
Vineet Kumar
Electron Microscopy Division, India
B.K. Kariappa
Central Sericultural Research and Training Institute, Srirampura, Mysore-570 008, India
A.M. Babu
Electron Microscopy Division, India
S.B. Dandin
Central Sericultural Research and Training Institute, Srirampura, Mysore-570 008, India
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 2 | Page No.: 68-81







ABSTRACT
The eggshell of the eri moth, Samia ricini (Donovan) (Lepidoptera: Saturniidae) was investigated by scanning electron microscopy. The surface of the egg chorion of eri moth, revealed the structural elements viz., the micropylar rosette surrounding the micropyle, micropylar canals, shell imprints, aeropyles and the regional differentiation at the different poles. The oval shaped eggs were measured 820-990 (878.72±21.23) μm in length. The highly decorated chorion of S. ricini had a micropylar rosette of 45-51 (48.0±1.02) μm diameter with a centrally located micropylar pit of 2.08-2.31 (2.21±0.036) μm diameter. The micropylar pit consists of seven micropylar openings, each micropylar opening ranged from 0.26-0.28 (0.27±0.003) μm in diameter. Each micropylar opening opens into the seven distinct micropylar canals. The micropylar apparatus encircled by 11-14 petal-shaped primary unequal cells. These cells ranged from 12.0-12.9 (12.48±0.13) μm in length forming an asymmetrical rosette. The secondary petal-shaped cells were short in length and measured 8.4-9.1 (8.72±0.12) μm surrounded the primary petal cells. The entire surface of the chorion had a reticulate pattern of pentagonal and hexagonal cells, each boarded by 4-6 aeropyles of diameter 0.22-0.25 (0.23±0.05) μm. Further, the sunken aeropyles were also observed for the first time in second hemisphere of the egg and measured 0.22-0.24 (0.23±0.003) μm in diameter.
PDF Abstract XML References Citation
How to cite this article
Vineet Kumar, B.K. Kariappa, A.M. Babu and S.B. Dandin, 2007. Surface Ultrastructure of the Egg Chorion of Eri Silkworm, Samia ricini (Donovan) (Lepidoptera: Saturniidae). Journal of Entomology, 4: 68-81.
DOI: 10.3923/je.2007.68.81
URL: https://scialert.net/abstract/?doi=je.2007.68.81
DOI: 10.3923/je.2007.68.81
URL: https://scialert.net/abstract/?doi=je.2007.68.81
INTRODUCTION
The insect eggs are characterized by an outer shell secreted by the follicular epithelium during the process of egg formation and provide strong and elastic mechanical protection to the developing embryo besides allowing air for respiration at the aeropylar regions. Another specialized area of the egg chorion called micropyle paves the way for the penetration of sperm (Leuckart, 1855). The morphology and architectural pattern of the egg chorion in different insects depend on the imprints of the follicular secretory cells on specialized regions such as micropyle, aeropylar, stripe and flat regions (Beament, 1948; Sakaguchi et al., 1973; Kafatos et al., 1977; Regier et al., 1980). The structures of insect eggshells are usually quite complex and a typical insect egg capsule consists of the vitelline envelope and chorion (Kumar et al., 2002a). The sculpturing of the outer part of envelope appears to be specific and is of considerable interest because of potential taxonomic applications (Rosciszewska, 1991). Further, the basic knowledge of insect eggshell structure and function, in particular of Lepidoptera, was provided by Leuckart (1855) and Korschelt (1887). Light Microscope (LM) and physiological studies of gas exchange and permeability (Beament, 1948; Wigglesworth and Beament, 1950) preceded a second phase of basic research that began with the establishment of transmission (TEM) and Scanning Electron Microscopy (SEM). Since then, the fine structure of a large number of lepidopteran eggs has been studied with particular emphasis on surface sculpturing (Matheny and Heinrichs, 1972; Mazzini, 1974; Downey and Allyn, 1980, 1981; Hill, 1982; Salkeld, 1983, 1984; Fehrenbach et al., 1987; Arbogast et al., 1989; Kumar et al., 1999; Kumar et al., 2002b; Kumar et al., 2003). The high resolution and 3-dimensional clarity of the scanning electron microscope make it an invaluable tool for studying diagnostic characters of insects. Hinton (1969) used scanning electron micrographs to illustrate respiratory systems of various insect eggshells. SEM studies revealed the egg surface to be regionally differentiated (Kafatos et al., 1977; Margaritis et al., 1980), also allowing a better distinction between closely related species than by Light Microscope (LM) (Arbogast et al., 1980).
