
J.A. Timbilla
Crops Research Institute, P.O. Box 3785, Kumasi, Ghana
K. Yeboah-Gyan
Department of Theoretical and Applied Biology,
Rwame Xlcrumah University of Science and Technology, Rumasi, Ghana
B.W. Lawson
Department of Theoretical and Applied Biology,
Rwame Xlcrumah University of Science and Technology, Rumasi, Ghana
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 2 | Page No.: 103-112







ABSTRACT
The increasing importance of dry season populations of the African polyphagous grasshopper, Z. variegatus as a pest in agriculture and forestry has been attributed to the sequestration of pyrrolizidine alkaloids (PAs) from the flowers of the exotic noxious weed Chromolaena odorata for defense against natural enemies and protection of its diapausing eggs. This phenomenon is, however, seen as a novel opportunity to lure the insect with PAs for the development of PA-based attracticides for its management. This, notwithstanding, there is no empirical data on the efficacy of the available PA containing plants and the stage (s) of Z. variegatus worth targeting for the development of an integrated management strategy. Four categories of the growth stages of Z. variegatus were evaluated for the extent of migration to the roots of C. odorata, Heliotropium indicum and Crotalaria retusa which are plants containing PAs. Subsequently, fifth instar hoppers of Z. variegatus were evaluated for their attraction to the dry and fresh roots and flowers of C. odorata with a blank control in the laboratory. The results showed that 300 g of the dry chopped roots of C. odorata hold promise for use as PA-lures for the development of PA-based attracticides. The 3rd to 6th instar larvae of Z. variegatus have the highest degree of attraction to PAs. Also, the roots of C. odorata stored for a year are effective lures for the grasshopper while the flowers lose their attractive principle after 24 h. The results raise hope for the cost efficient and sustainable management of the grasshopper to salvage the agriculture and timber industries in Ghana and the sub region.
PDF Abstract XML References Citation
How to cite this article
J.A. Timbilla, K. Yeboah-Gyan and B.W. Lawson, 2008. Attraction of Zonocerus variegatus (Orthoptera: Pyrgomorphidae) to Pyrrolizidine Alkaloids: A Potential Novel Approach to its Management. Journal of Entomology, 5: 103-112.
DOI: 10.3923/je.2008.103.112
URL: https://scialert.net/abstract/?doi=je.2008.103.112
DOI: 10.3923/je.2008.103.112
URL: https://scialert.net/abstract/?doi=je.2008.103.112
INTRODUCTION
The variegated grasshopper, Zonocerus variegatus (L.) (Orthoptera: Pyrgomorphidae) is a serious pest of agriculture and forestry in sub Saharan Africa (Baker et al., 1977; Chapman et al., 1986; Timbilla, 2006) due to among other factors, the introduction of the weed, Chromolaena odorata (L.) R.M. King and H. Robinson (Asteraceae: Eupatoriae).
Recent chemo-ecological studies have revealed a pharmacophagous relationship between Z. variegatus and C. odorata (Boppré, 1991; Timbilla, 2006) where the insect sequesters pyrrolizidine alkaloids (PAs) from the flowers of the weed (Biller et al., 1994) for defense of its dry season populations against antagonists (Bernays et al., 1977; Chapman et al., 1986; Boppré and Fischer, 1994; Biller et al., 1994; Fischer and Boppré, 1997; Idowu and Modder, 1998). The above phenomenon, however, leads to a novel opportunity for the management of the insect using the PAs in C. odorata as lures. The successful use of C. odorata in the management of Z. variegatus would be contributing, to a lesser extent, to the integrated management of the weed.
The present study therefore sought to determine whether the PAs in C. odorata could serve as effective lure for Z. variegatus.
MATERIALS AND METHODS
Rearing of Zonocerus variegatus in the Laboratory
Field collections of first instar hoppers of Z. variegatus were made with an entomological net and maintained with cassava foliage according to the method described by Cobbinah and Tuani (1992) in insectary cage measuring 2x2x3 m, thoroughly cleaned every other day and fresh feed provided.
Attraction Experiments
Hundred grams each of the dry roots of C. odorata, Crotalaria retusa (Fabaceae) and Heliotropium indicum (Boraginaceae) air dried for eight hours (serving as PA-sources) were spread in wooden trays measuring 0.3x0.3x0.02 m together with an empty wooden tray of the same dimensions as control. The contents of each tray were covered with a nylon mesh to prevent Z. variegatus from feeding on the roots.
