are not significantly different at (0.05%) (Duncan Multiple Rang test) | |
Life span of E. mundus also varied among the studied hosts, which recorded 34.6±1.7, 32.3±1.5 and 28.8±1.8 days on sweet pepper, common bean and squash, respectively. This means that the parasitoid life span was shorter on squash than on other two hosts. Statistical analysis showed no significant difference in parasitoid life span when reared on sweet pepper and common beans, but it significantly varied on squash plant.
Regarding female fecundity, host plant type showed significant variation in this respect when the parasitoid reared on three different hosts (Table 1). The daily deposited eggs/female was 3.6±0.21 on sweet pepper, 4.1±0.47 on common beans and 4.7±0.66 on squash. Collectively, parasitoid female laid more eggs on squash (58.7±2.3 eggs/female), followed significantly by common beans (48.3±2.1 eggs/female) and the sweet pepper came in last which recorded only 43.1±2.5 eggs/female (Table 1).
Impact of Host Plants on Both Bemisia tabaci Nymphs and Eretmocerus mundus Parasitism Rates
Bemisia tabaci Nymphs
The type of host plant directly affected whitefly population (as a mean of nymphs/inch2/leaf) and colonization percentages. A great number of whitefly nymphs had been recorded on squash plant, which averaged 114.93±4.87 nymphs/cm2/leaf for young nymphs and 83.95±4.91 nymphs/inch2 of leaf for old nymphs (Table 2). The common beans came next where number of young nymphs reached 87.18±3.98 and 59.8±3.32 nymphs/inch2 for old nymphs. On the other hand, the sweet pepper harbored the lowest population level of B. tabaci nymphs (74.87±3.11 and 46.2±2.65 for young and old nymphs, respectively). Statistically, number of young nymphs (LSD at 0.05% = 9.87; Calculated F = 35.24) and old nymphs significantly varied (LSD at 0.05% = 10.64; Calculated F = 25.09) among the three host plants (Table 2).
The insect pest colonization also varied among plant host types and nymph age. Young nymphs averaged 41.50, 31.46 and 27.04% on squash, common beans and sweet pepper, respectively. The same trend was also obtained with old nymphs, which lasted 43.95% on squash, 31.61% on common beans and 24.43% on sweet pepper as shown in Table 2.
Eretmocerus mundus Populations
To evaluate the impact of host plant on biological aspects of E. mundus two biological parameters were tested (Table 2). These parameters were signs of parasitized nymphs and percentage of emerged parasitoids. Number of B. tabaci that showed signs of parasitism by E. mundus varied based on the plant type. Whitefly nymphs that harbor squash plant were more preferred for parasitism followed by common beans and finally the sweet pepper. Nymphs mean (inch2 of leaf) that showed sign of parasitism was 80.8±4.63, 54.6±3.89 and 41.1±3.41 for squash, common beans and sweet pepper, respectively. The statically analysis revealed that number parasitized B. tabaci nymphs varied significantly (LSD at 0.05% = 8.63; Calculated F = 43.86) among the tested plants (Table 2). Moreover, the results showed that squash supported 43.95% of total parasitized nymphs, followed by common beans (30.08%) and sweet pepper (24.28%).
Additionally, the number of emerged parasitoids was also varied according to plant host. On squash, 93.47% parasitoid adults have emerged from parasitized nymphs, followed by common beans (87.02%) and sweet pepper (72.51%). This means that some plants could support a large number of the parasitoids than others. Therefore, these hosts should be used in rearing the pest and its parasitoid. It could be concluded that the squash as a host was the preferred one for rearing E. mundus followed by common beans and sweet pepper plants came in last.
