
S. Reza Kazemi Nezhad
Department of Biology, Johannes Gutenberg University of Mainz, D-55099 Mainz, Germany
Franz Romer
Department of Biology, Johannes Gutenberg University of Mainz, D-55099 Mainz, Germany
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 4 | Page No.: 224-238







ABSTRACT
Oenocytes of the cricket, Gryllus bimaculatus, were characterized by a continuously high secretion of ecdysteroids over several hours. A factor extracted from heads of the male adults caused the oenocytes to enhance immunoreactive ecdysteroid secretion in vitro. This factor was isolated by high performance size exclusion chromatography. Its molecular weight ranged from 26.5 to 30.0 kDa. The partially purified bioactive substance stimulates the ecdysteroid synthesis of oenocytes in a time-and dose-dependent manner. The ecdysiotropin was susceptible to protease degradation and furthermore showed resistance to boiling and alkylation. Exopeptidase did not affect the activity of the peptidergic factor, suggesting that N- and C-terminus are protected. Treatments with dithiothreitol and neuraminidase suggest that disulfide bridge and carbohydrates are necessary for the biological activity of the oenocyte-stimulating ecdysiotropic factor. This study is the first evidence that a factor of head adult insect increased in vitro ecdysteroid secretion in oenocytes that are epidermal in origin.
PDF Abstract XML References Citation
How to cite this article
S. Reza Kazemi Nezhad and Franz Romer, 2008. A New Ecdysiotropic Factor in Adult Male Crickets. Journal of Entomology, 5: 224-238.
DOI: 10.3923/je.2008.224.238
URL: https://scialert.net/abstract/?doi=je.2008.224.238
DOI: 10.3923/je.2008.224.238
URL: https://scialert.net/abstract/?doi=je.2008.224.238
INTRODUCTION
It is well known that different endocrine mechanisms are concerned in the regulation of insect growth and development. Two insect ecdysiotropic systems, brain-prothoracic gland and brain-gonad (De Loof et al., 2001), have been identified. The prothoracic glands are mainly responsible for the synthesis and release of ecdysteroids in immature stages. They are controlled by the cerebral neuropeptide, prothoracicotropic hormone (PTTH). After release into the hemolymph, PTTH activates the prothoracic glands to synthesize ecdysteroids that in turn induce and synchronize moulting in the epidermis and other target tissues (Bollenbacher et al., 1993). Thus a neuroendocrine axis is needed to initiate the individual steps of the moulting process.
Peptides have been identified, which appear to stimulate secretion of ecdysteroids outside the prothoracic glands (Koolman, 1995). For example, the stimulation of steroid production is effectuated in the testes of Lymantria dispar by a testis ecdysiotropin (Loeb et al., 1988), in the ovaries of Locusta migratoria by follicle cell tropic hormone (Charlet et al., 1979) and in the ovaries of Aedes aegypti by an egg development neurosecretory hormone (Hagedorn et al., 1979). Ecdysiotropic factors for cells of epidermal origin (oenocytes and abdominal integument) are not well known.
It is indisputable that in most insects the prothoracic glands degenerate after imaginal ecdysis, so that the primary source for the production and formation of moulting hormones is exhausted. In male crickets they degenerate in the course of the moulting phase that leads to adulthood. At the same time ecdysteroid secretion of the glands ceases (Martau and Romer, 1998). It was known from earlier studies that adult male crickets undergo rapid changes in their hormonal household between 9 and 12 day after imaginal moult. A low level of hormone is characteristic for day 9. In contrast, day 12 is characterized by a drastic increase in ecdysteroid amount (Shahab and Romer, 1982). The extremely high titers of ecdysteroids in adult male Gryllus appear at distinct times during the imaginal period (Behrens and Hoffmann, 1982; Shahab and Romer, 1982), which implies a de novo synthesis and not a transfer from the larval stage (Hoffmann and Behrens, 1982). However, the existence of ecdysteroids in adult males appears necessary and may play an important function in spermatogenesis (Hagedorn, 1985; Loeb et al., 1988). Therefore, sources other than the prothoracic glands have to be involved in the production of the hormone in adult Gryllus bimaculatus (Bressel et al., 1990; Weidner et al., 1992; Hoffmann and Wagemann, 1994). Such sites of synthesis have been demonstrated in the epidermis of other insects (Delbecque et al., 1990; Jenkins et al., 1992). As a candidate amongst others the oenocytes came into consideration (Romer, 1974; Romer et al., 1974). The oenocyte must be classified as a secondary source of ecdysteroids (Redfern, 1989; Delbecque et al., 1990). In the cricket they are embedded in the fat body and occur more frequently in the neural sinus. The principal secretion product of oenocytes is 20-hydroxyecdysone (Romer and Bressel, 1994). It is also important to know, how the secretion activity of the oenocytes is regulated. In order to answer these and related questions we developed an in vitro assay of oenocytes. The hormones secreted were quantified by radioimmunoassay (RIA).
In the present study we describe the occurrence of an ecdysiotropic factor in head extracts of male adults of the Mediterranean cricket that has not been published previously. The stimulating effect is significant, dose-dependent and has a first maximum response after 1 h incubation. We suggest the new ecdysiotropin be referred to as oenocyte-stimulating ecdysiotropic factor (OSEF).
MATERIALS AND METHODS
Experimental Animals
Gryllus bimaculatus (Ensifera, Gryllidae) were reared under mass-breeding conditions at room temperature and a photoperiod of 14:10 (L/D) hours. Under these conditions the last larval stage (L9) lasts 12 days. For the individual experiments the male insects were isolated in separate glasses at 27°C and 60% relative humidity. The day of ecdysis was designated as day zero. Nine and 11 day old crickets (days after imaginal ecdysis) were used for the preparation of tissues and the head extracts, respectively. They were fed with fish flakes, milk powder, porridge oats, fresh salad leaves and water.
Purification of Samples
The incubates were applied onto a reversed-phase C18 Sep-Pak® (Waters Associates) after filling up with ddH2O to 5 mL (Lafont et al., 1982). The free ecdysteroids were eluted with 60% (v/v) methanol: water. This fraction was used for RIA.
