
Donald A. Ukeh
School of Biological Sciences, University of Aberdeen, Tillydrone Avenue, AB24 2TZ, United Kingdom
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 3 | Page No.: 193-199







ABSTRACT
The essential oils extracted from Aframomum melegueta seeds and Zingiber officinale rhizomes were evaluated for their repellency against Rhyzopertha dominica in a four-armed airflow olfactometer. Parameters assessed were time spent and number of entries or visits made by male and female adults into the treated and control arms of the olfactometer. Ten microliters of both crude oil extracts significantly repelled the beetles when tested singly and in combination with 5 g winter wheat grains. These results suggest that the essential oils from A. melegueta and Z. officinale may be used in grain storage against insect pests. The details of the bioassay procedure used and the results obtained are reported.
PDF Abstract XML References Citation
How to cite this article
Donald A. Ukeh, 2008. Bioactivities of Essential Oils of Aframomum melegueta and Zingiber officinale both (Zingiberaceae) Against Rhyzopertha dominica (Fabricius). Journal of Entomology, 5: 193-199.
DOI: 10.3923/je.2008.193.199
URL: https://scialert.net/abstract/?doi=je.2008.193.199
DOI: 10.3923/je.2008.193.199
URL: https://scialert.net/abstract/?doi=je.2008.193.199
INTRODUCTION
The Lesser Grain Borer, Rhyzopertha dominica (Fabricius) (Coleoptera:Bostrichidae) is a destructive pest of stored grain throughout the warmer regions of the world (Ukeh and Umoetok, 2007). It is highly polyphagous and has been recorded feeding on diverse food crops such as cereals, legumes, tubers and packaging material made from wood (Edde et al., 2005). The major cereal crops commonly infested by the insect pest include maize, rice, wheat and sorghum (Hagstrum, 2001). R. dominica-infested grain decreases in value as a function of live insects, insect damage kernels, or insect fragments in milled products. R. dominica is not known to attack cereals in the field, but it is a strong flier and has been found infesting grain, in supposedly clean stores, within weeks or months after storage. Adults have very powerful mandibulate mouthparts and can bore into wood as well as grain (Gates, 1995; Rees, 2004). This rapid colonization behaviour, strong flight ability and broad polyphagy, coupled with the fact that R. dominica has been trapped in diverse environments, including wood-lands substantial distances from grain stores, suggests the movement of this pest between potential natural habitats and grain storage facilities. This movement by R. dominica is enhanced by the male-produced aggregation pheromones and host plant volatiles in storage houses (Landolt, 1997; Bashir et al., 2001). The adult beetles are 2-3 mm long and have cylindrical, dark brown or brown bodies. Eggs are laid singly and in small clusters between grains or in crevices in the storehouse walls and as this species are long lived, up to 500 eggs may be laid in total. After hatching, the larvae moults 2-5 times before pupation, this takes place inside the grain. Young larvae are not cannibalistic, but free living and as they mature, they become increasingly scarabaeiform and more immobile. Larvae and adults are voracious feeders of flour products as well as whole grains, causing characteristic tunnels. Detection of the species is difficult because larvae and adults tend to remain hidden inside the kernel (Vela-Coiffier et al., 1997). The minimum time to complete the life cycle under ideal conditions is 25 days at about 34°C and 70% rh, although such conditions are rarely found in the field. Effective control of R. dominica with minimal insecticide use requires an integrated management approach combining proper sanitation, monitoring and other environmentally friendly management practices (Stoll, 2000; Rees, 2004). There are some plants with repellent or behaviour-modifying properties which could be used to control R. dominica without the hazards associated with the use of synthetic pesticides such as insect resistance, human and eco-toxicity (Ukeh and Umoetok, 2007). The seeds, roots and leaves of alligator pepper, Aframomum melegueta (Rosk.) K. Schum and ginger, Zingiber officinale (Roscoe) (both Zingiberaceae) have been used in spicing meat, sauces and soups and mixed with other herbs for the treatment of body pains, catarrh, congestion, diarrhoea, sore throat and rheumatism in West Africa (Ajaiyeoba and Ekundayo, 1999).
This study describes the laboratory bioassay using vacuum-distilled volatile essential oils from A. melegueta seeds and Z. officinale rhizome for repellency against R. dominica in the four-arm airflow olfactometer.