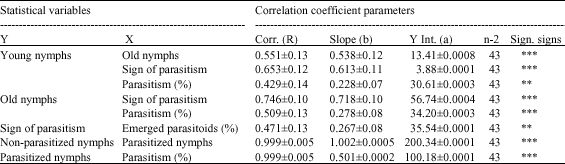
Table 3, clearly shows the correlation coefficient among B. tabaci nymphs and its parasitoid, E. mundus. The relationship between young nymphs and old nymphs of B. tabaci, parasitized nymphs and parasitism percentage was high with R values 0.551±0.13, 0.653±0.12 and 0.429±0.14 for old B. tabaci nymphs, parasitized nymphs and parasitism %, respectively (Table 3). Furthermore, old nymphs of B. tabaci relation with both of parasitized nymphs and parasitism % was also strong and their R values were 0.746±0.10 and 0.509±0.13, respectively. This means that not all young nymphs complete their development and transformed to the 3rd or 4th instars as a result of some mortality factors. The sign of parasitized nymphs was clearly noted on both young and old nymphs, but it was obvious with old nymphs (Table 3). The relation between nymphs with signs of parasitism and emerged parasitoid % was moderate but showed significant correlation and its "R" value was 0.471±0.13. The non-parasitized nymphs had a strong positive linear correlation with the parasitized nymphs (R = 0.999±0.005), as well as, a strong positive linear correlation between the parasitized nymphs and parasitism % (R = 0.999±0.005).
| Table 3: | Correlation coefficient parameters among B. tabaci nymphs and its associated numbers and percentages of E. mundus |
 | |
Evaluation of Eretmocerus mundus Role in Suppressing Bemisia tabaci Populations
Impact of Host Plant Type
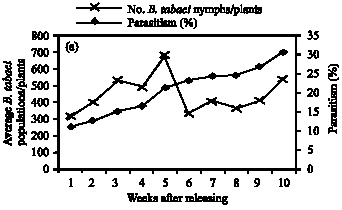
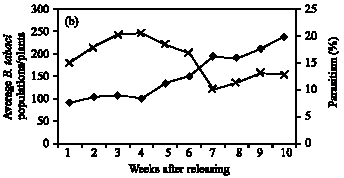
Host plants obviously affected E. mundus behavior in controlling B. tabaci. On squash plants, average number of B. tabaci was 1016.5/plant, but it declined to 448.5/plant after releasing the parasitoid (Fig. 1). This means that the parasitoid was able to reduce pest population by 55.9%. In this study, the parasitism percentages ranged between 11.08 to 30.64% with an average of 20.62% (Fig. 2). On common beans, the average B. tabaci number was 321.4 individuals/plant but it was reduced to 186.1 individuals/plant after releasing the parasitoids. Data revealed that E. mundus reduced the pest populations by 42.1% on the common beans (Fig. 1). The parasitism percentages fluctuated between 7.63 to 19.78% with an average of 12.63% (Fig. 2). Before the release of parasitoid, the average B. tabaci numbers reached 201.9 individuals/plant and listed 106.8 individuals/plant after releasing. The parasitoid caused 32.27% mortalities among B. tabaci population (Fig. 1) and ranged between 4.5 to 16.63 % with an average of 9.97% (Fig. 2) on sweet pepper plants.
On all host plants tested, the pest populations were higher at the beginning of the experiments and gradually reduced after that (Fig. 3a, b, c). These figures illustrated that the developing parasitoid population was lower in comparison with the pest population, but it multiplied and increased as it was higher at the end of experiments. Statistically, efficiency of the parasitoid in controlling the whitefly was significantly differed (p≥0.05) among plant hosts.
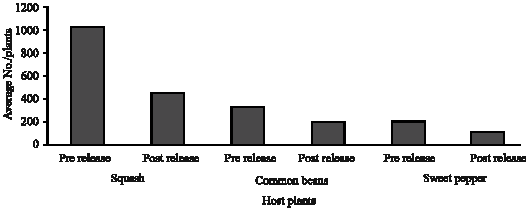 | |
| Fig. 1: | B. tabaci population as an average/plant before and after releasing the parasitiod |
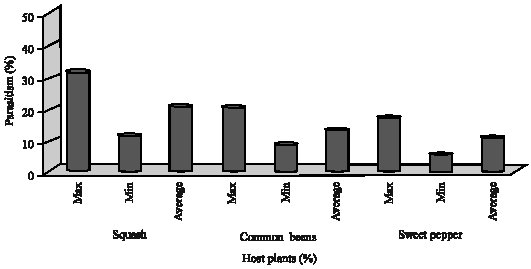 | |
| Fig. 2: | Parasitism percentage of E. mundus a parasitiod of B. tabaci among three plant hosts |
 | |
 | |
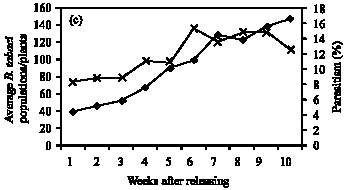 | |
| Fig. 3: | Role of E. mundus in suppression of B. tabaci population (a) squash, (b) common bean and (c) sweet pepper plants |
Impact of Releasing Rates
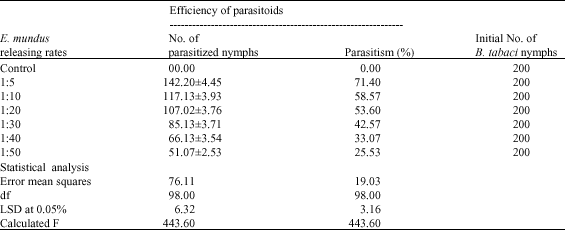
Releasing constant numbers of E. mundus, against varied pest densities, affected the controlling levels (Table 4). Good control levels could be achieved when the parasitoid faces low pest populations. About 71.4% mortalities of B. tabaci population (average of parasitized nymphs = 142.20±4.45/plant) were achieved when the parasitoid was released by one parasitoid against five individuals of the pests. The parasitism percentages were over 50% when releasing one parasitoid against 10 individuals of the pests (58.57%, with an average of 117.13±3.93 parasitized nymphs/plant) and one parasitoid against 20 individuals of the pests (53.6%, with an average of 107.02±3.76 parasitized nymphs/plant). Releasing the parasitoid with ratios of one parasitoid against 30, 40 and 50 individuals of the pests caused 42.57, 33.07, 25.53% mortalities among the pest populations, respectively (Table 4). The average of parasitized nymphs in the pervious releasing ratios was 85.13±3.71, 66.13±3.54 and 51.07±2.53 individuals/plant, respectively.