Each test material, serving as treatment was placed a meter away from the test hoppers confined to one end of the insectary cage. Counts of Z. variegatus attracted to each treatment were made at 30 min intervals using four categories of the developmental stages of Z. variegatus viz., 1st and 2nd, 3rd and 4th, 5th and 6th instar nymphs and adults as the insects grew and repeated 16 times each in a randomized complete block design.
Following the above experiments, the dry roots of C. odorata were selected as a candidate lure for Z. variegatus and subsequently, specific attraction experiments were conducted on the relative efficacy of the PAs from the flowers and roots of C. odorata, efficacy of pulverized and chopped roots of C. odorata and the determination of least effective quantity of dry chopped roots of C. odorata as attractants of Z. variegatus. Here, the 3rd-5th instar larvae were used for the attraction experiments. Trays of similar dimensions described above (0.3x0.3x0.02 m) were used for each respective treatments i.e., flowers versus roots of C. odorata, pulverized versus chopped roots of C. odorata and for the determination of least effective quantity of chopped roots of C. odorata.
A choice test using 300 g dry chopped roots of C. odorata (as the PA-source), cut stems of cassava (about 30 cm) as feed and an empty tray as control were evaluated for the diurnal behavioural pattern of Z. variegatus with respect to its attraction to sources of PAs.
There were thus three treatments, namely:
| • | Dry chopped roots of C. odorata. |
| • | Cut cassava stems. |
| • | Control (Empty tray). |
The experiment was replicated three times in a randomized complete block design with about 200 4th instar nymphs of both sexes of Z. variegatus. Each experiment lasted from 7:00 am to 5:00 pm for each replication. Here, the insects had the choice to consume the dry roots of C. odorata as PA-source and the cassava stems provided as feed to simulate field conditions.
The diurnal behavioural pattern of Z. variegatus with respect to the dry chopped roots of C. odorata (acting as a source of PAs) was further evaluated in a citrus field (acting as feed) and a blank control treatment. The field experiment was replicated 4 times in a randomized complete block design, each replication lasting from 7:00 am to 5:00 pm each day.
Data Analysis
The experiment on plant roots containing PAs evaluated with respect to stages of Z. variegatus was analysed using the Two-Way Analysis of Variance to study any interaction between the PA-sources and insect stages. The rest of the data were analysed using One -Way ANOVA. All count data were square root transformed with the formula y = √(x + 0.5) using GraphPad Prism 4.00 for Windows statistical package for analysis of variance (GraphPad Prism 4.00, 2003).
RESULTS
The Effect of Pyrrolizidine Alkaloids Produced from Different Plant Roots as Attractants for Zonocerus variegatus in the Laboratory
The results did not show any interaction between the PA-sources and the stages of Z. variegatus tested (Fig. 1, 2). There were however, significant differences observed within the PA-plant roots and the insect stages evaluated. In experiment one, the relative mean proportion of attraction of Z. variegatus per root treatment were 34.2, 19.8 and 38.3% in respect of C. odorata, C. retusa and H. indicum and these were all significantly higher in attracting the hoppers than the Control treatment of 7.7% (p<0.05) (Fig. 1). The attraction of Z. variegatus to the dry roots of C. odorata and H. indicum were the same (p<0.05) and both significantly attracted hoppers better than the effect of the dry roots of C. retusa (p<0.05) (Fig. 1a).
In experiment two, all the PA-lures tested were significantly better in attracting Z. variegatus than the Control which recorded a proportion of 2.3% insects (Fig. 1b). Among the PA treatments evaluated, the mean proportion of attraction of Z. variegatus to the dry roots of H. indicum was 46.2%. This was significantly higher than the remaining two PA treatments (p<0.05). The performance of the dry roots of C. odorata as a lure for the hoppers was also significantly more efficient (33.5%) than the dry roots of C. retusa (18.0%) (p<0.05) (Fig. 1).
With respect to the attraction of 3rd and 4th, 5th and 6th and the adults of Z. variegatus to the PAs plants, no significant differences were observed in experiment one (p<0.05) with values 34.4, 26.8 and 28.8% of insects, respectively but all were significantly attracted to the PA-plants than the 1st and 2nd instars of 12.0% of insects (p<0.05) (Fig. 2).
