
Omar Hamarsheh
Institute for Microbiology and Hygiene, Charite Campus Mittevon Humboldt University of Berlin, Dorotheenstrase 96, 1 01 1 7 Berlin, Germany
Sameer Barghouthi
Laboratoy of Leishmaniasis, Faculty of Medicine, Al-Quds University, Jerusalem, Palestine
Amer Al-Jawabreh
Department of Zoology, Faculty of Science, El Azhar University, Assiut, Egypt
Abdel Baset Zayed
Laboratoy of Leishmaniasis, Faculty of Medicine, Al-Quds University, Jerusalem, Palestine
Kifaya Azmi
Laboratoy of Leishmaniasis, Faculty of Medicine, Al-Quds University, Jerusalem, Palestine
Ahmad Amro
Laboratoy of Leishmaniasis, Faculty of Medicine, Al-Quds University, Jerusalem, Palestine
Ziad Abdeen
Laboratoy of Leishmaniasis, Faculty of Medicine, Al-Quds University, Jerusalem, Palestine
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 6 | Page No.: 425-434







ABSTRACT
Phlebotomine sand flies are weak flies, with maximum dispersal seldom exceeds one kilometer away from their breeding sites. For Phlebotomus papatasi, which has a broad geographical distribution, it is expected however, that the local populations are genetically isolated. The present study developed a Polymerase Chain Reaction (PCR) method to distinguish between the DNA profiles of two laboratory colonies and several field populations of P. papatasi sand flies originating from different sites in the West Bank. Primers, designed from the P. papatasi α-amylase gene, amplify population-specific bands in all male and female specimens tested. The profiles were based on the variability of PCR bands exhibited on agarose gels. The data analysis showed significant variations among the two colonies and eight field populations, the genetic variability evaluated by genetic distance indicated that colony populations, which originated from the Jordan Valley exhibit low genetic variability and are closely related to each other, whereas high genetic variation was detected among some populations originating from different sites in the West Bank. Population genetic analysis of sand fly populations from leishmaniasis-endemic areas may provide information on the amounts of genetic connectedness among P. papatasi populations distributed on large scale (i.e., among geographic regions) or within a region. These informations associated with estimation of their abilities to harbor and transmit Leishmania parasites will be necessary for developing effective control strategies.
PDF Abstract XML References Citation
How to cite this article
Omar Hamarsheh, Sameer Barghouthi, Amer Al-Jawabreh, Abdel Baset Zayed, Kifaya Azmi, Ahmad Amro and Ziad Abdeen, 2007. Genetic Variability of Sand Fly Phlebotomus papatasi Populations (Diptera: Psychodidae) Originating from the West Bank, Palestine. Journal of Entomology, 4: 425-434.
DOI: 10.3923/je.2007.425.434
URL: https://scialert.net/abstract/?doi=je.2007.425.434
DOI: 10.3923/je.2007.425.434
URL: https://scialert.net/abstract/?doi=je.2007.425.434
INTRODUCTION
The sand fly Phlebotomus papatasi (Diptera: Psychodidae) is a vector of Leishmania major, which is the causative agent of cutaneous leishmaniasis in the Palestinian Territories (Schlein et al., 1982; Jaffe et al., 2004; Nasereddin and Jaffe, 2004) and other regions in the world (Ashford and Bettini, 1987; Killick-Kendrick, 1990, 1999; Guernaoui et al., 2005; Yaghoobi-Ershadi et al., 2005). It also transmits other human diseases like sand fly fever (Lane, 1993). Sand flies, including P. papatasi, feed on sugar meals to obtain energy, without which they live only few days after emergence (Schlein and Jacobson, 1999; Müller and Schlein, 2004). The sand flies are able to select sugar meals from different sources including plants and aphid honeydew (Cameron et al., 1995; Müller and Schlein, 2004, 2005). It was shown that P. papatasi sand flies express α-amylase and α-glucosidase enzymes (Jacobson and Schlein, 2001). α-amylase activity was demonstrated in the salivary glands of the sand fly Lutzomyia longipalpis (Ribeiro et al., 2000; Cavalcante et al., 2006). On the genetic level, α-amylase is a multi copy gene in some insects (Maczkowiak and Da Lage, 2006); Drosophila ananassae has seven copies scattered on several