Ecdysteroid RIA
The content of ecdysteroids in the incubation medium and tissue was quantified by the RIA as described by Spindler et al. (1978). Separation of bound from free radiolabel was by the ammonium sulphate method (Warren et al., 1984).
Briefly, the ecdysteroids were extracted with methanol and separated using C18 cartridges. An aliquot was evaporated in vacuo and then subjected to RIA. The antiserum (final concentration 0.013%) was induced in a rabbit against 20-ecdysone-2-hemisuccinate coupled with thyroglobulin, which shows the greatest specificity for the ecdysteroid side chain (Soumoff et al., 1981). The ratio of the cross-reaction of this antiserum between ecdysone and 20-hydroxyecdysone was 1:2.2, respectively. Tritiated ecdysone (specific activity: 53.0 Ci/mmol) from New England Nuclear Corp. (Boston) was used as the radioligand. The quantity of ecdysteroid was determined by the use of a β-counter. The assay was calibrated with 20-hydroxyecdysone (Simes, Milan, Italy) as the standard (working range: 0.1-32.0 ng) for each analysis.
Separation of Ecdysteroids by High-Performance
Liquid Chromatography (HPLC)
To cover the spectrum of ecdysteroids a part of the purified extracts were analyzed by HPLC. Reversed-phase HPLC (RP-HPLC) was performed via a Bischoff Lichrosorb RP 18 (4.0x250 mm, 7 μm particle size) with isocratic conditions, using 50% methanol in water, at 1.0 mL min-1 flow rate for 45 min. UV absorption was monitored at 242 nm (Model LC-UV, Pye Unicam, Cambridge). Fractions were collected every 0.5 min, dried in vacuum and stored at -20°C. The ecdysteroid content of each fraction was assayed by RIA. Standard ecdysteroids (ecdysone and 20-hydroxyecdysone) were applied to the HPLC under the conditions described earlier.
Size-Exclusion Chromatography
Two hundred heads (approx. 16 g) of 11 day old male adults of Gryllus bimaculatus were homogenized in an ice-cooled Dounce homogenizer (Braun; Melsungen) with methanol:water:trifluoroacetic acid (90:10:0.1, v/v/v) and centrifuged (12,000 g, 4°C, 15 min). The volume of the resulting supernatant was reduced to 200 μL in a vacuum concentrator and fractionated on an HP-SEC system pumping (1 mL min-1, wavelength 214 nm) acetonitrile:water:trifluoroacetic acid (40:60:0.1, v/v/v) per two Protein-Pak 125 (7.8x300 mm, Waters Corp., Milford) size exclusion columns arranged in tandem. Fractions were collected every 0.5 min in polypropylene microcentrifuge tubes. Then the fractions were dried, dissolved in Grace`s medium and stored at 4°C until assayed for ecdysiotropic activity. Molecular weight was estimated from a standard curve generated from retention times of bovine serum albumin (66 kDa), carbonic anhydrase (29 kDa), cytochrome c (12.4 kDa) and aprotinin (6.5 kDa). All protein markers were from Sigma Chemical Co. (Munich, Germany).
Tissue Preparation and in vitro Incubation
All dissections and in vitro incubations were performed under sterile conditions. Insects were anesthetized by water submersion, their surface sterilized with ethanol and the oenocytes were carefully removed under cricket Ringer (containing 86 mM NaCl, 5.4 mM KCl and 3 mM CaCl2). The tissue was then cleaned of remaining muscle and tracheae. After washing in fresh Ringer solution to remove adhering hemolymph, the tissue was first pre-incubated in Grace´s medium for 10 min and then transferred into incubation vials. The oenocytes were incubated for a period of 5 h under continuous oxygen supply and gentle shaking in 500 μL Grace`s medium without hemolymph (GIBCO, Grand Island, NY) containing 0.1 mg mL-1 penicillin and 0.3 mg mL-1 streptomycin. After every hour 100 μL of the incubation medium was removed, fixed with 100 μL 100% methanol for RIA and replaced with an equal volume of fresh medium. This method was used to examine the influence of medium replacement on the time-course of ecdysteroid secretion. For dose-response experiments, a net synthesis assay was used by subtracting the amount of ecdysteroid secreted during the 3rd hours of incubation in medium from the amount secreted during the following hour in the presence of head extracts.
Prothoracic glands from precisely timed larvae (day 4 of L9) were dissected in cricket Ringer (see above). Glands were briefly washed in Ringer and then incubated for 1 h with gentle shaking in 500 μL Grace`s medium containing either eluates from the HP-SEC (collected in five groups of fractions) or control medium. After incubation, the released ecdysteroid was determined by RIA.
Characteristics of the Ecdysiotropic Factor
The fractions from 24.5 to 26.5 min of HP-SEC that caused an increase of ecdysteroid secretion in the bioassay were pooled together. The pooled fractions were dried, dissolved in 200 μL Grace`s medium and afterwards examined for biochemical properties. In the experiments, 0.5 head equivalents per 500 μL incubation medium have been used.
Heating
Aliquots of the pooled fractions were incubated 1 to 4 min at a temperature of 100°C and then tested for ecdysiotropic activity.
Treatments with N-ethylmaleimide (NEM) and Dithiothreitol (DTT)
In order to test the involvement of sulfhydryl groups in OSEF activity, the reaction with NEM was carried out as suggested by Leslie et al. (1962). The reduction of disulfide bonds with DTT was performed as described by Sondack and Light (1971).
For the alkylation 0.1 mg NEM (Sigma) was dissolved in 50 μL 0.05 M Tris-HCl (pH 7.8) containing 6 M Urea, 1 mM EDTA-2Na and 2% n-butanol and then thoroughly mixed with the same amount of pooled active fraction. The mixture was allowed to react for 1 h at room temperature.
The disulfide bonds of OSEF were reduced by adding 0.1 mg DTT to the buffer described above and the mixture was stirred for 90 min at room temperature. The control samples contained neither NEM nor DTT.
Enzymatic Digestion
Trypsin from bovine pancreas (2750 NF units mg-1, Serva) was dissolved in 1 mM HCl and 20 mM CaCl2 at a concentration of 10 μg mL-1. In the following step, 10 μL of this enzyme solution were mixed with 50 μL of the active sample. This mixture was then incubated at 30°C for 1 h on a shaker and then heated for 2 min in boiling water. Enzyme and substrate thus had a volume ratio of 1:5.