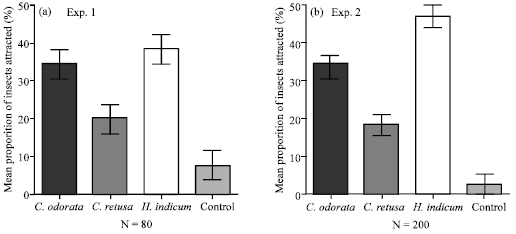 | |
| Fig. 1: | Relative attraction of all stages of Z. variegatus to dry roots of C. odorata, C. retusa and H. indiacum (sources of pyrrolizidine alkaloids) and a blank control in the laboratory. Bars indicate standard error of means |
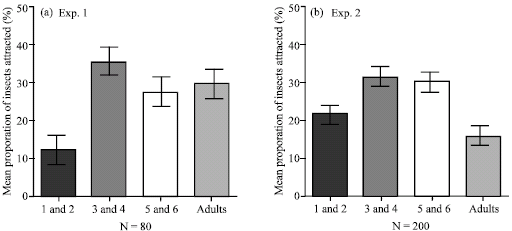 | |
| Fig. 2: | Relative attraction of four categories of the developmental stages of Z. variegatus to pyrrolizidine alkaloid sources in the laboratory. Bars indicate standard error means |
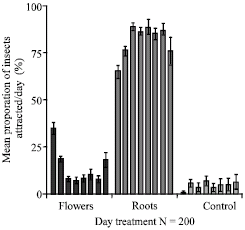 | |
| Fig. 3a: | Relative progression of attraction of 3rd to 5th instar larvae of Z. variegatus to flowers and roots of C. odorata. Bars indicate standard error of means |
In experiment two (Fig. 2a, b), the mean proportion of attraction to the PA-plants tested by the 3rd and 4th and 5th and 6th nymphal stages of Z variegatus were the same (p<0.05) i.e., 31.6 and 30.6%, respectively. These values were however significantly higher in luring Z. variegatus than the 1st and 2nd nymphs and adults which were also significantly the same with proportional values of 21.7 and 16.1% of the insects, respectively (p<0.05) (Fig. 2).
The Relative Efficacy of the Pyrrolizidine Alkaloids Produced from the Flowers and Roots of Chromolaena odorata as Attractants for Zonocerus variegatus
The relative attraction of Z. variegatus to the fresh roots of C. odorata was significantly higher (64.9%) than the fresh flowers (34.5%) (p<0.05) (Fig. 3a). The performance of the roots of C. odorata in attracting Z. variegatus ranged from 64.9 to 88.8% of hoppers within the period and these were all significantly higher than the performance of the flowers and the blank control compared over the same period. With the exception of fresh flowers and those dried for one and seven days, the performance of the flowers for the remaining days were not statistically different from the blank control (p<0.05) (Fig. 3a).
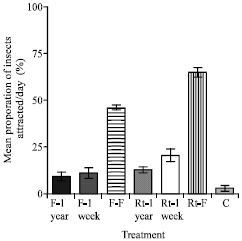 | |
| Fig. 3b: | Mean attraction of 3rd to 5th instar larvae of Z. variegatus to flowers and roots of C. odorata compared with those dried for a week and year. Bars indicate standard error of means. F-1 year designated as flowers of C. odorata dried for one year, F-1 week as flowers of C. odorata dried for a week, F-F as fresh flowers, Rt-1 week and Rt-1 year as roots of C. odorata dried for a year and week, respectively, Rt-F as fresh roots and C as Control |
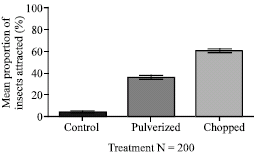 | |
| Fig. 4: | Relative attraction of third to fifth instar larvae of Zonocerus variegatus to pulverized and chopped roots of Chromolaena odorata formulations in the laboratory. Bars indicate standard error of means |
In another experiment comparing fresh treatments and those stored for up to a year, the results showed that the fresh roots of C. odorata were significantly better in luring Z. variegatus (65.0% of hoppers) than the fresh flowers (46.4% of hoppers).
The next was the roots dried for a week, recording 20.8% of hoppers (Fig. 3b). There was no significant difference between the roots dried for a year and flowers dried for a week and a year, but these were all significantly higher in attracting hoppers than the control.
Efficacy of Pulverized and Chopped Roots of Chromolaena odorata as Pyrrolizidine Alkaloid Lures for the Attraction of Zonocerus variegatus in the Laboratory
The results on the relative attraction of Z. variegatus to the pulverized and chopped root formulations of C. odorata showed that the effect of the dry chopped root formulation was statistically more efficacious than the pulverized root formulation with values of 59.9 and 36.4% of hoppers, respectively p<0.05) (Fig. 4). These values were both statistically better in attracting hoppers than the blank control treatment of 3.7% of hoppers (p<0.05).