chromosomal arms (Da Lage et al., 2003), Grossman et al. (1997) characterized the genomic structure of two α-amylase encoding genes in the Aedes aegypti. In case of sand flies, there is no available information on the α-amylase gene structure and the biological functions of sand fly and Leishmania glycosidases during their interaction remains an open and interesting field, since regular sugar meal can enhance the number of promastigotes developing in sand fly’s gut and also the number of parasites egested by infected flies while taking blood meal (Schlein and Jacobson, 2001). The presence of α-amylase and α-glucosidase enzymes enable P. papatasi to survive by digesting carbohydrates in the form of starch obtained from different plants (Schlein and Jacobson, 2000; Jacobson et al., 2001). This may lead to alter the distribution of P. papatasi populations in certain habitats. Therefore, the characterization of the genetic variation among populations of P. papatasi, will pave the way for understanding the relationship between the distribution of L. major strains, their reservoir hosts and their vectors.
The techniques most widely used for analysis of polymorphisms in sand flies are electrophoretic analysis of isoenzymes (Ward et al., 1981; Kassem et al., 1990, 1999; Fryauff and Hanafi, 1991; Perrotey et al., 2005), analysis of the internal transcribed spacer 2 (Hamarsheh et al., 2007) the use of microsatellite markers (Hamarsheh et al., 2006) and cytochrome b gene (Parvizi et al., 2003; Parvizi and Ready, 2006).
A polymerase chain reaction based technique known as Random Amplified Polymorphic DNA (RAPD-PCR) has been used for the identification of sand flies (Adamson et al., 1993) and polymorphisms are expressed as the presence or absence of a fragment of a particular size. It has also been used to investigate DNA polymorphism in L. migonei sand flies originated from three populations in Brazil and one laboratory colony from Venezuela (Silva, 2000). This technique produced useful genetic information and data generated by this approach have been used for population studies (Lanzaro and Warburg, 1995). This study discussed the utility of a simple and reliable PCR technique to characterize different P. papatasi populations originating from different areas in the West Bank.
MATERIALS AND METHODS
Sand Flies
The sand flies used in this study were collected from 10 sources; two laboratory colonies and eight field populations, all originating from different locations within the West Bank (32° 00 N, 35° 15 E).
Colony Sand Flies
The laboratory colonies (J and A) were already started in 1998 with sand flies collected in houses of an arid site close to Jericho, 15 km East of Jerusalem. The only difference between J and A colony is that the flies in the J colony were believed to be inbred; in which male and female P. papatasi individuals from other populations were introduced to the colony.
Sand fly colonies were reared at the laboratory, newly emerged sand flies were constantly offered 30% sucrose solution and water. For oviposition, female flies were offered a blood meal from an anaesthetized rabbit. Insectary conditions were 26±1°C, 80% relative humidity and 14:10 h light:dark photoperiod.
Field Collected Sand Flies
The field-collected sand flies were collected from different sites in the West Bank; Jericho (31° 52 0 N, 35° 27 0 E), Tubas ( 32° 19 17 N, 35° 22 9 E), Yabad (32° 26 42 N, 35° 9 57 E), Wadi Badan (32° 17 50 N, 35°21 0 E), Far'ah (32° 17 60 N, 35° 21 0 E), Salfit ( 32° 4 51 N, 35° 10 34 N), Attil (32° 23 7N, 35° 7 11 E) and Baqah (32° 12 18 N, 35° 6 56 E), the environmental characterization of the study areas are shown in Table 1.
| Table 1: | P. papatasi sample locations and environmental conditions of the study areas in the West Bank |
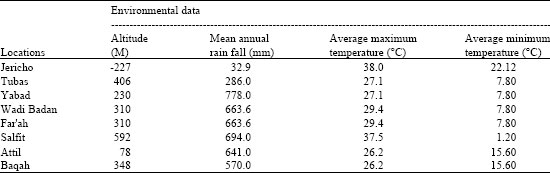 | |
The collections were carried out by means of CDC miniature light traps, between May and September in the year 2000. The map locations of the sand fly origins are shown in Fig. 1. The collected flies were transported live to the laboratory at Al-Quds University, Jerusalem, Palestine, immobilized using CO2 and stored at -70°C before being used for PCR work.