Leucine aminopeptidase from porcine kidney (10 units mg-1, Sigma) was dissolved in 0.1 M ammonium acetate buffer (pH 8.1) at a concentration of 1 mg/100 mL. This mixture was then incubated with the sample at 37°C for 24 h. After this incubation it was heated at 100°C for 2 min. Enzyme and substrate thus had a volume ratio of 2.5:1.
Carboxypeptidase A from bovine pancreas (41.2 units mg-1, Sigma) was dissolved in 0.1 M ammonium acetate buffer (pH 8.1) containing 10% NaCl at a concentration of 500 μg mL-1. Fifty microliters of the enzyme solution and 50 μL of the sample were incubated at a temperature of 37°C for 24 h. Afterwards it was heated in boiling water for 2 min. Enzyme solution and substrate thus had a volume ratio of 1:1.
Neuraminidase from a bacterium Clostridium perfringens (1 unit mg-1, Sigma) was dissolved in 0.1 M ammonium acetate buffer (pH 5.5) at a concentration of 1 μU mL-1. One milliliter of the enzyme solution was then mixed with 50 μL of the biologically active sample and incubated at 37°C for 1 h. The hydrolysis was terminated by 2 min of heating at 100°C.
Statistical Analyses
Statistical evaluations were made using Student`s t-test (computer program WinSTAT 3.1). The results are means±SE for the number of individual measurements indicated in the legend of each figure.
RESULTS
Effects of Head Extract on Target Tissues
Frozen heads (- 60°C) were homogenized and centrifuged as described above. The supernatant was taken to 200 μL by vacuum centrifugation. The head extracts were subjected to HP-SEC and the eluates were divided into five groups of fractions G1: 14-21 min, G2: 21.5-28.5 min, G3: 29-36 min, G4: 36.5-43.5 min and G5: 44-50 min retention time. Each of these groups of fractions was tested for
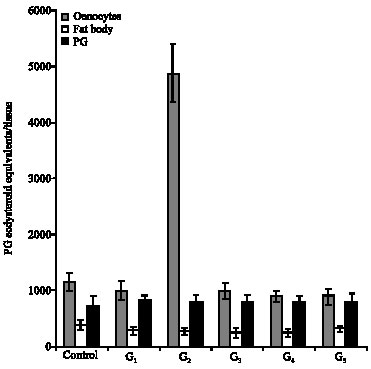 |
| Fig. 1: | Effects of head extract from HP-SEC on ecdysteroid secretion by oenocytes, thoracic fat body of 9 day old adult Gryllus bimaculatus and prothoracic glands (PG) of the last-larval instar (day 4). The eluates were collected in 0.5 min intervals and pooled in five fractions (G1-G5). Tissues were incubated for 1 h in 500 μLGrace`s medium with or without (control) the five fractions. Medium was analyzed for ecdysteroids by RIA. Each column represents the mean of five separate determinations. Bars denote standard error of the mean |
its capacity to elevate the secretion rate of ecdysteroids in oenocytes, thoracic fat body (not containing oenocytes) and nymphal prothoracic glands. It is obvious that only oenocytes were activated by eluates of group 2 (about the 5-fold in comparison to controls). Fat body and nymphal prothoracic glands are not affected by exposure to extracts (Fig. 1).
Molecular Weight Determination of the Biologically Active Factor
Extracts were made from whole heads of 11 day old male crickets. The isolation of the brain was difficult and too time-consuming due to very strong head musculature. Head extracts contained no immunologically detectable ecdysteroids.
To determine the molecular size of OSEF, head extract was fractionated on a size exclusion system with an acidic organic solvent. Only the fractions from 24.5 to 26.5 min retention time (a total of 5 fractions from G2), with a major peak at 25.5 min, stimulated the secretion of ecdysteroids in the oenocytes. Oenocytes were exposed to approx. 0.5-head equivalent for 1 h incubation. The molecular weight could be estimated between 26.45 and 30.02 kDa by using linear regression equation (Fig. 2).
Time-Course and Dose-Response Relationship of Ecdysteroid Secretion
The ability of the isolated and partially purified head extracts to stimulate ecdysteroid release by the oenocytes in vitro was tested using the oenocytes of day 9 old adult animals. Our (unpublished) observation has established that these oenocytes continue to secrete ecdysteroids in incubation medium at a low release rate and can be readily activated to show a well-recognizable increase in secretion when
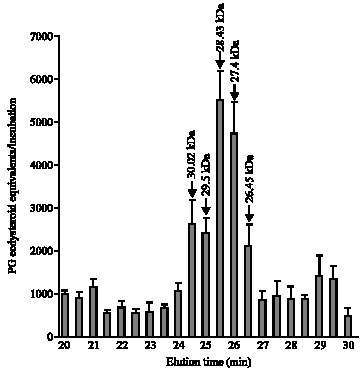 |
| Fig. 2: | HP-SEC Determination of the molecular size of the oenocyte stimulating ecdysiotropin present in the head extracts of 11 day old male adults of Gryllus bimaculatus. Molecular weights were estimated between 26.45 and 30.02 kDa (fractions of groups 2) by using the linear regression equation (r2 = 0.99), y = 1.1047Ln(x) + 5.7392, y is molecular weight and x the retention time in minutes. The molecular standards used were bovine serum albumin (66 kDa, Ve/Vo = 1.16 mL), carbonic anhydrase (29 kDa, Ve/Vo = 1.96 mL), cytochrome c (12.4 kDa, Ve/Vo = 2.92 mL) and aprotinin (6.5 kDa, Ve/Vo = 3.72 mL). The void volume (Vo) of the Dextran blue was 12.5 mL. Ve/Vo characterized the retention coefficient. Arrows show OSEF activity. Each data point represents the mean±SEM of four replicates |
head extracts are added to the medium. The profile of the ecdysteroid level secreted by oenocytes was investigated over a period of 5 h. Oenocytes are characterized by a continuously high secretion during the first 1-3 h of incubation (Fig. 3a).