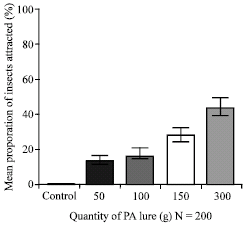 | |
| Fig. 5a: | Attraction of 3rd to 5th instar larvae of Z. variegatus to different qualities of chopped roots of C. odorata in the laboratory. Bars indicate standard error of means |
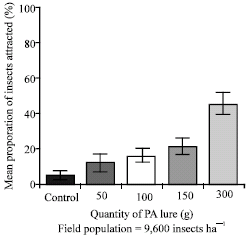 | |
| Fig. 5b: | Attraction of 3rd to 5th instar larvae of Z. variegatus to different quantities of dry roots of C. odorata in the field. Bars indicate standard error of means |
The Determination of Least Effective Quantity of Dry Chopped Roots of Chromolaena odorata as Pyrrolizidine Alkaloid Source on the Attraction of Zonocerus variegatus
In the laboratory, all the PA treatments were significantly higher than the blank control in attracting hoppers (Fig. 5a). Using 300 g of dry chopped roots of C. odorata to lure hoppers in the laboratory attracted a mean proportion of 43.3% hoppers which was significantly better than using 50, 100 and 150 g where mean values of 12.4, 16.0 and 27.8% of hoppers were recorded, respectively (p<0.05). There was no significant difference when 50 or 100 g of the dry chopped roots of C. odorata were used in attracting Z. variegatus (p<0.05) but these were significantly lower than using 150 g of the root material.
The results of a field evaluation of the same quantities of 50, 100, 150, 300 g and a control yielded similar results (Fig. 5b). All the treatments were significantly higher in attracting hoppers than the control which recorded a mean proportion of 4.6% hoppers. Using 300 g of dry chopped roots of C. odorata in attracting Z. variegatus in the field was significantly better (45.1% of hoppers) than the smaller quantities of 150, 100 and 50 g which were statistically the same with values of 21.7, 16.6 and 12.1% of hoppers, respectively (p<0.05) (Fig. 5b).
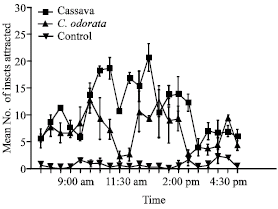 | |
| Fig. 6: | Mean diurnal behaviour of third to fifth instar larvae of Z. variegatus to cassava and dry roots of C. odorata in the laboratory. Bars indicate standard error of means |
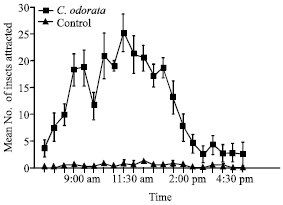 | |
| Fig. 7: | Mean diurnal pattern of 3rd and 4th instar larvae of Z. variegatus to dry roots of C. odorata in a fallow field. Bars indicate standard error of means |
Diurnal Attraction of Zonocerus variegatus to Dry Chopped Roots of Chromolaena odorata
In the choice laboratory experiment which compared the attraction of Z. variegatus to the dry roots of C. odorata (as PA-source) and cassava (as feed) the results are presented (Fig. 6). The attraction of Z. variegatus to each of the treatments per day varied with time and the peaks recorded for the cassava and C. odorata generally alternating, each recording about six peaks during the experimental period. Significantly, more hoppers were attracted to the feed at 8:00am, between 10:00 and 10:30 am, at 12:30 pm and 3:30 pm (Fig. 6). The highest attraction of the insect to the PA-source used was between 7:30 and 9:30 am, 12:00 pm, 1:00 pm and 4:30 pm (Fig. 6).
The effect of the dry chopped roots of C. odorata as PA-lure on the diurnal pattern of Z. variegatus studied in the field revealed that the insect is continuously attracted to PAs during the day (Fig. 7). The mean diurnal behavioural pattern indicated that the attraction of Z. variegatus to PAs in the field was similar to the observations made in the laboratory and more prolonged from about 9:00 am to 2:00 pm.
DISCUSSION
The dry roots of C. odorata and H. indicum emerged as promising PA-lures for Z. variegatus. Considering the abundance and menace caused by C. odorata, any positive use of the plant, particularly the roots in insect pest management will be contributing (to a lesser extent) to its reduction in the integrated management context.