DNA Extraction
DNA was extracted from individual P. papatasi males and colony originated females by guanidine thiocyanate method (Boom et al., 1990) with slight modifications. Individual sand flies were ground dry in 1.5 mL autoclaved and UV radiated microfuge tubes, using micro-pestles (Ependorf, Germany). Each sample was then suspended in 0.5 mL of 4 M guanidine solution (0.1 M Tris-HCl pH 6.4, 0.02 M EDTA pH 8 and 1.3% Triton X-100) and incubated at 56°C under gentle agitation overnight. Samples were then boiled for 10 min, centrifuged for 5 min at 15,000 g and the supernatant was transferred to a new tube. One milliliter of 6 M sodium iodide solution and 10 μL of suspended silica beads were added to each tube mixed gently for 5 sec and incubated on ice for 1 h. The supernatant was removed, the pellet was washed twice with 500 μL of ice-cold washing buffer (0.2 M Tris-HCl pH 7.5, 1 M sodium chloride and 20 mM EDTA pH 8), then washed twice with 100% ethyl alcohol and the pellet was air dried. The DNA was suspended in 100 μL double distilled sterile water and incubated at 56°C for 1 h.
PCR Conditions
PCR amplifications were performed in 50 μL reaction volume containing 10 μL (about 50 ng) genomic DNA, 1.25 U recombinant Taq DNA polymerase (MBI Fermentus), 1x PCR buffer (750 mM Tris-HCl pH 8.8 at 25°C, 200 mM (NH4)2SO4 and 0.1% Tween-20), 4 mM MgCl2, 0.2 mM of deoxynucleotide mixture and 2 μM of AU 489 (5’-AAGCAGACTTTCATCCT-3’) and AL 997 (5’-CAGCTCCATGACCACG-3’) primers which were designed to amplify variable parts of the α-amylase gene (Hamarsheh, 2000). The mixtures were overlaid with one drop of mineral oil (Sigma). Amplification was performed using Personal Thermocycler (version 2.71bc, Biotron) and programmed as follows: DNA was initially denatured at 95°C for 5 min, followed by annealing at 52°C for 90 sec, elongation at 72°C for 90 sec, with 44 repeated cycles and the final extension time was at 72°C for 10 min.
The PCR product was analyzed on 3% agarose gel stained with ethidium bromide and the DNA bands were visualized using a UV camera by which the image was photographed, captured and analyzed using NIH image, National Institute of Health computer graphics.
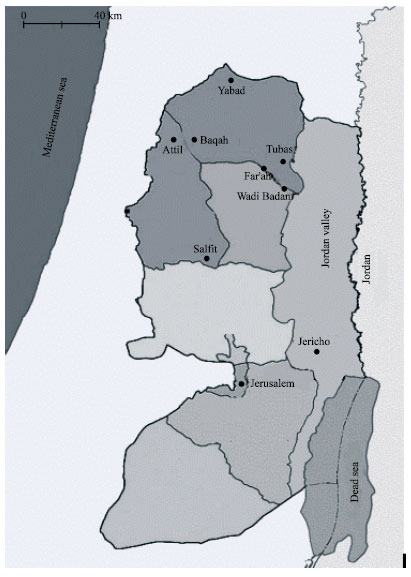 | |
| Fig. 1: | Gegraphical origins of colony and field collection of P. papatasi populations in the West Bank |
Data Collection and Analysis
Data were scored as presence or absence of PCR bands in each genotype. The band presence- absence matrix was constructed and imported into TreeCon program for Windows, version 1.3b (Van de Peer and De Wachter, 1994) which was used to calculate Nei-Li distances (Nei and Li, 1979) and to display the unweighted pair-group method (UPGMA) dendrogram.