Using a constant dose of 0.5 head equivalents per oenocyte culture, the incubation time was varied. Addition of the head extract leads to a rapid increase of ecdysteroid secretion from 0.8±0.2 to 2.27±0.45 ng after 1 h (Fig. 3a). The maximum concentration (total ecdysteroid content: 4.14±0.55 ng) of the RIA-detectable hormones was reached after 4 h incubation time, an about four-fold increase of the hormone released was observed as compared to control (1.1±0.31 ng). Extending the length of culture time did not produce a significant increase in overall ecdysteroid content.
The head extract elicited a dose-dependent increase in ecdysteroid secretion by oenocytes (Fig. 3b). Serial dilutions of extract evoked a range of responses from oenocytes in vitro that approximates a sigmoidal curve with a plateau reached at 0.5 head equivalents. The secretion of ecdysteroid was strongly elevated more than 2.5-fold by the addition of 0.4 head equivalents. Maximum ecdysteroid output was induced by 0.5 head equivalents. Addition of more extract did not lead to a higher response. Since the oenocytes released almost constant amounts of ecdysteroids to the medium in the first hours of incubation, different doses of diluted head extracts (0.1-1 head equivalents) have been added to the incubate after the third hour. Incubation was then prolonged for one more hour to observe the effects on the dose-response relationship. The difference between the amount of
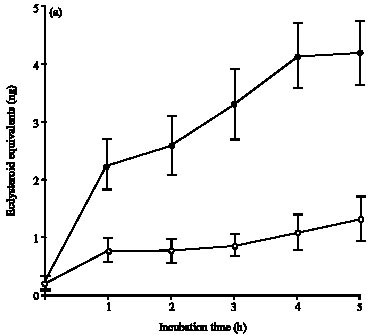 |
| Fig. 3a: | Time course of accumulation of ecdysteroid immunoreactivity in the medium secreted by oenocytes of day 9. Oenocytes were incubated in the presence (•) of 0.5 equivalents per incubation or absence (Ο) of adult head extracts from day 11. The medium was changed every hour and analyzed for ecdysteroids by means of RIA. Time zero represents the ecdysteroid extracted from oenocytes that were not incubated. Each point represents the mean ±SEM of 10 separate determinations |
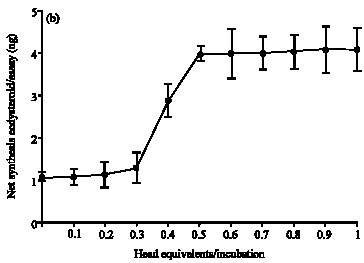 |
| Fig. 3b: | Dose-response of head extracts on the ecdysteroid secretion by oenocytes in vitro. Immunoreactive ecdysteroid secretion by oenocytes dissected from 9 day old male adults of Gryllus bimaculatus in response to increasing amounts of adult head extracts from day 11. Time zero represents the ecdysteroids extracted from oenocytes that were not incubated. Net synthesis was determined by RIA subtracting the ecdysteroids in the medium after the 3rd h from that after the 4th h. Data obtained by RIA are presented as 20-hydroxyecdysone equivalents. Eight bioassays were performed for each data point Bars indicate±SEM |
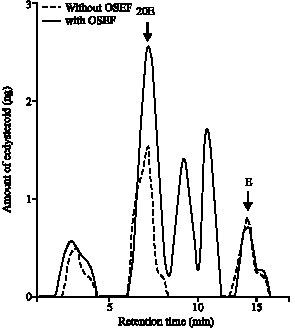 |
| Fig. 4: | Reversed-phase HPLC profile of the free ecdysteroid fraction secreted from oenocytes (day 9) in the presence or absence of OSEF. Fractions were collected every 0.5 min for RIA. Retention time of authentic 20-hydroxyecdysone (20E) and ecdysone (E) are indicated by arrows |
secreted hormones in the fourth hour of incubation, i.e., after application of the extracts and the secreted ecdysteroids in the third hour of incubation (before application of the extracts) is denoted as net synthesis. This is a measure of activation (Fig. 3b).
These results clearly indicate that the head extracts contain a factor which acts directly on oenocytes to stimulate ecdysteroid secretion.
Identification of Secretion Products by RP-HPLC/RIA
Ecdysteroids were extracted and partially purified as described in Materials and Methods (60% methanol/water fraction containing free ecdysteroids from Sep-Pak C18 separation). Subsequently to identify the composition of ecdysteroids in untreated and with head extract treated oenocytes were (equal amounts) subjected to HPLC/RIA analysis. Two distinctive peaks of immunoreactive components were detected in both samples that had identical retention times as authentic 20-hydroxyecdysone (7.5 min) and ecdysone (13.8 min) standards (Fig. 4). With OSEF treated oenocytes showed a marked increase (1.5-fold as untreated oenocytes) of the twenty-hydroxyecdysone-like component. Furthermore, two new unidentified RIA-active compounds (10 and 11.5 min) could be observed only after addition of the head extract to oenocytes culture. The results were reconfirmed also by means of normal-phase HPLC (data not shown).
Biochemical Characteristics of the OSEF
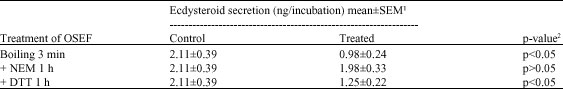
OSEF is stable to heat-treatment up to 2 min (Table 1) as well as to exposure to aqueous acetonitrile as the solvent.
NEM as an alkylator of the sulfhydryl groups does not influence the stimulatory effect in a significant manner (p>0.05). If at all free SH groups are present in OSEF, they play no role in the activity of ecdysiotropin. In contrast, intact disulfide bonds are important for biological activity (Table 1). On incubation with DTT, the effect of OSEF was significantly reduced (p<0.05).
| Table 1: | Characteristics of the OSEF |
 | |
| 1Mean and SEM of 4 assays. 2Titers are statistically different at α = 0.05 with the Student`s t-test | |
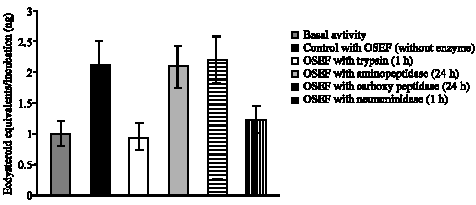 |
| Fig. 5: | Effects of endopeptidase, exopeptidase and neuraminidase on the OSEF-stimulated ecdysteroid secretion. Oenocytes of 9 day old adult Gryllus bimaculatus were incubated alone (basal activity), with OSEF (comprising 0.5 head equivalents) or digestion enzyme + OSEF in 500 μL Grace`s medium. Data are expressed as total immunoreactive material and presented as mean of four separate determinations. Bars denote standard error of the mean |
The treatment with trypsin destroyed all of the ecdysiotropic activity. Hormone production was lowered to the basal value (Fig. 5). Inactivation by trypsin makes it probable that biological activity of OSEF involves basic amino acids.