The effect of PAs in luring Z. variegatus as unravelled in the present study is novel and different from the effect or action of many commercial lures such as food or sex pheromone lures (Butler and Las, 1983; Haynes et al., 1986; Rice and Kirsch, 1990; Charmillot et al., 2000; Krupke et al., 2002; Meagher Jr., 2002; Evenden and Mclaughlin, 2004) where the target organism is attracted to the lure source by a food or sex drive without much control. This is the first case of a lure for a grasshopper which could lead to a novel way of managing the insect in the Integrated Pest Management (IPM) context.
The results also indicated that Z. variegatus responds significantly to the fresh roots (dried for only six hours) and flowers of C. odorata than their corresponding dried forms, however, the flowers tend to lose their attractive principle after detachment from the plant. The low attraction of hoppers to the dry flowers was probably due to the rapid release of volatiles due to drying and cannot be used as effective lures for Z. variegatus. Modder (1984) suggested the use of fresh flowers for trapping Z. variegatus as part of a management strategy but this is not feasible since the insect is usually not found in dense populations on individual flowers apart from the vast distribution of the weed in areas where it has colonized.
The study has confirmed that all stages of Z. variegatus are attracted to PAs (Modder, 1984, 1986; Chapman et al., 1986; Boppré and Fischer, 1994; Fischer and Boppré, 1997) and that the 3rd and 4th instar hoppers are most attracted. This attribute of the insect has positive implications on management. The 3rd to 5th instar nymphs resulting from early hatchlings of August are most abundant in the field from September to mid November, a period when damage by the insect to dry season crops, particularly young cassava and vegetables is highest. Thus management targeted at these stages of Z. variegatus raises hope for mitigating its menace in the dry season by removing a substantial part of the dry season population. Management of hoppers during this period would also avoid competition with the flowers of C. odorata present from mid-November to February.
The results have also shown that the dry chopped roots of C. odorata as a PA-lure has an overall sustaining power in attracting hoppers than the pulverized form, probably because the latter gets depleted of volatiles within a short period due to complete breakdown of the vacuoles which contain the PA molecules.
The observations made indicate that Z. variegatus is attracted to all the PA-sources and that attraction of the insect to PAs is continuous throughout the day with peak response periods between 9:00 am and about 3:00 pm.
The results of the choice test on the diurnal behaviour shows that Z. variegatus will go after PAs irrespective of its primary food plants and this attribute of the insect is a positive indication for its management in the field with PA-lures. The higher affinity of Z. variegatus for its feed than PAs tend to give more weight to the assertion that PAs are not food for the insect (Modder, 1984, 1986; Boppré, 1991; Boppré and Fischer, 1994; Fischer and Boppré, 1997).
The results also suggest that the odour plumes resulting from the breakdown of PA molecules are constantly released from the PA-sources into the air and the first peaks attained by Z. variegatus, to the PAs sources between 8:00 and 9:00 am agrees with the observations of Modder (1984) who indicated that the activity of Z. variegatus starts at about 9:00 am. The two to six peaks observed regarding the diurnal behavioural pattern of Z. variegatus to PAs acting as lures corroborates the work by Adu-Mensah (1994) who investigated the diurnal attraction of Z. variegatus to pure PAs.
The exact mechanism controlling the alternating peaks exhibited by Z. variegatus with respect to its attraction to PAs and its primary food is not known, though this was found to be irrespective of the physiological condition of the insect. From the literature, PAs are phago-stimulatory (Bernays et al., 2002a, b). Bernays et al. (2003) also observed that extensive feeding of larvae of Estigmene acrea (Lepidoptera) on plants rich in PAs results in a temporary inhibition in the responsiveness of its PA receptor up to two hours, allowing their larvae to leave the PA-source to seek out PA-free food plants. This observation is also exemplified in sensory responses to chemicals in some other Lepidoptera (Schoonhoven, 1969; Blaney et al., 1986).
Zonocerus variegatus sequester PAs similar to arctiid caterpillars (Boppré, 1990; Hartmann and Witte, 1995; Weller et al., 1999), however, the particular mechanisms responsible is yet to be investigated.
From the above, Z. variegatus is attracted to PAs all through the day and hold promise as effective PA-lures for the development of PA-based attracticides/baited traps for the management of Z. variegatus.
REFERENCES
- Bernays, E.A., J.A. Edgar and M. Rothschild, 1977. Pyrrolizidine alkaloids sequestered and stored by the aposematic grasshopper, Zonocerus variegatus. J. Zool., 182: 85-87.