RESULTS
PCR Profiles
The amplification conditions used were chosen to generate discrete amplification products of different sizes that could be used to differentiate between P. papatasi populations.
The banding pattern of male sand flies that came from colony J were different from those from colony A (Fig. 2, lanes 1 and 2) that originated from the same site. An interesting finding that the PCR profiles of female sand flies that originated from J colony (Fig. 2, lanes 3-6) were similar to the banding pattern of the male profile of A colony.
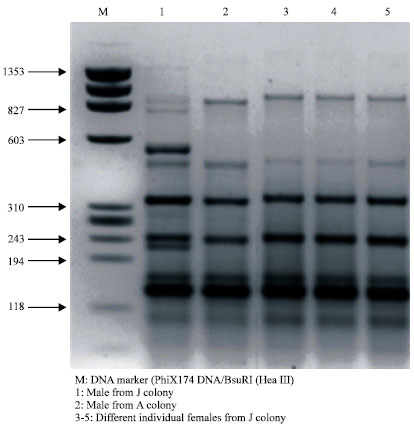 | |
| Fig. 2: | Characterization of different male and female P. papatasi sand flies from J and A colonies |
On the other hand females or males P. papatasi that came from A colony generated similar PCR profiles, the overall amplification patterns expressed in the number of bands and the molecular weight sizes were identical (data not shown).
The reproducibility of our system was further tested with other P. papatasi individuals originating from different sites in the West Bank. Significant variations were found among these populations both in number and sizes of the PCR bands and the variation was evident except for those from Wadi Badan and Yabad (Fig. 3, lanes 1-2). The variation in the flies from these two villages was manifested by the lack of the 603 bp band in Yabad population. However, most of the populations studied showed conserved bands that were common in many populations (band size 234 bp, Fig. 3). Reactions were also examined using each primer individually. No products were obtained when either AU 489 or AL997 were used (data not shown).
Phylogenetic Analysis
The phylogenetic tree generated by UPGMA method showed the existence of two distinct groups of P. papatasi in the studied areas. The dendrogram pointed to a genetic clumping that is depicted geographically in Fig. 1 and graphically in Fig. 4. The first group represent all individuals derived from the Jordan Valley except one derived from Tubas, two sub-clusters were identified in this group; Tubas and Jericho joined together forming minor branch to which the colony derived individuals (J and A) were connected to form another minor branch. It was surprising that the Jericho field individual, which came from the Jordan Valley was distant from the neighboring J and A colony individuals and found to be close to Tubas individual with genetic distance of 0.2.
The other group represent the other individuals derived from the Northern areas of the West Bank; Yabad, Wadi Badan and Attil. In this group, Yabad and Wadi Badan individuals were grouped together with genetic distance less than 0.2, this actually pointed to the fact that these two populations are genetically very close.
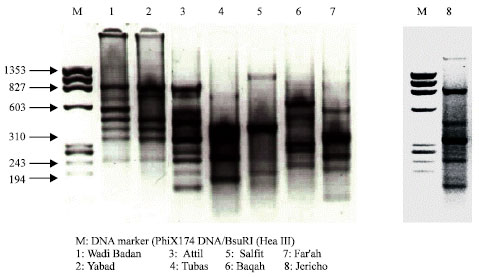 | |
| Fig. 3: | PCR profiles of eight P. papatasi individual sand flies collected from different geographical areas in the West Bank as indicated. Ten microliter of DNA from individual sand flies from different locations was analyzed |
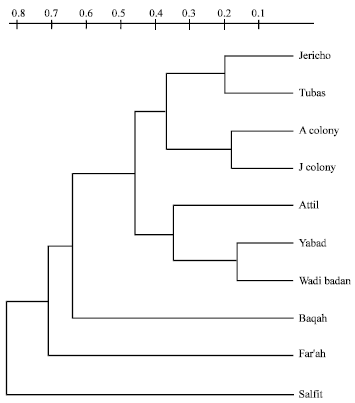 | |
| Fig. 4: | Unweighted pair-group method dendrogram of P. papatasi sand flies collected from different geographical areas in the West Bank |
On the other hand, the genetic distance between other field individuals (Baqah, Fara'h and Salfit) showed high level of divergence and lack prominent clustering.