MATERIALS AND METHODS
Insect Culture
R. dominica was obtained from stock culture maintained by the Central Science Laboratory, York, United Kingdom in March 2006 and reared on winter wheat seeds in a Constant Temperature and Humidity (CTH) room running at 25°C, 65% rh on a 12:12 DL photoperiod. Fifty pairs of adult R. dominica were introduced into 300 g wheat seeds in bell jars and allowed to feed on the grains for 12 days after which they were removed and discarded. After about 30 days, a laboratory test sieve size of 2 mm, (Endecotts Limited London, England) was used to sieve out the emerged insects daily and records were kept of their sexes and dates of emergence. Adult beetles were sexed by examination of the tip of the abdomen using a Nikon binocular microscope (Nikon House, Surrey, England). Males were identified by the presence of a punctuate groove on the fifth abdominal sternite (Ghorpade and Thyagarajan, 1980), which was rather shallow and of variable development, but was present on both sides or at least on one side of the mid-ventral line.
Plant Materials Collection and Preservation
The matured rhizomes of Z. officinale and seeds of A. melegueta, were collected from fields around the town of Akamkpa, Cross River State (situated between latitude 5°00' and 5°15’ North and longitude 8°04’ and 8°25’ East) in Southern Nigeria in December 2006. The identity of the plants was confirmed by the Crop Science Department of the University of Calabar, Nigeria. Z. officinale and A. melegueta plant products were selected on the basis of their ethnomedical studies and endemicity.
Vacuum Distillation of Plant Products
Fifty milliliter of re-distilled diethyl ether was added to 30 g of cut pieces of Z. officinale rhizomes. The contents were transferred to a 100 mL round bottomed flask connected to the vacuum distillation apparatus which was connected to a vacuum pump (Edwards ES50 Vacuum Pump, England). The U-tube and the pear-shaped vessel for the collection of the distillate was completely immersed in liquid nitrogen of -196°C and the extract residue was then distilled, under a vacuum of < 3mbar for 24 h. Also, 30 g A. melegueta seeds crushed in a mortar was vacuum distilled as explained earlier. Diethyl ether distillates of these substances were then pipetted from the vacuum distillation apparatus into a 50 mL separation funnel to remove water. The extracts were dried using magnesium tetraoxosulphate (VI) (MgSO4), filtered and concentrated to obtain 3 mL each Z. officinale and A. melegueta extracts. The vacuum distillates were sealed under nitrogen, labelled and stored at -20°C.
Bioassay Method
The olfactometer was a four-pointed star-shaped exposure chamber milled into a circular transparent plate with a diameter of 11.5 cm and which consisted of 3 layers of 6 mm Perspex (Rothamsted Research, Harpenden, England). The bottom of the apparatus was lined with a sheet of Whatman QL 100 filter paper (Whatman Limited, England). The olfactometer side arms made of socket glass (P.A. Brooks, Oxford, England) were inserted through the holes of the chamber walls. The air stream through the olfactometer was supplied by an Air entrainment system (KNF Neuberger, Germany) through Teflon tubing (Camlab Ltd., UK). Immediately after the pump, the air was divided through 2 carbon rods to clean it. From each carbon rod, the air stream was then further divided and pushed through two flow meters (GPE Limited, Leighton Buzzard, UK) to give a total of four air flows going into the behaviour chambers each at the rate of 200 mL min-1. Each air stream then passed through a glass side arm, which contained either the odour source or the control. From each glass side arm, air was delivered into the bioassay exposure chamber by the four air-delivery tubes and removed from the centre of the olfactometer at a rate of 800 mL min-1. The air streams formed four distinct zones in the chamber as confirmed by smoke tests. All the single choice bioassays were conducted in the CTH room between 9.00 am and 12.00 noon. The olfactometer was coded as five areas: One square shaped central area and four rectangular areas corresponding to the four arms of the olfactometer, each area was marked with a number between 1 and 5. In these trials, the test arm was given number 1; the 3 controls numbered 2, 3 and 4, respectively, while the central arena was given number 5. The test arm contained filter paper disc loaded with 10 μL A. melegueta or Z. officinale extract, or the extract +5 g wheat grains and the controls had 10 μL diethyl ether-loaded filter paper discs respectively. Beetles, whose responses were to be tested, were kept singly for 5 days after emergence and then starved for 24 h before being used for the bioassays. They were then releases into the centre of the olfactometer at one beetle per replication. Each beetle was observed for 10 min using a stopwatch and each trial was replicated 10 times. Odour samples were replaced after every 2 replications. After starting the experiment by releasing the insect into the centre of the olfactometer, the insect was followed visually as it made choices to any of the 4 olfactometer arms. A computer programme for collecting and analysing behavioural data with the four-armed olfactometer (33100 Udine, Italy) was used. The data recorded includes the time spent by the insect in the different areas of the olfactometer and the number of entries into each area or odour zone.