The surface structure of lepidopteran eggs, as revealed by scanning electron microscopy, provides reliable characters for separation of species (Arbogast et al., 1989) and the external morphology of a wide variety of insect eggs has been examined using different techniques (Hinton, 1981). A taxonomy key has been formulated based on the egg chorionic characteristics and its architecture in sod webworms (Matheny and Heinrichs, 1972).
India is the only country, which produces all four kinds of silk viz., mulberry, eri, tasar and muga and the second highest country in the total production of silk, after China. The eri silk is produced by Samia ricini (Donovan) and is an economically important insect for sericulture point of view. Here a study has been taken on the surface ultrastructure on the egg chorion of eri silkworm, Samia ricini (Donovan) using scanning electron microscope. The results of the study reported here describing the surface ultrastructure of the egg chorion of Samia ricini (Donovan) may find use in the study of taxonomy and phylogeny of the insect.
MATERIALS AND METHODS
A laboratory culture of Samia ricini (Donovan) was established from adults that emerged from green pupae. Freshly emerged male and female adults were released in a cage (28x18 cm) and were provided with a 5% sucrose solution for feeding and fresh caster branches for egg layings. After mating, the freshly laid eggs were gently removed from the caster leaves using a fine tipped hair brush.
For scanning electron microscopic study, the eggs were fixed for two hours at room temperature in 2.5% glutaraldehyde prepared in 0.2 M cacodylate buffer (pH = 7.2), dehydrated in a graded alcohol-acetone series and dried in a critical point drier (EMS-850) using CO2 as the transition fluid. The dried samples were mounted onto copper stubs and coated with gold (20 nm thickness) in a Sputter coater (EMS-550) and examined using a JEM 100 CX II electron microscope fitted with ASID 4D attachments (JEOL Ltd., Tokyo, Japan) at 20 kV.
RESULTS AND DISCUSSION
Gravid females of Samia ricini laid eggs on the lower surface of its food plant leaves. The eggs were found deposited upright serially arranged in 6 to 36 clutches of 317 to 510 eggs. They were attached to the leaf with their posterior poles and among each other with their lateral sides, by a sticky secretion (Fig. 1-5). The freshly laid eri eggs were slight white in colour. As the embryo developed inside the egg, the colour of the shell changed from whitish to yellowish, yellowish to ashy and finally ashy to blackish just before hatching (Fig. 2). The eri eggs were of medium size compared to mulberry or muga egg and have oval shape. Fehrenbach et al. (1987) have investigated the eggs of three lepidopteran moths by scanning (SEM) and transmission (TEM) electron microscopy, which revealed that the eggs of Heliothis virescens were standing type (upright) and ca 550 μm in diameter and ca 600 μm in length, whereas the eggs of Spodoptera littoralis was also standing type and almost spheroid with a diameter of 500-550 μm and slightly less in length.
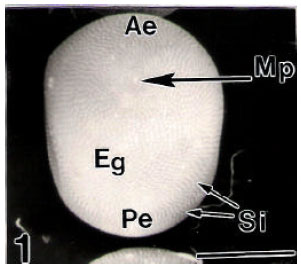 | |
| Fig. 1: | A single egg (Eg) of Samia ricini showing anterior end (Ae), posterior (Pe) and anteriorly located micropyle (Mp) and the shell imprints (Si) on the surface. (Scale bar = 190 μm) |
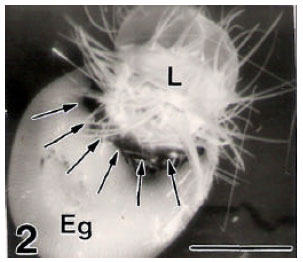 | |
| Fig. 2: | A neonate larva (L) emerges out through the anterior end. Arrows indicate the anterior cut surface of the egg (Eg). (Scale bar = 120 μm) |
However, the eggs of Cydia pomonella belonged to the lying (flat) type which measured ca 1350 μm in length and ca 1050 μm in width. Arbogast et al. (1989) studied the eggs of three tineid pests under scanning electron microscope and found the eggs of Tinea pallescentella Stainton as subcylindrical, 0.61-0.67 (0.64±0.02) mm long and 0.31-0.35 (0.33±0.01) mm in diameter at the widest point. But the eggs of Tinea occidentella Chambers was subcylindrical, 0.61-0.65 (0.62±0.01) mm long and 0.36-0.39 (0.38±0.01) mm in diameter at the widest point, bluntly rounded at both ends; the anterior end usually broader than the posterior. Though the eggs of Niditinea fuscella (L.) were also found subcylindrical their anterior end was bluntly rounded and the posterior end more acutely rounded, but occasionally ellipsoid, 0.42-0.50 (0.45±0.02) mm long with 0.26-0.30 (0.28±0.01) mm in diameter at the widest point (Arbogast et al., 1989). Hinton (1981) emphasized that in most Lepidoptera, the egg is a prolate or more or less spherical or rarely hemispherical.