Incubation of OSEF with leucine aminopeptidase and carboxypeptidase A (final concentration: 2 mU mL-1) for 24 h did not affect OSEF activity. It is thus indicated that N- and C-terminus of the ecdysiotropin are blocked (Fig. 5). The biological activity of OSEF was approximately lost 80% by after 1 h incubation with neuraminidase (final concentration: 0.75 μl mL-1). Thus, we obtained evidence suggesting the necessity of carbohydrates for OSEF activity (Fig. 5).
DISCUSSION
Present data suggest the existence of a substance with oenocyte ecdysiotropic activity in the head of adult males of Gryllus bimaculatus. While synthesis and release of ecdysteroids in the prothoracic glands are regulated by release of PTTH from the brain (Bollenbacher et al., 1979; Westbrook and Bollenbacher, 1990), nothing is up to now published about the hormone which controls ecdysteroid release from epidermis, including oenocytes.
The present study clearly shows that a factor extracted from heads, was capable of stimulating immunodetectable ecdysteroid secretion by oenocytes in vitro above their basal activity (Fig. 1). The degeneration of the prothoracic glands of the male cricket in last instar nymphs as well as in adult animals was documented by Martau and Romer (1998). The glands start to degenerate already before the imaginal moult. The availability of moulting hormone in adult insects with a long-lasting mature stage, such as in the cricket (45 days), poses the question about the regulation of the ecdysteroidogenesis. Because the vast majority of adult insects no longer possess functioning prothoracic glands, it is certain that alternative sources of ecdysteroids exist and possibly regulated by superordinate centers. Among others (in particular, testis and ovary) the oenocytes were proposed as an alternative site of ecdysteroid secretion outside the prothoracic glands (Locke, 1969; Bonner-Weir, 1970). These findings were confirmed as a result of ultrastructural and in vitro synthesis of ecdysteroids by the oenocytes (Romer, 1974; Romer et al., 1974; Rinterknecht and Matz, 1983; Romer and Bressel, 1994). The oenocytes of insects are derivatives of the ectoderm and therefore represent specialized cells of the epidermis (Harmsen and Beckel, 1960; Lawrence and Johnston, 1982). It may not be surprising that the epidermal tissue can synthesize ecdysteroids, if one considers that prothoracic glands and epidermis share a common ectodermal origin and that it has been proven that the epidermal tissues of various insects can act as a source of ecdysteroids in the absence of prothoracic glands (Redfern, 1989; Delbecque et al., 1990). In experiments with an insect cell line of epidermal origin it was shown that alternate sites for ecdysteroid production in vivo may exist and could play a role in local regulation of development (Spindler and Spindler-Barth, 1991; Mesnier et al., 2000).
The oenocytes of Gryllus bimaculatus are located in the neural sinus lateral to the abdominal nerve cord embedded in the fat body and it is impracticable to isolate them from the adipose tissue. Therefore, we prepared the entire complex consisting of oenocytes and adipose tissue for use in the following experiments. Furthermore, an even distribution of the oenocyte cluster in the left and right side was not possible as because of its complex anatomic structure. Thus a suitable in vitro assay was developed in order to determine the secretion kinetics (Fig. 3a). The possibility that the observed ecdysteroid synthesis or secretion in medium is due to any contaminating fat body tissue associated with the oenocytes can be excluded, since such synthesis was not observed in previous studies using fat body alone (Shahab and Romer, 1982; Bressel et al., 1990). This is in agreement with our findings (Fig. 1). A longer storage of hormones was also not found in the fat body (Bulenda et al., 1986). However, fat body tissue contains ecdysone 20-mono-oxygenase, required for hydroxylation of ecdysone into the more active 20-hydroxyecdysone (Zhu et al., 1983).
Head extracts fractionated by means of HP-SEC method with an acidic organic solvent showed stimulating activity of oenocytes (Fig. 2). We found significant activity in 5 fractions, but because those fractions lay adjacent to one another, we conclude the existence of a single ecdysitropic factor, that was spread in varying concentrations in the fractions of 24.5-26.5 min. The molecular size of OSEF could be determined in a range between 26.5 and 30 kDa. The most active fractions had a molecular weight of 28.4 and 27.4 kDa. OSEF does not stimulate ecdysteroid synthesis in prothoracic glands and fat body of Gryllus (Fig. 1).
Molecular variants of PTTH appear to exist in each class size. In the three well-studied insect species with regard to PTTH, Bombyx mori, Manduca sexta and Lymantria dispar, PTTH having two molecular weight ranges: 4-7 kDa for small PTTH and 11-30 kDa for big PTTH (Masler et al., 1986; Nagasawa et al., 1986; Bollenbacher et al., 1993). For Drosophila melanogaster there exist conflicting results (Pak et al., 1992; Kim et al., 1997). This variation between insect species could be due to discrepancies in the degree of glycosylation or the conditions utilized for fractionation.
It is possible that one or even more biologically active molecules (in any case, larger than 30 kDa) were denatured by our solvent system. However, there is a certain risk that acidic organic solvents can reduce the activity of molecules through the dissociation of dimer into monomer. Above all, other authors have used organic solvent systems in their molecular weight determination studies and have detected two molecular sizes of PTTH (O`Brien et al., 1986; Kelly et al., 1992; Fescemyer et al., 1995) or only one small form of PTTH (Gelman et al., 1992). The use of organic solvent was successfully applied for determination of molecular mass of other insect neuropeptides such as egg development neurosecretory hormone (EDNH) from Aedes aegypti (Masler et al., 1983), testis ecdysiotropin (TE) from Lymantria dispar (Wagner et al., 1997) and ecdysiostatin from Calliphora vicina (Hua and Koolman, 1995), to only name some.