DISCUSSION
About 122 species of Phlebotomine sand flies have been identified in the Old World and among of these, the role of 50 species has been established as vectors of various diseases, particularly leishmaniasis (Killick-Kendrick, 1990; Lane, 1993). Vector competence and vectorial capacity of many species still remain to be investigated (Wu and Tesh, 1990; Hanafi et al., 1998; Aransay et al., 2003). The vector parasite-relationship in leishmaniasis has not been unraveled in several cases because of the lack of molecular markers to distinguish between the closely related sand flies (Mukhopadhyay et al., 2000; Hamarsheh et al., 2007).
In Drosophila, the α-amylase is a multicopy gene of homologous chromosome regions carrying the standard gene arrangement contain the complete coding sequence. Recently, multigene families gained considerable attention, since it has been argued that reconstructing phylogeniesof multigene families is central to understanding the evolutionary meaning of divergence of genes as a result of physiological adaptation (Araki et al., 2005).
In this study we present a PCR system that uses primers to amplify α-amylase gene sequences to distinguish between different P. papatasi populations. The PCR profiles yielding five to eight products were obtained and easily visualized by agarose gel electrophoresis. The banding patterns were unique for each population and much simpler to analyze than the isoenzyme electrophoresis.
The minor intra-specific differences in the PCR profiles observed between J and A colonies (Fig. 2, lanes 1 and 2) that were originated from the same site is related to the fact that the sand fly individuals in J colony was inbred in which both male and female P. papatasi sand flies from other populations were introduced to the colony in early generations (personal communication with Dr. R. Jacobson, the founder of the colony). These minor variations in the banding pattern between A and J seemed to be restricted to the males of the J colony rather than to the females of the same colony. This conclusion was drawn when high similarity was generated between the banding patterns of different females from J colony and the banding pattern of a male from A colony. Although there is no molecular evidence that could prove this conclusion in sand flies, Steinemann and Steinemann (1999) provide an explanation for the presence of an amylase gene cluster on the evolving sex chromosomes of Drosophila miranda.
On the other hand the presence of minor differences in the PCR profiles between A and J colonies indicate the specificity of the primers used to detect these minor differences even after several generations of colonization.
Present study confirms the presence of genetic differences among different local populations of P. papatasi, which has been never reported before for this widely distributed species. The variation between the Jordan Valley colonies (A and J) and the neighboring Jericho population is probably due to local structuring of P. papatasi populations in this area. This result is in congruence with studies carried on L. major; the Leishmania species usually transmitted by P. papatasi vector, where two genetically isolated groups were found to co-exist in the same territory in the Middle East as shown by different genetic markers (Elfari et al., 2005). Tubas was grouped with Jericho individual rather than with the other Northern West Bank individuals, this probably related to the fact that Tubas area is located on an edge close to the Jordan Valley which is considered as wide low territory extending from the North to the South, it is almost dry area but vegetation increased by moving from South to the North.
P. papatasi sand flies that were collected from Yabad and Wadi Badan grouped in the UPGMA tree in a paraphyletic arrangement despite the large geographical distance between the two localities. This might reflect that these populations are very closely related. Migration of sand flies between these areas was not likely since sand flies were unable to migrate for longer distances, conversely, human aided transportation of sand flies or even sand fly larvae were not excluded in high agricultural activity areas. On the other hand, Baqah, Fara’h and Salfit individuals failed to group with other individuals in the tree and therefore did not show any significant structuring. This might be that these individuals were genetically variable and not significantly characterized at this level using this marker. Other genetic markers for P. papatasi may be needed to characterize individuals of these populations.