| Table 4: | Efficiency of E. mundus in controlling B. tabaci when released against different pest densities |
 | |
DISCUSSION
Classical biological control of whiteflies, which depends upon parasitoids, becomes the primary objective and seen as an important component of integrated management (Faust, 1992; Pickett et al., 2004). Controlling a serious pest on different plant hosts with one parasitoid species and even different populations of the same species, often vary as a result of changes in parasitoid characteristics such as host acceptance, fecundity, development time, etc. and these are often highly dependent on the host plant type and the surrounding environmental conditions (Hoelmer and Goolsby, 2002).
The biological characters of E. mundus are affected by many important factors such as an environmental factors, host type, density, size or age and host plant type (Jones et al., 1999). The previous studies indicated that the single female of E. mundus laid approximately 81.1 to 247.5 eggs, during its life time of 10-16 days under laboratory conditions (Gerling and Fried, 2000). The reproduction and biological characteristics of E. mundus have been shown obviously to be influenced by host plant. Previously, Eretmocerus spp. was found to lay more eggs on the glabrous leaves of collard than on the hirsute leaves of eggplant, cucumber and tomato. Host plant morphology, in particular leaf hair density and structure was found to have a major effect on the searching efficiency of parasitoids by slowing or inhibiting its walking speeds and in turn reducing parasitism rates (Van Lentern et al., 1987; Headrick et al., 1996; McAuslane and Nguyen, 1996; Qiu et al., 2005).
However, the parasitoid fecundity, in this study (Table 1), was lower compared with the previous studies. The differences in parasitoid fecundity may be due to its oviposition behavior, which lays its eggs externally beneath nymphs and not within them (Buncker and Jones, 2005). Where in, Foltyn and Gerling (1985) considered E. mundus as an eco-endoparasitiod, oviposition under 2nd and/or 3rd nymphal bodies of whiteflies. Once the egg hatched, the larva burrows through the body. A delay in penetration and/or slower developmental rates after penetration could be responsible for the extended period of development.
The mechanisms of host instar suitability for whitefly parasitoids have not been studied and only can be surmised. Gerling et al. (2001) suggested that young host instars are less able to defend themselves with horizontally protruding wax filaments than the third and fourth instars. The non-penetrating E. mundus larva almost induced permanent developmental arrest in its 4th instar of whitefly host and also caused a reduction in whole body host ecdysteroid titers. Ghahari et al. (2005) studied the biology of thelyotokous biotype of E. mundus as part of an evaluation of its potential for biological control of B. tabaci. They found that the parasitoid deposited more eggs under 2nd and 3rd nymphal instars than 1st or 4th instars. Moreover, when females fed honey, with no access to whitefly nymphs, they lived significantly longer (13.6±4.7 day) than those given access to nymphs with no honey (7.6±2.21 day). In this point, their results were completely in agreement with our finding (Table 1). They concluded that E. mundus longevity on host-infested disks was significantly less than when parasitoids were supplied with honey. Therefore, the net reproductive rates, measured as the number of progeny per female was within the range reported for Eretmocerus spp. or within E. mundus biotypes (Vet and Van Lenteren, 1981; McAuslane and Nguyen, 1996; Ghahari and Ostovan, 2002).