The oenocytes of 9 day old adult crickets were characterized by a low content of steroids. Therefore, oenocytes at this stage were well suitable, because they have shown the necessary sensibility for use in an in vitro assay for detection of OSEF activity. Studies on the prothoracic glands of Lepidoptera demonstrated that the glands containing a higher hormone pool could not be further activated by PTTH (Bollenbacher et al., 1983; Kelly et al., 1992). The amount of hormone produced by oenocytes was constant (0.8-0.9 ng) during the first 3 h of incubation. This allows a comparison of the secretion performance of the oenocytes with and without head extract. The oenocytes treated with head extract increase ecdysteroid secretion at all time points (Fig. 3a). Incubation of oenocytes with head extract results in enhanced, nearly linear increase in ecdysteroid secretion.
A dose-dependent activation response was generated (Fig. 3b). The method used (i.e., subtracting 3rd h synthesis of oenocytes from 4th h synthesis of oenocytes) allowed the determination of net synthesis. Similar in vitro bioassays were developed with prothoracic glands for the determination of PTTH activity (Okuda et al., 1985; Kelly et al., 1996). Further, the specificity of the in vitro bioassay was supported by the fact that partially purified head extract induced a sigmoidal dose-response curve. The stimulatory activity of OSEF showed a dose-dependency starting with 0.4 head equivalents causing a strong increase (approx. 3-fold) in oenocytes ecdysteroid content. However, the dose response to extract was only within a narrow range (0.3-0.5 head equivalents) linear in vitro, suggesting that the titer of OSEF may regulate ecdysteroid secretion by oenocytes in vivo. Increasing the extracts from 0.5 to 1 head equivalents in incubation medium did not further increase hormone secretion.
Romer and Bressel (1994) have reported that the secretion capacity of oenocytes was low at day 9 and showed a maximum at day 12. The timing of this is compatible with our in vitro observations. The head extracts from 11-day-old crickets contained ecdysiotropic factors, which caused the oenocytes of day 9 with low endogenous steroid levels to maximize hormone secretion. Interestingly, the rate of ecdysteroid formation after head extract addition is comparable to the increase in ecdysteroid levels observed in oenocytes from 12-day-old animals. The coincidence of these time periods is yet another indication that OSEF plays a role in ecdysteroid secretion in vivo.
The secretion products of oenocytes were analysed by means of RP-HPLC combined with RIA. The findings from this procedure showed a distinct increase of 20-OH-ecdysone and the appearance of two additional conceivably ecdysteroid peaks after treatment with OSEF which could not be defined more specifically (Fig. 4).
Biological activity in head extracts was not significantly affected by 2 min boiling, indicating that the extract comprised to some degree heat resistant ecdysiotropin (Table 1). The activity of OSEF was almost retained after treatment with NEM, suggesting that SH-groups are not contained in ecdysiotropin or are not responsible for the stimulating activity if present. It could be shown that OSEF is glycosylated with oligosaccharide chains and contains disulfide bond(s), which are necessary for its biological activity (Table 1). The carbohydrate residues are assumed to be required for binding to the receptor and the disulfide bonds for stabilization of the secondary structure. The occurrence of disulfide bonds in OSEF is a sign for its di- or oligomer formation. In contrast to OSEF, neuraminidase had no effect on PTTH (Yamazaki and Kobayashi, 1969; Matsuo et al., 1985). PTTH of Lepidoptera maintains prothoracotropic activity if expressed in E. coli (Kawakami et al., 1990; Gilbert et al., 2000). Therefore, the glycosylation of PTTH is not necessary for biological activity (Adachi-Yamada et al., 1994).
In addition, OSEF was treated with protease and peptidase (Fig. 5). The isolated ecdysiotropic factor was sensitive to trypsin digestion and, like PTTH (Bollenbacher et al., 1984; Nagasawa et al., 1984), EDNH (Hagedorn et al., 1979) and testis ecdysiotropin (Loeb et al., 2001), appears to be a peptide, because all activity is destroyed by incubation of the head extract with trypsin before incubation with oenocytes. The results indicated more that both the N- and the C-Terminus were protected against exopeptidases. This was also confirmed by the attempted sequencing analysis (Dr. Marcus Macht, Center for Molecular Medicine, University of Cologne, Germany). It is to be remembered that the peptidal neurohormones of some insects possess C- and N-termini which are inaccessible (Isobe et al., 1975; Stone et al., 1976; Matsuo et al., 1985; Kataoka et al., 1991).
In summary, the results indicate the occurrence of a novel endocrine axis-head-oenocyte in adult males of Gryllus bimaculatus.
ACKNOWLEDGMENT
We are grateful to Professor Dr. August Dorn from University of Mainz, Germany (Institute of General Zoology) for critical reading of the manuscript.
REFERENCES
- Adachi-Yamada, T., M. Iwami, H. Kataoka, A. Suzuki and H. Ishizaki, 1994. Structure and expression of the gene for the prothoracicotropic hormone of the silkmoth Bombyx mori. Eur. J. Biochem., 220: 633-643.
CrossRefDirect Link - Bollenbacher, W.E., N. Agui, N.A. Granger and L.I. Gilbert, 1979. In vitro activation of insect prothoracic glands by the prothoracicotropic hormone. Proc. Natl. Acad. Sci. USA., 76: 5148-5152.
Direct Link - Bollenbacher, W.E., M.A. O'Brien, E.J. Katahira and L.I. Gilbert, 1983. A kinetic analysis of the action of the insect prothoracicotropic hormone. Mol. Cell. Endocrinol., 32: 27-46.
CrossRef - Bollenbacher, W.E., E.J. Katahira, M.A. OBrien, L.I. Gilbert and M.K. Thomas et al., 1984. Insect prothoracicotropic hormone: Evidence for two molecular forms. Science, 224: 1243-1245.
CrossRefDirect Link - Bollenbacher, W.E., R.S. Gray, D.P. Muehleisen, S.A. Regan and A.L. Westbrook, 1993. The biology of the prothoracicotropic hormone peptidergic neurons in an insect. Am. Zool., 33: 316-323.
CrossRefDirect Link - Bulenda, D., A. Stecher, M. Freunek and K.H. Hoffmann, 1986. Ecdysone metabolism in adult crickets, Gryllus bimaculatus. Insect Biochem., 16: 83-90.