The high level of variation among P. papatasi populations manifested by different PCR profiles is consistent with studies carried by Hamarsheh et al. (2006), Kassem et al. (1993) and Ghosh et al. (1999), in which microsatellite markers and isoenzyme electrophoresis were used to detect polymorphisms in a group of P. papatasi sand flies derived from different populations in Egypt and Palestine respectively, these studies showed high rates of variation between Egyptian and Palestinian populations.
These data and the results of this study are in contrast to studies carried by Esseghir et al. (1997), who sequenced a mitochondrial DNA fragment (cytochrome b). They scored low levels of genetic variation among P. papatasi populations that came from different countries. This contrast probably related to the fact that mitochondrial DNA usually has maternal and nonrecombining mode of inheritance. The PCR system used here is characterized by high reproducibility among PCR assays made with the same individual and among individual members of the same population.
PCR-based methods are more robust than methods reliant on functional proteins and conveniently, specimens for PCR analysis can be preserved in alcohol, dried or frozen at -20°C. Methods that depend on assaying and measuring migration of isoenzymes require unbroken cold chains to prevent the degradation of enzymes.
Therefore, this PCR system may provide an accurate and simple method for detecting genetic variation among certain P. papatasi populations; it may provide more information on population structures. It is necessary to point out that a more extensive study using samples of wider geographical areas and evaluating the capacity of P. papatasi samples from each population to enhance Leishmania infection, is necessary to get more information on the structure of P. papatasi populations in Palestine.
ACKNOWLEDGMENTS
The authors thank Jose M.C. Riberio for providing original sequences of the α-amylase gene. This research was supported by Deutsch Forschungs-Gemeinschaft (DFG). Thanks also extend to Deutscher Akademischer Austausch Dienst (DAAD) for providing a Ph.D scholarship to Mr. O. Hamarsheh.
REFERENCES
- Adamson, R.E., R. Ward, M. Feliciangeli and R. Maingon, 1993. The application of random amplified polymorphic DNA for sand fly species identification. Med. Vet. Entomol., 7: 203-207.
Direct Link - Araki, H., S. Yoshizumi, N. Inomata and T. Yamazaki, 2005. Genetic coadaptation of the amylase gene system in Drosophila melanogaster: Evidence for the selective advantage of the lowest AMY activity and of its epistatic genetic background. J. Hered., 96: 388-395.
Direct Link - Aransay, A.M., P. Ready and F. Morillas-Marquez, 2003. Population differentiation of Phlebotomus perniciosus in Spain following postglacial dispersal. Heredity, 90: 316-325.
Direct Link - Boom, R., C.J. Sol, M.M. Salimans, C.L. Jansen, P.M.E. Wertheim-van Dillen and J. van der Noordaa, 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol., 28: 495-503.
PubMedDirect Link - Cameron, M.M., P. Milligan, A. Llanos-Cuentas and C. Davis, 1995. An association between Phlebotomine sand flies and aphids in the Peruvian Andes. Med. Vet. Entomol., 9: 127-132.
Direct Link - Cavalcante, R.R., M. Pereira, J. Freitas and F. Gontijo Nde, 2006. Ingestion of saliva during carbohydrate feeding by Lutzomyia longipalpis (Diptera: Psychodidae). Mem. Inst. Oswaldo Cruz., 101: 85-87.
Direct Link - Da-Lage, J., C. Maisonhaute, F. Maczkowiak and M. Cariou, 2003. A nested alpha-amylase gene in Drosophila ananassae. J. Mol. Evol., 57: 355-362.
Direct Link - Esseghir, S., P.D. Ready, R. Killick-Kendrick and R. Ben-Ismail, 1997. Mitochondrial haplotypes and phylogeography of Phlebotomus vectors of Leishmania major. Insect Mol. Biol., 6: 211-225.