CrossRefDirect Link - Bernays, E.A., R.F. Chapman and T. Hartmann, 2002. A highly sensitive taste receptor cell for pyrrolizidine alkaloids in the lateral galeal sensillum of a polyphagous caterpillar, Estigmene acrea. J. Comp. Physiol. A., 188: 715-723.
Direct Link - Bernays, E.A., R.F. Chapman and T. Hartmann, 2002. A taste receptor neuron dedicated to the perception of pyrrolizidine alkaloids in the medial galeal sensillum of two polyphagous arctiid caterpillars. Physiol. Entomol., 27: 312-321.
Direct Link - Bernays, E.A., D. Rodrigues, R. F. Chapman, M. S. Singer and T. Hartmann, 2003. Loss of gustatory responses to pyrrolizidine alkaloids after their extensive ingestion in the polyphagous caterpillar Estigmene acrea. J. Exp. Biol., 206: 4487-4496.
Direct Link - Blaney, W.M., L.M. Schoonhoven and M.S.J. Simmonds, 1986. Sensitivity variations in insect chemoreceptors: A review. Experientia, 42: 13-19.
CrossRefDirect Link - Boppre, M., 1990. Lepidoptera and pyrrolizidine alkaloids Exemplification of complexity in chemical ecology. J. Chem. Ecol., 16: 165-185.
CrossRefDirect Link - Butler, Jr. G.D. and A.S. Las, 1983. Predaceous insects: Effect of adding permethrin to the sticker used in gossyplure applications. J. Econ. Entomol., 76: 1448-1451.
CrossRefDirect Link - Chapman, R.F., W.W. Page and A.R. McCaffery, 1986. Bionomics of the variegated grasshopper (Zonocerus variegatus) in West and Central Africa. Ann. Rev. Entomol., 31: 479-505.
CrossRefDirect Link - Charmillot, P.J., D. Hofer and D. Pasquier, 2000. Attract and kill: A new method for control of the codling moth Cydia pomonella. Entomol. Exp. Appl., 94: 211-216.
Direct Link - Cobbinah, J.R. and G.K. Tuani, 1992. Antifeedant effect of Jatropha curcas L. seed oil and extracts on the variegated grasshopper, Zonocerus variegatus L. (Orthoptera: Arcridae). J. Univ. Sci. Tech., 12: 136-139.
CrossRef - Evenden, M.L. and J.R. Mclaughlin, 2004. Initial development of an attracticide formulation against the oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae). Environ. Entomol., 33: 213-220.
Direct Link - Fischer, O.W. and M. Boppre, 1997. Chemoecological Studies Reveal Causes for Increased Population Densities of Zonocerus (Orth.: Pyrgomorphidae) and Offer New Means for Management. In: New Strategies in Locust Control, Krall, S., R. Peveling and D. Ba Diallo (Eds.). Birkhauser-Verlag, Basel, Switzerland, ISBN-13: 9783764354428, pp: 267-282.
- GraphPad Software, 2003. GraphPad Prism Version 4.00. GraphPad Software Inc., San Diego, California, USA.
Direct Link - Haynes, K.F., W.G. Li and T.C. Baker, 1986. Control of pink bollworm moth (Lepidoptera: Gelechiidae) with insecticides and pheromones (Attracticide): Lethal and sublethal effects. J. Econ. Entomol., 79: 1466-1471.
CrossRefDirect Link - Krupke, C.H., B.D. Roitberg and G.J.R. Judd, 2002. Field and laboratory responses of male codling moth (Lepidoptera: Tortricidae) to a pheromone-based attract-and-kill strategy. Environ. Entomol., 31: 189-197.
Direct Link - Meagher, Jr. and L. Robert, 2002. Trapping noctuid moths with synthetic floral volatiles lures. Entomol. Exp. Appl., 103: 219-226.
Direct Link - Modder, W.W.D., 1984. The attraction of Zonocerus variegatus (L.) (Orthoptera: Pyrgomorphidae) to the weed Chromolaena odorata and associated feeding behaviour. Bull. Entomol. Res., 74: 239-247.
CrossRefDirect Link - Weller, S.J., N.L. Jacobson and W.E. Conner, 1999. The evolution of chemical defenses and mating systems in tiger moths (Lepidoptera: Arctiidae). Biol. J. Linn. Soc., 68: 557-578.
CrossRefDirect Link