Consequently, Abdel-Baky and Ragab (2005) explained many scenarios that could interprets the lower fecundity in the current study,( 1) the hatching parasitoid larva may be unable to penetrate the nymph cuticle, due to the host defense system or thick wax layers (Tuda and Bonsall, 1999), (2) females of E. mundus may be unable to discriminate between parasitized and non-parasitized nymphs, so, the same female or another one from the same species or from others may lay more than one egg under a parasitized nymphs which leads to inter or intra-specific competitions. In this trend, Sarhan (1976) mentioned that E. mundus female deposited one egg/whitefly nymph and in few cases the parasitoid laid two eggs under the same nymph, but only one parasitoid adult was emerged; (3) females of different species of Eretmocerus share the same host, this may leads to super or multi-parasitism (Ardeh et al., 2005) and the recent study added additional interpretation, (4) the parasitoid adults may attack a non-suitable instar (4th instars) resulted to un-completed life cycle and no parasitoids adults were obtained as in idiobiont concept (Jones and Greenberg, 1998; Jervis et al., 2001; Ghahari et al., 2005). The previous results may have influenced the parasitoids biological characters and its efficiency as a bio-control agent against whiteflies in the open field or in greenhouses.
In respect to the effect of host plant type on the biological characters of E. mundus, Mandour et al. (2007) reported that the parasitoid use the pest honeydew as a contact kairomone to locate its hosts. The sweet potato whitefly produce large quantities of sugar-rich honeydew while feeding on the phloem of their host plants. Plant sap contains amino acids and secondary plant products in small quantities which may affect of parasitoid attractions (Byrne and Miller, 1990; Romeis and Zebitz, 1997). The parasitoid is responded to whitefly honeydew extraction up to 10 days and search time decreased with increasing honeydew age. In Table 1 and 2 higher population of the pest on squash resulted on increasing honeydew secretion and this gave good biological characters and higher parasitism rates on the plants. Additionally, parasitoid response to the host was not only due to honeydew quantity but also due to the enzymatic involved and sugar concentrations and type (Lewis et al., 1998; Salvucci et al., 1997). Hendrix et al. (1992) reported that the Homopteran honeydew sugar composition is determined by both the insect and plant species.
Finally, current results show that E. mundus is a promising candidate bio-agents against whiteflies. The reproductive potential and certain biological aspects of the parasitoid could be increased and give a satisfied control measures when reread on the suitable host plant and insect instars. Further studies should be applied in the future for integrate the parasitoid with other biological control agents to enhance their role in controlling whiteflies.
ACKNOWLEDGMENTS
The authors are very grateful to an anonymous reviewer for most valuable comments improving the manuscript considerably.
REFERENCES
- Ardeh, M.J., P.W. Jony and J.C. Van Lantern, 2005. Intra-and interspecific host discrimination in arrhenotokous and thelytokous Eretmocerus spp. Biol. Cont., 33: 74-80.
CrossRef - Buckner, J.S. and W.A. Jones, 2005. Transfer of methyl-branched hydrocarbons from the parasitoid, Eretmocerus mundus, to silverleaf whitefly nymphs during oviposition. Comparative Biochem. Physiol. A Mol. Integrative Physiol., 140: 59-65.
CrossRef - Byrne, D.M. and W.B. Miller, 1990. Carbohydrate and amino acid composition of phloem sap and honeydew produced by Bemisia tabaci. J. Insect. Physiol., 36: 433-439.
CrossRef - Damásio, J., L. Guilhermino, A.M.V.M. Soares, M.C. Riva and C. Barata, 2007. Biochemical mechanisms of resistance in Daphnia magna exposed to the insecticide fenitrothion. Chemosphere, 70: 74-82.