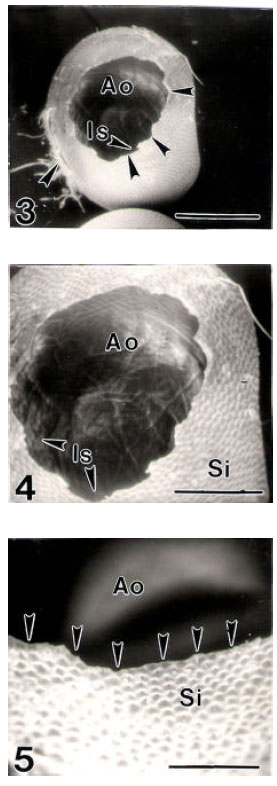 | |
| Fig. 3-5: | Vacated egg shows the anterior opening (Ao), which was cut open irregularly by the hatched larvae. Arrow head shows the inner surface (Is), The detached lateral side (Left sides: single arrow head in Fig. 3) is also shown with glued substances. (Scale bar = 120 μm for Fig. 3; 60 μm for Fig. 4 and 30 μm for Fig. 5) |
He also revealed that the size and shape of the eggs of certain insect like a tortricid, Rhyaciona duplana (Hub.) may also vary slightly according to the species of pine on which they are laid. In any insect eggshell there are three major layers viz., vitelline membrane, the endochorion and the third one is occasionally absent (Margaritis, 1985). The protection against environmental hazards during embryogensis is one of the functions of the eggshell.
The eggs of S. ricini measured 820-990 (878.72±21.23) μm in length. Matheny and Heinrichs (1972) have studied the egg chorion of 15 species of Lepidoptera moths and measured 0.598±0.002 μm in Thaumatopsis edonis, 0.425±0.009 mm in Chrysoteuchia topiaria and 0.537±0.009 μm in Pediasia trisecta. Recently, Kumar et al. (1999) observed the eggs of a lepidopteran moth, Spilarctia obliqua using electron microscope and measured 600-650 μm in diameter at the widest point. Further, Kumar et al. (2003) studied the fine structure of the egg of a lepidopterous moth, Amata passalis and measured the eggs 488±0.595 μm in diameter under scanning electron microscope.
The eggshell of S. ricini has a highly decorated chorion and its micropylar apparatus is located at the anterior pole of the egg (Fig. 1, 7-16). Hinton (1981) stated that the micropyles are without exception at the anterior pole of the lepidopteran eggs. Sakaguchi et al. (1973) have described the arrangement of follicular imprints of the micropylar and aeropylar regions of the egg chorion of Bombyx mori. In recent past many workers also observed the micropylar apparatus at the anterior end in the egg chorion of lepidopterous moths (Arbogast et al., 1989; Fehrenbach et al., 1987; Fehrenbach, 1989; Kumar et al., 1999, 2003). However, an unusual position of the micropylar structure was found on the lower surface of an egg in a lepidopterous moth Cydia pomonella (Fehrenbach et al., 1987). A micropylar rosette consisting of micropylar openings encircled with primary petal-shaped cells, which are followed by the secondary petal-shaped cells, was observed in S. ricini. The micropylar rosette of S. ricini ranged 45-51 (48.0±1.02) μm in diameter (Fig. 8). Fehrenbach et al. (1987) have observed a slightly bigger micropylar rosette ca 65 μm in diameter in Heliothis virescens, 55 μm in Spodoptera littoralis and about 50 μm in Cydia pomonella. A rosette of 11-14 petal-shaped primary cells surrounds the micropylar opening of S. ricini (Fig. 8-10, 13). The primary cells were again surrounded by a second row of 15-19 petal-shaped cells of unequal sizes (Fig. 8). The number of primary cells around the micropylar pit is 15-19 in A. passalis (Kumar et al., 2003), 10-14 in S. obliqua (Kumar et al., 1999), 5-7 in T. pallescentella, 5-8 in T. occidentella and 6-9 in N. fuscella (Arbogast et al., 1989).