CrossRef - Charlet, M., F. Goltzene and J.A. Hoffmann, 1979. Experimental evidence for a neurodocrine control of ecdysone biosynthesis in adult females of Locusta migratoria. J. Insect Physiol., 25: 463-466.
CrossRef - De Loof, A., G. Baggerman, M. Breuer, I. Claeys and A. Cerstiaens et al., 2001. Gonadotropins in insects: An overview. Arch. Insect Biochem. Physiol., 47: 129-138.
Direct Link - Delbecque, J.P., K. Weidner and K.H. Hoffmann, 1990. Alternative sites for ecdysteroid production in insects. Invertebr. Reprod. Dev., 18: 29-42.
CrossRefDirect Link - Fescemyer, H.W., E.P. Masler, T.J. Kelly and W.R. Lusby, 1995. Influence of development and prothoracicotropic hormone on the ecdysteroids produced in vitro by the prothoracic glands of female gypsy moth (Lymantria dispar) pupae and pharate adults. J. Insect Physiol., 41: 489-500.
CrossRef - Gelman, D.B., B.S. Thyagaraja, T.J. Kelly, E.P. Masler, R.A. Bell and A.B. Borkovec, 1992. Prothoracicotrophic hormone levels in brains of the European corn borer, Ostrinia nubilalis: Diapause vs the non-diapause state. J. Insect Physiol., 38: 383-395.
Direct Link - L.I., R. Rybczynski, Q. Song, A. Mizoguchi, R. Morreale and W.A. Smith et al., 2000. Dynamic regulation of prothoracic gland ecdysteroidogenesis: Manduca sexta recombinant prothoracicotropic hormone and brain extracts have identical effects. Insect Biochem. Mol. Biol., 30: 1079-1089.
Direct Link - Hagedorn, H.H., J.P. Shapiro and K. Hanaoka, 1979. Ovarian ecdysone secretion is controlled by a brain hormone in an adult mosquito. Nature, 282: 92-94.
CrossRefDirect Link - Harmsen, R. and W.E. Beckel, 1960. The intraovular development of the subspiracular glands in Hyalophora cecropia (L.)(Lepidoptera; Saturnidae). Can. J. Zool., 38: 883-893.
CrossRefDirect Link - Hoffmann, K.H. and W. Behrens, 1982. Free ecdysteroids in adult male crickets, Gryllus bimaculatus. Physiol. Entomol., 7: 269-279.
CrossRefDirect Link - Hoffmann, K.H. and M. Wagemann, 1994. Age dependency and tissue distribution of ecdysteroids in adult male crickets, Gryllus bimaculatus de Geer (Ensifera, Gryllidae). Comp. Biochem. Physiol., 109A: 293-302.
CrossRef - Isobe, M., K. Hasegawa and T. Goto, 1975. Further characterization of the silkworm diapause hormone A. J. Insect Physiol., 21: 1917-1920.
CrossRef - Jenkins, S.P., M.R. Brown and A.O. Lea, 1992. Inactive prothoracic glands in larve and pupae of Aedes aegypti: Ecdysteroid release by tissues in the thorax and abdomen. Insect Biochem. Mol. Biol., 22: 553-559.
CrossRef - Kataoka, H., H. Nagasawa, A. Isogai, H. Ishizaki and A. Suzuki, 1991. Prothoracicotropic hormone of the silkworm, Bombyx mori: Amino acid sequence and dimeric structure. Agric. Biol. Chem., 55: 73-86.
PubMedDirect Link - Kawakami, A., H. Kataoka, T. Oka, A. Mizoguchi and K.M. Kawakami et al., 1990. Molecular cloning of the Bombyx mori prothoracicotropic hormone. Science, 247: 1333-1335.
CrossRefDirect Link - Kelly, T.J., E.P. Masler, B.S. Thyagaraja, R.A. Bell and R.B. Imberski, 1992. Development of an in vitro assay for prothoracicotropic hormone of the gypsy moth, Lymantria dispar (L.) following studies on identification, titers and synthesis of ecdysteroids in last-instar females. J. Comp. Physiol. B, 162: 581-587.
CrossRefDirect Link - Kelly, T.J., T.G. Kingan, C.A. Masler and C.H. Robinson, 1996. Analysis of the ecdysiotropic activity in larval brains of the tobacco hornworm, Manduca sexta. J. Insect Physiol., 42: 873-880.
CrossRef - Kim, A.J., G.H. Cha, K. Kim, L.I. Gilbert and C.C. Lee, 1997. Purification and characterization of the prothoracicotropic hormone of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA., 94: 1130-1135.
PubMedDirect Link - Koolman, J., 1995. Control of ecdysone biosynthesis in insects. Neth. J. Zool., 45: 83-88.
CrossRefDirect Link - Lafont, R., J.L. Pennetier and M. Andrianjafintrimo, 1982. Sample processing for high-performance liquid chromatography of ecdysteroids. J. Chromatogr., 236: 137-149.
CrossRef - Lawrence, P.A. and P. Johnston, 1982. Cell lineage of the Drosophila abdomen: The epidermis, oenocytes and ventral muscles. J. Embryol. Exp. Morphol., 72: 197-208.
Direct Link - Leslie, J., D.L. Williams and G. Gorin, 1962. Determination of mercapto groups in protein with N-ethylmaleimide. Anal. Biochem., 3: 257-263.
CrossRef - Locke, M., 1969. The ultrastructure of the oenocytes in the molt/intermolt cycle of an insect. Tissue Cell, 1: 103-154.
PubMedDirect Link - Loeb, M.J., E.P. Brandt, C.W. Woods and R.A. Bell, 1988. Secretion of ecdysteroid by sheaths of testes of the gypsy moth, Lymantria dispar and its regulation by testis ecdysiotropin. J. Exp. Zool., 248: 94-100.
CrossRef - Loeb, M.J., A. De Loof, D.B. Gelman, R.S. Hakim and H. Jaffe et al., 2001. Testis ecdysiotropin, an insect gonadotropin that induces synthesis of ecdysteroid. Arch. Insect Biochem. Physiol., 47: 181-188.
Direct Link - Martau, T. and F. Romer, 1998. Degeneration of moulting glands in male crickets. J. Insect Physiol., 44: 981-989.