CrossRef - De Barro, P.J., P.I. Hart and R. Morton, 2000. The biology of two Eretmocerus spp. (Haldeman) and three Encarsia spp. forster and their potential as biological control agents of Bemisia tabaci biotype B in Australia. Entomol. Exp. Applied, 94: 93-102.
CrossRef - Foltyn, S. and D. Gerling, 1985. The parasitoids of aleyrodid Bemisia tabaci in Israel: Development, host preference and discrimination of the aphelinid wasp Eretmocerus mundus. Entomol. Exp. Applied, 38: 255-260.
CrossRef - Gerling, D. and R. Fried, 2000. Biological studies with Eretmocerus mundus Mercet (Hym.: Apheliniae) in Israel. Bulletin OILB/SROP, 23: 117-123.
Direct Link - Gerling, D., O. Alomar and J. Arno, 2001. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot., 20: 779-799.
CrossRef - Ghahari, H. and H. Ostovan, 2002. Preliminary study on the biology of two populations of Eretmocerus mundus Mercet (Hymenoptera: Aphelinidae) on Bemisia argentifolii (Homoptera: Aleyrodidae). Proceedings of the 15th Iranian Plant Protection Congress, September 7-11, 2002, Razi University of Hermanshah, pp: 325.
- Ghahari, H., J. Hunang and Z.H. Wang, 2005. Biology of a thelyotokous biotype of Eretmocerus mundus (Hymenoptera: Aphelinidae) on Bemisia tabaci (Homoptera: Aleyrodidae). Insect. Sci., 12: 461-465.
CrossRefDirect Link - Goolsby, J.A., M.A. Ciomperlik, B.C. Legaspi, J.C. Legaspi and L.E. Wendel, 1998. Laboratory and field evaluation of exotic parasitoids of Bemisia tabaci (Gennadius) (Biotype B) (Homoptera: Aleyrodidae) in the Lower Rio Grande Valley of Texas. Biol. Cont., 12: 127-135.
CrossRef - Greenberg, S.M., W.A. Jones and T. Liu, 2002. Interactions among two species of Eretmocerus (Hymenoptera: Aphelinidae), two species of whiteflies (Homoptera: Aleyrodidae) and tomato. Environ. Entomol., 31: 397-402.
Direct Link - Headrick, D.H., T.S. Bellows and T.M. Perring, 1996. Host plant effects on the behaviors of Eretmocerus sp. nr californicus raised from melon. BioControl, 41: 15-26.
CrossRefDirect Link - Hendrix, D.L., Y. Wei and J.E. Leggett, 1992. Homopteran honeydew sugar composition is determined by both the insect and plant species. Comparative Biochem. Physiol. Part B: Biochem. Mol. Biol., 101: 23-27.
CrossRef - Hoddle, M.S. and R.G. Van Driesche, 1999. Evaluation of Eretmocerus eremicus and Encarsia Formosa (Hymenoptera: Aphelinidae) Beltsvile strain in commercial greenhouses for biological control of Bemisia argentifolii (Homoptera: Aleyrodidae) on colored poinsettia plants. Florida Entomol., 82: 556-569.
Direct Link - Hoddle, M.S., R.G. Van Driesche, S.M. Lyon and J.P. Sanderson, 2001. Compatibility of insect growth regulators with Eretmocerus eremicus (Hymenoptera: Aphelinidae) for Whitefly (Homoptera: Aleyrodidae) control on poinsettias I. laboratory assays. Biol. Cont., 20: 122-131.
CrossRef - Jervis, M.A., G.E. Heimpel, P.N. Ferns, J.A. Harvey and N.A.C. Kidd, 2001. Life-history strategies in parasitoid wasps: A comparative analysis of ovigeny. J. Animal Eol., 70: 442-458.
CrossRefDirect Link - Jones, W.A. and S.M. Greenberg, 1998. Suitability of Bemisia argentifolii (Hom.: Aleyrodidae) instars for the parasitoid Eretmocerus mundus (Hym.: Apheliniae). Environ. Entomol., 27: 1569-1573.
Direct Link - Jones, W.A., S.M. Greenberg and B. Legaspi Jr., 1999. The effect of varying Bemisia argentifolii and Eretmocerus mundus ratios on parasitism. BioControl, 44: 13-28.