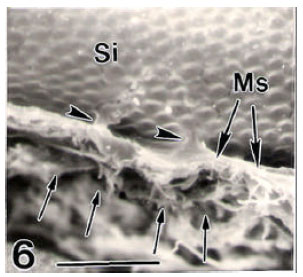 | |
| Fig. 6: | The posterior end of the egg showing its surface detached form host leaves (arrows). The membranous structure (Ms) which adheres to the leaf is connected to the egg by short vertical bars (arrow heads). (Scale bar = 30 μm) |
 | |
| Fig. 7-10: | General view of anterior egg pole revealing the rosette (Mr) of eleven primary petals |
However, in some eggs of N. fuscella the primary cells were found to be of subequal length forming symmetrical rosettes. Fehrenbach et al. (1987) described the fine structure of three lepidopteran moth eggs and explained that in Heliothis virescens, the micropylar rosette is composed of 13-15 petal-shaped primary cells with 4 or 5 micropylar canals opening into the micropylar plate, whereas in Spodoptera littoralis the number of the rosette petals varies between 6 and 11, 7 and 8 being the most frequent and with 3-4 micropylar openings present in the slightly depressed rosette center. In Cydia pomonella the micropylar rosette is composed of 8 or 9 rosette petals.
In a few eggs of S. ricini, the primary cells were subequal in length, arranged in a symmetrical rosette, but more often, some primary cells were longer than others and formed an asymmetrical rosette (Fig. 8 and 13). Similar asymmetrical rosettes of unequal petal-shaped cells have been reported in the eggs of six other lepidopteran moths viz., Tinea pallescentella, T. occidentella, Niditinea fuscella (Arbogast et al., 1989), Spilarctia obliqua (Kumar et al., 1999) and Amata passalis (Kumar et al., 2003).
The length of each primary rosette cells of S. ricini eggs ranged 12-12.9 (12.48±0.13) μm and some of the cells were observed with a protuberance on the distal end of the cell (Fig. 9-11). The secondary petal-shaped cells were found short in length and measured 8.4-9.1 (8.72±0.12) μm (Fig. 8 and 9).
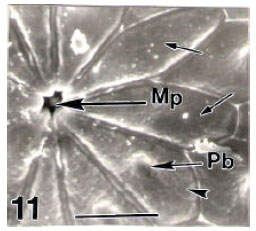 | |
| Fig. 11: | Arrow head shows the petal cell with a single protuberance (Pb) while the adjacent petal cells were observed without protuberance (arrows). (Scale bar = 3 μm) |
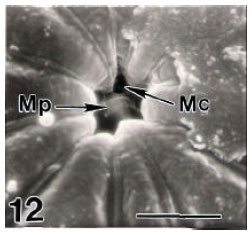 | |
| Fig. 12: | Top view of egg pole showing micropylar region entrally located micropyle (Mp) and seven micropylar canals (Mc). (Scale bar = 1 μm) |
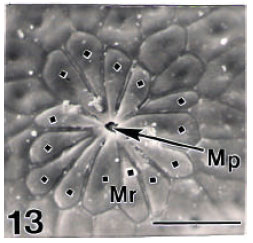 | |
| Fig. 13: | A top view of the other egg shows its micropylar rosette (Mr) with fourteen petal cells surrounding the micropyle (Mp). Scale bar = 7.5 μm) |
Each micropylar pit was 2.08-2.31 (2.21±0.036) μm in diameter and had seven distinct micropylar openings ranged from 0.26-0.28 (0.27±0.003) μm diameters and each micropylar opening radiates down into micropylar canal in micropylar plate in the eggs of S. ricini (Fig. 14-16). Kumar et al. (1999) studied 50 eggs of S. obliqua under SEM and found interestingly only a single egg had three micropylar openings whereas in all other eggs the micropylar apparatus had four micropylar openings (micropylae). Hinton (1981) described that in Lepidoptera; eggs have usually four micropyles, but may have more also. In eggs of 15 species of Notodontidae studied by him, the number of micropyles varied from 4 to 20. The number sometimes varies considerably in same species, e.g., in a few shells of Cerura vinula, the number of micropyles varied from 16 to 20. Study of micropyles has been a favorite subject since early times. Muller (1938) gave a very good description of the 4 micropyles of the pyralid, Plodia interpunctella and also described how they develop. Fehrenbach (1989) studied the fine structure of eggshells of 4 primitive moths and revealed that the egg chorion of Hepialis hecta and Wiseana umbracula had a different egg surface, with two micropylar openings at the opposite sides of the longer axis of the oval micropylar plate. In Mnesarchaea fusitella 2-3 oval micropylar openings were seen at the base of the micropylar rim. Matheny and Heinrichs (1972) studied the egg chorion of 15 species of sod webworm moth eggs (Lepidoptera) and developed a taxonomic key for egg identification.