Statistical Analysis
The software Statistical Package for Social Scientists (SPSS) version 15 (SPSS 2007) was used for data analysis. A pair-wise t-test was used to test for significant differences between the treated arm and the mean of the control arms.
RESULTS AND DISCUSSION
These results have shown the repellent effect of 10 μL vacuum distilled seed extract of A. melegueta on male and female R. dominica, the mean time spent and the mean number of entries made being, respectively (0.92 min, p<0.0001), (2.5, p<0.0001) and (1.16 min, p<0.0001), (3.2, p<0.0001) compared to the control (Fig. 1a, b). Ten microliter rhizome extract of Z. officinale also significantly repelled males and females, the mean time spent and the mean number of entries made being respectively (0.94 min, p<0.0001), (2.5, p<0.0001) and (0.81 min, p<0.0001), (2.6, p<0.0001) than controls (Fig. 1c, d).
When 10 μL Z. officinale or A. melegueta vacuum-distilled extract loaded on filter paper disc was incorporated with 5 g winter wheat seeds and tested against R. dominica, significant repellency was also observed.
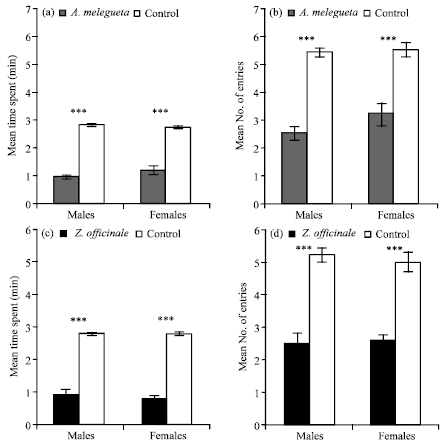 | |
| Fig. 1: | Mean time spent (1a) and mean number of entries made (1b) to 10 μL A. melegueta and mean time spent (1c) and mean number of entries made (1d) to 10 μL Z. officinale tests out of a total of 10 min by R. dominica in response to vacuum- distilled extracts from the plants in an airflow olfactometer assays. Error bars = SEM, N = 12 |
With A. melegueta, males were significantly repelled in the mean time spent (1.78 min, p = 0.007) and mean number of entries made (3, p<0.05); and females were also repelled as seen in the mean time spent (1.74 min, p<0.001) and mean number of entries or visits made (3.1, p<0.001) compared with the control (Fig. 2a). Z. officinale rhizome extract also exhibited repellent effect against both male R. dominica in the mean time spent (1.52 min, p = 0.003) and mean number entries made (2.8, p<0.001); and females in the mean time spent (1.49 min, p = 0.023) and mean number of entries made (2.7, p = 0.007) compared with the controls (Fig. 2b).
The use of botanical extracts in the control of stored products insect pests is an ancient practice (Qi and Burkholder, 1981). Oils are commonly used in insect control because they are relatively bioactive against virtually all life stages of insects (Adedire, 2003; Arannilewa et al., 2006).
The repellency bioassay of plant extracts on adult male and female R. dominica has been examined with A. melegueta and Z. officinale essential oils. Steam-distilled essential oils from these plants may have been very potent because of the strong pungent odours they produce and which may have exerted repellent effect on adult R. dominica as the beetles exhibited avoidance behaviour and limited their entries or visits to treated olfactometer arms. The oils retained their repellency against R. dominica even when incorporated with 5 g winter wheat kernels, a known host plant of the insect.
Plant volatiles provide reliable cues enabling herbivores to distinguish host and non-host plants (Bernays and Chapman, 1994). These behaviours potentially confound the benefits to pest management strategies of using repellents and non-host volatiles that interfere with selection of host plants. Non-host volatiles can be used to mask host odours or evoke non-host avoidance and repellent behaviours (Ukeh and Umoetok, 2007).