CrossRefPubMedDirect Link - Elfari, M., L. Schnur, M. Strelkova, L. Eisenberger and R. Jacobson et al., 2005. Genetic and biological diversity among populations of Leishmania major from Central Asia, the Middle East and Africa. Microbes Infect., 7: 93-103.
Direct Link - Fryauff, D. and H. Hanafi, 1991. Demonstration of hybridization between Phlebotomus papatasi (Scopoli) and Phlebotomus bergeroti Parrot. Parasitologia, 33: 237-243.
Direct Link - Ghosh, K.N., J. Mukhopadhyay, H. Guzman, R. Tesh and L. Munstermann, 1999. Interspecific hybridization and genetic variability of Phlebotomus sand flies. Med. Vet. Entomol., 13: 78-88.
Direct Link - Grossman, G.L., Y. Campos, D. Severson and A. James, 1997. Evidence for two distinct members of the amylase gene family in the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol., 27: 769-781.
Direct Link - Guernaoui, S., A. Boumezzough, B. Pesson and G. Pichon, 2005. Entomological investigations in Chichaoua: An emerging epidemic focus of cutaneous leishmaniasis in Morocco. Med. Vet. Entomol., 42: 697-701.
CrossRefPubMedDirect Link - Hamarsheh, O., W. Presber, Z. Abdeen, S. Sawalha, A. Al-Lahem and G. Schoenian, 2006. Isolation and characterization of microsatellite loci in the sand fly Phlebotomus papatasi (Diptera: Psychodidae). Mol. Ecol. Notes, 6: 826-828.
CrossRefDirect Link - Hamarsheh, O., W. Presber, Z. Abdeen, S. Sawalha, A. Al-Lahem and G. Schonian, 2007. Sequence analysis of the ribosomal DNA ITS2 region for Phlebotomus papatasi (Diptera: Psychodidae). J. Entomol., 4: 167-180.
CrossRefDirect Link - Hanafi, H.A., B. El-Sawaf, D. Fryauff, G. Beavers and G. Tetreault, 1998. Susceptibility to Leishmania major of different populations of Phlebotomus papatasi (Diptera: Psychodidae) from endemic and non-endemic regions of Egypt. Ann. Trop. Med. Parasitol., 92: 57-64.
Direct Link - Jacobson, R.L. and Y. Schlein, 2001. Phlebotomus papatasi and Leishmania major parasites express α-amylase and B-glucosidase. Acta Trop., 78: 41-49.
Direct Link - Jacobson, R.L., Y. Schlein and C. Eisenberger, 2001. The biological function of sand fly and Leishmania glycosidases. Med. Microbiol. Immunol., 190: 51-55.
Direct Link - Jaffe, C.L., G. Baneth, Z. Abdeen, Y. Schlein and A. Warburg, 2004. Leishmaniasis in Israel and the Palestinian authority. Trends Parasitol., 20: 328-332.
CrossRefPubMedDirect Link - Kassem, H.A., D. Fryauf, B. El Sawaf, M. Shehata and N. Shoumar, 1990. Electrophoretic comparison of the Leishmania vectors Phlebotomus papatasi and Phlebotomus langeroni (Diptera: Psychodidae). J. Med. Entomol., 27: 592-601.
Direct Link - Kassem, H.A., D. Frayauf, M. Shehata and B. El-Sawaf, 1993. Enzyme polymorphism and genetic variability of one colonized and several field populations of Phlebotomus papatasi (Diptera: Psychodidae). J. Med. Entomol., 30: 407-413.
Direct Link - Kassem, H.A., D. Fryauff and H. Hanafi, 1999. Enzyme polymorphism and genetic variability of Laboratory populations of Phlebotomus papatasi, P. bergeroti, P. langeroni and P. perniciosus. J. Egypt. Soc. Parasit., 29: 459-472.
Direct Link - Killick-Kendrick, R., 1990. Phlebotomine vectors of the leishmaniases: A review. Med. Vet. Entomol., 4: 1-24.