In S. ricini the rosette of two rows of petal-shaped cells at the top and anterior pole of eggs is followed by the dense ridged shell imprints which cover rest of egg surface (Fig. 7-10, 19 and 22) except at the posterior pole where the egg was attached to the substratum (Fig. 6, 7, 20 and 21). Further, it was also observed that each shell imprint bears a centrally located blunt protuberance (Fig. 8-10). Though the posterior poles of egg sometime do not have the shell imprints but the blunt protuberances were observed in that area (Fig. 17). Similar to the observation on S. obliqua (Kumar et al., 1999), the shell imprints were mostly pentagonal or hexagonal in shape and measured 9.0-9.6 (9.24±0.10) μm in diameter in S. ricini. Generally it was found that 4 to 6 aeropyles of the size of 0.22-0.25 (0.23±0.05) μm were located on the ridges of each shell imprints, which transverse the shell down to the trabecular layer, on the anterior and middle region of the egg (Fig. 18 and 19), however, the posterior surface, which was observed with or without shell imprints, have the aeropyles in a pit (sunken aeropyles) (Fig. 20 and 21). The diameters of sunken aeropyles were measured 0.22-0.24 (0.23±0.003) μm. In S. ricini the total number of aeropyles on a single egg is too high to be counted. Matheny and Heinrichs (1972) have investigated the egg chorion of 15 species of sod webworm moths, using scanning electron microscope and revealed a smallest aeropyle i.e., 1.46±0.07 μm in Agriphila ruricollatta whereas the largest aeropyles were 3.03±0.16 μm in diameter in Crambus lequealellus. Fehrenbach et al. (1987) have reported about 50 aeropyles per egg measuring ca 1.9 μm in diameter in Heliothis virescens, ca 400 aeropyles per egg in Spodopters littoralis measuring about 0.9 μm wide, whereas the number of aeropyles was 140 per egg and ca 0.6 μm in diameter in the eggs of Cydia pomonella (Fig. 23 and 24). Arbogast et al. (1989) reported the aeropyles ranging from 0.67-1.20 (0.94±0.15) μm in T. pallescentella, 0.55-0.93 (0.77±0.04) μm in T. occidentella and 0.65-1.50 (1.10±0.36) μm in Niditinea fuscella. Recently, Kumar et al. (2003) observed the aeropyles of Amata passalis which measured 0.36±0.08 μm in diameter. However, the aeropyles were not observed on the first two rows of cells at the micropylar zone in S. ricini, as in the eggs of S. obliqua (Kumar et al., 1999) and S. littoralis (Fehrenbach et al., 1987). The panels of shell imprints adjacent to the micropylar zone in S. ricini have less number of aeropyles, whereas the shell imprints which are away from micropylar zone are completely boarded by aeropyles as in S. obliqua (Kumar et al., 1999). In T. occidentella aeropyles are distributed over the entire surface of egg whereas in T. pallescentella and N. fuscella they are restricted towards the anterior and posterior end of the eggs. (Arbogast et al., 1989). The number of aeropyles in S. obliqua varies from 18 to 47 on a single shell imprint; however, some large aeropyles were also observed which may be formed by the fusion of two adjacent small aeropyles (Kumar et al., 1999).