CrossRef - Lewis, W.J., J.Q. Stapel, A.M. Cortesero and K. Takasu, 1998. Understanding how parasitoids balance food and host needs: Importance to biological control. Biol. Control, 11: 175-183.
CrossRef - Mandour, N.S., S.X. Ren and B.L. Qiu, 2007. Effect of Bemisia tabaci honeydew and its carbohydrates on search time and parasitization of Encarsia bimaculata. J. Applied Entomol., 131: 645-651.
CrossRef - McAuslane, H.J. and R. Nguyen, 1996. Reproductive biology and behavior of a thelytokous species of Eretmocerus (Hymenoptera: Aphelinidae) parasitizing Bemisia argentifolii (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am., 89: 686-693.
Direct Link - Pickett, C.H., G.S. Simmons, E. Lozano and J.A. Goolsby, 2004. Augmentative biological control of whiteflies using transplants. BioControl, 49: 665-688.
CrossRef - Qiu, B.L., P.J. De Barro and S.X. Ren, 2005. Development, survivorship, and reproduction of Eretmocerus sp. nr furuhashii (Hymenoptera: Aphiliendae) parasitizing Bemisia tabaci (Hemiptera: Aleyrodidae) on glabrous and non-glabrous host plants. Bull. Entomol. Res., 95: 313-319.
CrossRef - Rodriguez-Rodrigez, M.D., R. Moreno, M.M. Tellez, M.P. Rodriguez- Rodrigez and R. Fernandez-Fernandez, 1994. Eretmocerus mundus (Mercet), Encarsia lutea(Masi) and Encarsia transvena (Timberlake) (Hym., Apheliniae), parasitoids of Bemisia tabaci (Homoptera: Aleyrodidae) in protected vegetable crops in Almeria. Boletin de Sanidad Vegetal, Plagas, 20: 695-702.
- Romeis, J. and C.P.W. Zebitz, 1997. Searching behavior of Encarsia formosa as mediated by color and honeydew. Entomol. Exp. Appl., 82: 299-309.
CrossRef - Salvucci, M.E., G.R. Wolfe and D.L. Hendrix, 1997. Effects of sucrose concentration on carbohydrate metabolism in Bemisia argentifolii: Biochemical mechanism and physiological role for trehalulose synthesis in the silverleaf whitefly. J. Insect Physio., 43: 457-464.
CrossRef - Stansly, P.A., J. Calvo and A. Urbaneja, 2005. Release rates for control of bemisia tabaci (Homoptera: Aleyrodidae) biotype Q with Eretmocerus mundus (hymenoptera: Aphelinidae) in greenhouse tomato and pepper. Biol. Control, 35: 124-133.
CrossRef - Tuda, M. and M.B. Bonsall, 1999. Evolutionary and populations dynamics of host-parasitoid interactions. Res. Pop. Ecol., 41: 81-91.
Direct Link - Urbaneja, A., E. Sánchez and P.A. Stansly, 2007. Life history of Eretmocerus mundus, a parasitoid of Bemisia tabaci, on tomato and sweet pepper. BioControl, 52: 25-39.
CrossRef - Van Driesche, R.G., M.S. Hoddle, S.M. Lyon and J.P. Sanderson, 2001. Compatibility of insect growth regulators with Eretmocerus eremicus (Hymenoptera: Aphelinidae) for whitefly (Homoptera: Alyerodidae) control on poinsettias II. Trials in commercial poinsettia crops. Biol. Control, 20: 132-146.
CrossRef - Van Lenteren, J.C., M.M. Roskam and R. Timmer, 1997. Commercial mass production and pricing of organisms for biological control of pests in Europe. Biol. Control, 10: 143-149.
CrossRef - Vet, L.E.M. and J.C. Van Lenteren, 1981. The parasite-host relationship between Encarsia formosa gahan (Hymenoptera: Aphelinidae) and Trialeurodes vaporariorum westwood (Homoptera: Alyerodidae). Zeitschrift fur Angewandte Entomol., 91: 327-348.
CrossRefDirect Link - Zolnerowich, G. and M. Rose, 1998. Eretmocerus Haldeman (Hymenoptera: Aphelinidae) imported and released in the United States for control of Bemisia tabaci complex) (Homoptera: Aleyrodidae). Proc. Entomol. Soc. Washington, 100: 310-323.
Direct Link