PubMedDirect Link - Masler, E.P., H.H. Hagedorn, D.H. Petzel and A.B. Borkovec, 1983. Partial purification of egg development neurosecretory hormone with reverse-phase liquid chromatographic techniques. Life Sci., 33: 1925-1931.
CrossRef - Masler, E.P., T.J. Kelly, B.S. Thyaragaja, C.W. Woods, R.A. Bell and A.B. Borkovec, 1986. Discovery and Partial Characterization of Prothoracicotropic Hormones of the Gypsy Moth, Lymantria dispar. In: Insect Neurochemistry and Neurophysiology, Borkovec, A.B. and D.B. Gelman (Eds.). Humana Press, Clifton, New Jersey, pp: 331-334.
- Matsuo, N., Y. Aizono, G. Funatsu, M. Funatsu and M. Kobayashi, 1985. Purification and some properties of prothoracicotropic hormone in the silkworm, Bombyx mori. Insect Biochem., 15: 189-195.
CrossRef - Mesnier, M., N. Partiaoglou, H. Oberlander and P. Porcheron, 2000. Rhythmic autocrine activity in cultured insect epidermal cells. Arch. Insect Biochem. Physiol., 44: 7-16.
Direct Link - Nagasawa, H., H. Kataoka, Y. Hori, A. Isogai and S. Tamura et al., 1984. Isolation and some characterization of the prothoracicotropic hormone from Bombyx mori. Gen. Comp. Endocrinol., 53: 143-152.
PubMedDirect Link - Nagasawa, H., H. Kataoka, A. Isogai, S. Tamura and H. Ishizaki, 1986. Amino acid sequence of a prothoracicotrophic hormone of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA., 83: 5840-5843.
Direct Link - O'Brien, M.A., N.A. Granger, N. Agui, L.I. Gilbert and W.E. Bollenbacher, 1986. Prothoracicotropic hormone in the developing brain of the tobacco hornworm, Manduca sexta: Relative amounts of the two molecular forms. J. Insect Physiol., 32: 719-725.
CrossRef - Okuda, M., S. Sakurai and T. Ohtaki, 1985. Activity of the prothoracic gland and its sensitivity to prothoracicotropic hormone in the penultimate and last-larval instar of Bombyx mori. J. Insect Physiol., 31: 455-461.
CrossRef - Pak, M.D., K.W. Chung, C.C. Lee, K. Kim, Y. Namkoong and J. Koolman, 1992. Evidence for multiple forms of the prothoracicotrophic hormone in Drosophila melanogaster and indication of a new function. J. Insect Physiol., 38: 167-176.
CrossRef - Rinterknecht, E. and G. Matz, 1983. Oenocyte differentiation correlated with the formation of ectodermal coating in the embryo of a cockroach. Tissue Cell, 15: 375-390.
PubMedDirect Link - Romer, F., 1974. Ultrastructural changes of the oenocytes of Gryllus bimaculatus DEG (Saltatoria, Insecta) during the moulting cycle. Cell Tissue Res., 151: 27-46.
PubMedDirect Link - Romer, F., H. Emmerich and J. Nowock, 1974. Biosynthesis of ecdysones in isolated prothoracic glands and oenocytes of Tenebrio molitor in vitro. J. Insect Physiol., 20: 1975-1987.
CrossRef - Romer, F. and H.U. Bressel, 1994. Secretion and metabolism of ecdysteroids by oenocyte-fat body complexes (OEFC) in adult males of Gryllus bimaculatus DEG (Insecta). Z. Naturforsch. Biosciences, 49: 871-880.
Direct Link - Sondack, D.L. and A. Light, 1971. Comparative studies on the modification of specific disulfide bonds of trypsinogen and chymotrypsinogen. J. Biol. Chem., 246: 1630-1637.
Direct Link - Soumoff, C., D.H.S. Horn and J.D. O'Connor, 1981. Production of a new antiserum to arthropod moulting hormone and comparison with two other antisera. J. Steroid Biochem., 14: 429-435.
CrossRef - Spindler, K.D., C. Beckers, U. Groschel-Stewart and H. Emmerich, 1978. A radioimmunoassay for arthropod moulting hormones, introducing a novel method of immunogen coupling. Hoppe-Seylers Z. Physiol. Chem., 359: 1269-1272.
PubMedDirect Link - Spindler, K.D. and M. Spindler-Barth, 1991. Ecdysteroid production and metabolism by an epithelial cell line from Chironomus tentans. Naturwissenschaften, 78: 78-79.
CrossRefDirect Link - Stone, J.V., W. Mordue, K.E. Batley and H.R. Morris, 1976. Structure of locust adipokinetic hormone, a neurohormone that regulates lipid utilisation during flight. Nature, 263: 207-211.
PubMedDirect Link - Wagner, R.M., M.J. Loeb, J.P. Kochansky, D.B Gelman, W.R. Lusby and R.A. Bell, 1997. Identification and characterization of an ecdysiotropic peptide from brain extracts of the gypsy moth, Lymantria dispar. Arch. Insect Biochem. Physiol., 34: 175-189.
CrossRefDirect Link - Warren, J.T., W.A. Smith and L.I. Gilbert, 1984. Simplification of the ecdysteroid RIA by use of protein A from Staphylococcus aureus. Experientia, 40: 393-394.
CrossRefDirect Link - Weidner, K., M. Clasz, H. Rieck and K.H. Hoffmann, 1992. Developmental changes in ecdysteroid biosynthesis in vitro during adult life and embryogenesis in a cricket, Giyllus bimaculatus de Geer. Invertebr. Reprod. Dev., 21: 129-139.
CrossRefDirect Link - Westbrook, A.L. and W.E. Bollenbacher, 1990. The development of identified neurosecretory cells in the tobacco hornworm, Manduca sexta. Dev. Biol., 140: 291-299.
CrossRefDirect Link - Yamazaki, M. and M. Kobayashi, 1969. Purification of the proteinic brain hormone of the silkworm, Bombyx mori. J. Insect Physiol., 15: 1981-1990.
CrossRef - Zhu, X.X., H. Gfeller and B. Lanzrein, 1983. Ecdysteroid during oogenesis in the ovoviviparous cockroach Nauphoeta cinerea. J. Insect Physiol., 29: 225-235.
CrossRef