 | |
| Fig. 14-16: | The micropyle (Mp) reveals the distinct seven-micropylar openings (Mo), which further radiates down into the micropylar canal. (Scale bar = 2 μm for Fig.14; 1.2 μm for Fig. 15 and 0.4 μm for Fig. 16) |
 | |
| Fig. 17: | Posterior end (Pe) of an egg revealing a large number of protuberances (Pb) with out definite boundary of shell imprints. (Scale bar = 30 μm) |
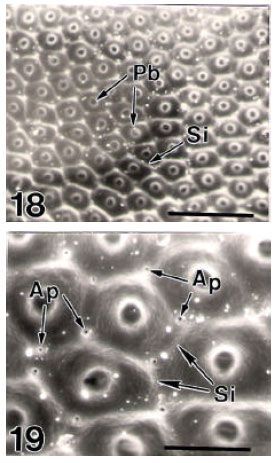 | |
| Fig. 18-19: | Middle region of an egg revealing the shell imprints (Si) and centrally located protuberance (Pb). The aeropyle were observed on the shell imprints (Si). (Scale bar = 20 μm for Fig. 18 and 6 μm for Fig. 19) |
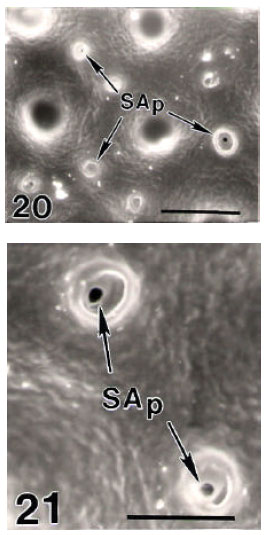 | |
| Fig. 20-21: | Sunken aeropyles (SAp) were observed on the posterior region of an egg. The sunken aeropyles were not found located on the shell imprints. (Scale bar =3 μm for Fig. 20 and 1.2 μm for Fig. 21) |
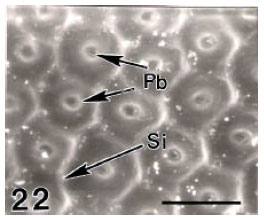 | |
| Fig. 22: | Extreme posterior end of the egg shows that each protuberance (Pb) is encircled by a shell imprints (Si). (Scale bar = 7.5 μm) |
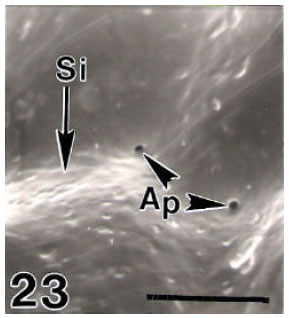 | |
| Fig. 23: | Two normal aeropyles (Ap) situated on the shell imprints (Si). (Scale bar = 2 μm) |
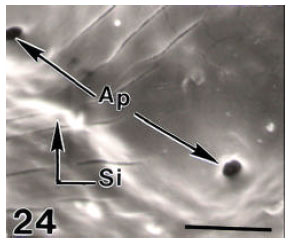 | |
| Fig. 24: | Magnified view of egg chorion reveals two normal aeropyles (Ap) on shell imprints (Si). (Scale bar = 0.6 μm) |
The main function of the aeropyles is to conduct ambient air into the trabecular layer (gas containing meshwork), which finally passes on to the oocyte. Very little is yet known about the permeability of the chorion. Tuft (1950) demonstrated that when the aeropyles of the eggs of Rhodnius were blocked with shellac, the egg continued to take up oxygen, though only at about a tenth of the normal rate. It may be that eggs depend not only upon oxygen entering through the aeropyles but also on that diffusing through the areas of chorion without aeropyles.
REFERENCES
- Kafatos, F.C., J.C. Regier, G.D. Mazur, M.R. Nadel and H.M. Balu et al., 1977. The Eggshell of Insects: Differentiation-specific Proteins and the Control of Their Synthesis and Accumulation During Envelopment. In: Results and Problems in Cell Differentiation, Beeman, W. (Ed.). Vol. 8, Springer Verlag, Berling, pp: 45-145.
- Kumar, V., M.N. Morrison, A.M. Babu and V. Thiagrajan, 2002. Egg shell architecture of the stink bug, Eocanthecona furcellata (Wolff.): Ultrastructure of micropylar processes and egg burster. Insect. Sci. Applic., 22: 67-73.
Direct Link
ASUTOSH KUMAR SRIVASTAVA Reply
Respected sirs,
an excellant work through SEM. As I am doing my Doctorate in Insect Endocrinology it become very use full for me.
i want to become in regular regular touch with you (all) sir so kindly forward e-mail id of yours.