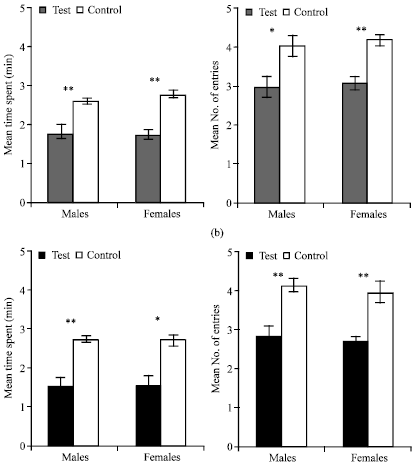 | |
| Fig. 2: | Responses of adult R. dominica to 10 μL A. melegueta seed extract +5 g wheat grains (2a) and 10 μL Z. officinale rhizome extract +5 g wheat grains (2b) in a four-arm olfactometer under 10 min observation period. Error bars = SEM, N = 12 |
The repellent action of the oils of A. melegueta and Z. officinale may be caused by the cumulative effect of most of the compositionally significant compounds of these plants. The composition of the volatile oils from the rhizomes and seeds of these plants have been investigated: the major components of Z. officinale rhizome are dominated by monoterpenoids and sesquiterpenoids including, β-phellandrene, β-pinene, α-pinene, camphene, β-mycrene, geranyl isobutyrate, α-muurolene, α-zingiberene, α-farnesene, β-sesquiphellandrene, butanol, p-cineole and 3,7-dimethyl-1,6-octadien-3-ol (Shao et al., 2003; Gong et al., 2004). Ajaiyeoba and Ekundayo (1999) reported that 2 sesquiterpene hydrocarbons, α-humulene and β-caryophyllene and their oxides were the major constituents of the hydrodistillation volatile oil of A. melegueta from Nigeria, while monoterpenes accounted for only 1% of the volatile oil. But the hexane and methanol extracts of A. melegueta seed yielded gingerol, zingiberone, paradol, trans-6-shogaol, cis-isoelemicin, β-bisabolene, α-guaiene, aromadendrene, trans-β-farnesene and geraniol (Ntonifor et al., 2006).
Plant metabolites and essential oils have been reported to show various forms of bioactivities against arthropods. For example, Ndungu et al. (1995) reported the repellency of the essential oil of the shrub Cleome monophylla against the livestock tick, Rhipicephalus appendiculatus and the maize weevil, Sitophilus zeamais which, at the highest dose, was comparable to that of the commercial arthropod repellent N,N-diethyl toluamide (DEET). Mason and Linz (1997) reported the activity of garlic Allium sativum as a repellent, antifeedant, bactericide, fungicide and nematicide. Similarly, Tapondjou et al. (2005) reported the repellent and toxic activities of essential oils extracted from the leaves of Eucalyptus saligna and Cupressus sempervirens against S. zeamais and Tribolium confusum, as both oils considerably reduced the F1 progeny production and grain weight loss. Ukeh and Umoetok (2007) had earlier on reported the repellency of West African black pepper, Piper guineense and African nutmeg Monodora myristica seed volatiles alone and in combination with different host plant grains against R. dominica in a four-arm airflow olfactometer. The essential oil of lavender, Lavendula angustifolia has been reported as a potential repellent for the pollen beetle Meligethes aeneus (Cook et al., 2007).
These studies indicate that A. melegueta seed and Z. officinale rhizome vacuum-distilled oils have some potential for use as repellents against R. dominica. During these trials, no death of the beetles was recorded. Investigation is presently going on to identify which of the active compounds are responsible for repellency of these plants and the mode of action of the compounds.
ACKNOWLEDGMENTS
The author is grateful to Professor Jenny Mordue of the School of Biological Sciences, University Aberdeen, UK for supervision, Professor John Pickett and Dr. Mike Birkett of Biological Chemistry Department, Rothamsted Research, England for facilities used for vacuum distillation and on-going GC-MS of the plant extracts. This research was funded by Commonwealth Scholarships.
REFERENCES
- Adedire, C.O., 2002. Use of nutmeg Myristica fragrans (Houtt.) powder and oil for the control of cowpea storage bruchid, Callosobruchus maculatus Fabricius. J. Plant Dis. Plant Prot., 109: 193-199.
Direct Link - Ajaiyeoba, E.O. and O. Ekundayo, 1999. Essential oil constituents of Aframomum melegueta (Roscoe) K. Schum. Seeds (alligator pepper) from Nigeria. Flavour Fragrance J., 14: 109-111.
CrossRefDirect Link - Arannilewa, S.T., T. Ekrakene and J.O. Akinneye, 2006. Laboratory evaluation of four medicinal plants as protectants against the maize weevil, Sitophilus zeamais (Mots). Africa J. Biotechnol., 5: 2032-2036.
Direct Link - Bashir, T., L.A. Birkinshaw, D.R. Hall and R.J. Hodges, 2001. Host odours enhance the responses of adult Rhyzopertha dominica to male‐produced aggregation pheromone. Entomol. Exp. Applied, 101: 273-280.
CrossRefDirect Link - Cook, S.M., M. Jonsson, M.P. Skellern, D.A. Murray, P. Anderson and W. Powell, 2007. Responses of Phradis parasitoids to volatiles of lavender, Lavendula angustifolia-a possible repellent for their host, Meligethes aeneus. Biocontrol, 52: 591-598.
CrossRefDirect Link - Edde, P.A., T.W. Phillips and M.D. Toews, 2005. Responses of Rhyzopertha dominica (Coleoptera: Bostrichidae) to its aggregation pheromones as influence by trap design, trap height and habitat. Environ. Entomol., 34: 1549-1557.
Direct Link - Ghorpade, K.D. and K.S. Thyagarajan, 1980. A reliable character for sexing live or dead Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res., 16: 151-153.
CrossRefDirect Link - Gong, F., Y.S. Fung and Y.Z. Liang, 2004. Determination of volatile components in ginger using gas chromatography-mass spectrometry with resolution improved by data processing techniques. J. Agric. Food Chem., 52: 6378-6383.
Direct Link - Hagstrum, D.W., 2001. Immigration of insects into bins storing newly harvested wheat on 12 Kansas farms. J. Stored Prod. Res., 37: 221-229.
Direct Link - Landolt, P.J., 1997. Sex attractant and aggregation pheromones of male phytophagous insects. Am. Entomol., 43: 12-22.
CrossRefDirect Link - Mason, J.R. and G. Linz, 1997. Repellency of garlic extract to European starlings. Crop Prot., 16: 107-108.
CrossRefDirect Link - Ndungu, M.W., W. Lwande, A. Hassanali, L. Moreka and S.C. Chhabra, 1995. Cleome monophylla essential oil and its constituents as tick (Rhipicephalus appendiculatus) and maize weevil (Sitophilus zeamais) repellents. Entomol. Exp. Appl., 76: 217-222.
CrossRefDirect Link - Ntonifor, N.N., I. Mueller-Harvey, H.F. van Emden and R.H. Brown, 2006. Antifeedant activities of crude seed extracts of tropical African species against Spodoptera littoralis (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci., 26: 78-85.
Direct Link - Qi, I.T. and W.E. Burkholder, 1981. Protection of stored wheat from the granary weevil by vegetable oils. Econ. Entomol., 74: 502-505.
CrossRefDirect Link - Shao, Y., P. Marriott, R. Shellie and H. Hugel, 2003. Solid-phase micro-extraction: Comprehensive two-dimensional gas chromatography of ginger (Zingiber officinale) volatiles. Flav. Fragr. J., 18: 5-12.
Direct Link - Tapondjou, A.L, C. Adler, D.A. Fontem, H. Bouda and C. Reichmuth, 2005. Bioactivities of cymol and essential oils of Cupressus sempervirens and Eucalyptus saligna against Sitophilus zeamais Motschulsky and Tribolium confusum du Val. J. Stored Prod. Res., 41: 91-102.
Direct Link - Ukeh, D.A. and S.B.A. Umoetok, 2007. Effects of host and non-hosts plant volatiles on the behaviour of the lesser grain borer, Rhyzopertha dominica (Fab.). J. Entomol., 4: 435-443.
CrossRefDirect Link - Vela-Coiffier, E.L., W.S. Fargo, E.L. Bonjour, G.W. Cuperus and W.D. Warde, 1997. Immigration of insects into on-farm stored wheat and relationships among trapping methods. J. Stored Prod. Res., 33: 157-166.
CrossRefDirect Link