CrossRefDirect Link - Kellick-Kendrick, R., 1999. The biology and control of Phlebotomine sand flies. Clinics in Dermatology, 17: 279-289.
CrossRef - Lanzaro, G.C. and A. Warburg, 1995. Genetic variability in Phlebotomine sand flies: Possible implications for leishmaniasis epidemiology. Parasitol. Today, 11: 151-154.
Direct Link - Maczkowiak, F. and J.L. Da-Lage, 2006. Origin and evolution of the Amyrel gene in the alpha-amylase multigene family of Diptera. Genetica, 128: 145-158.
Direct Link - Mukhopadhyay, J., K. Ghosh and H. Braig, 2000. Identification of cutaneous leishmaniasis vectors, Phlebotomus papatasi and P. duboscqi using random amplified polymorphic DNA. Acta Trop., 76: 277-283.
Direct Link - Muller, G. and Y. Schlein, 2004. Nectar and honeydew feeding of Phlebotomus papatasi in a focus of Leishmania major in Neot-Hakikar oasis. J. Vector Ecol., 29: 154-158.
Direct Link - Muller, G. and Y. Schlein, 2005. Plant tissues: The frugal diet of mosquitoes in adverse conditions. Med. Vet. Entomol., 19: 413-422.
Direct Link - Nasereddin, A. and C. Jaffe, 2004. Effect of the security fence on zoonotic visceral leishmaniasis in Israel and Palestine. Lancet Infect. Dis., 4: 393-394.
Direct Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Parvizi, P., M. Benlarbi and P. Ready, 2003. Mitochondrial and Wolbachia markers for the sand fly Phlebotomus papatasi: Little population differentiation between peridomestic sites and gerbil burrows in Isfahan province, Iran. Med. Vet. Entomol., 17: 351-362.
Direct Link - Parvizi, P. and P.D. Ready, 2006. Molecular investigation of the population differentiation of Phlebotomus papatasi, important vector of Leishmania major, in different habitats and regions of Iran. Iran. Biomed. J., 10: 69-77.
Direct Link - Perrotey, S., S.S. Mahamdallie, B. Pesson, K.J. Richardson, M. Gallego and P.D. Ready, 2005. Postglacial dispersal of Phlebotomus perniciosus into France. Parasite, 12: 283-291.
Direct Link - Ribeiro, J.M., E. Rowton and R. Charlab, 2000. Salivary amylase activity of the phlebotomine sand fly, Lutzomyia longipalpis. Insect. Biochem. Mol. Biol., 30: 271-277.
Direct Link - Schlein, Y., A. Gunders and A. Warburg, 1982. Leishmaniasis in the Jordan Valley, I Attraction of Phlebotomus papatasi (Psychodidae) to turkeys. Ann. Trop. Med. Parasitol., 76: 517-520.
CrossRefDirect Link - Schlein, Y. and R. Jacobson, 1999. Sugar meals and longevity of the sandfly Phlebotomus papatasi in an arid focus of Leishmania major in the Jordan Valley. Med. Vet. Entomol., 13: 65-71.
Direct Link - Schlein, Y. and R. Jacobson, 2000. The rate of photosynthesis modulates the plant feeding of the sand fly Phlebotomus papatasi (Diptera: Psychodidae). J. Med. Entomol., 37: 319-324.
Direct Link - Steinemann, S. and M. Steinemann, 1999. The amylase gene cluster on the evolving sex chromosomes of Drosophila miranda. Genetics, 151: 151-161.
Direct Link - Van de Peer, Y. and R. de Wachter, 1994. TREECON for Windows: A software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput. Applied Biol. Sci., 10: 569-570.
CrossRefDirect Link - Ward, R., N. Pasteur and J.A. Rioux, 1981. Electrophoretic studies on genetic polymorphism and differentiation of phlebotomine sandflies (Diptera: Psychodidae) from France and Tunisia. Ann. Trop. Med. Parasitol., 75: 235-245.
CrossRefDirect Link - Wu, W.K. and R. Tesh, 1990. Genetic factors controlling susceptibility to Leishmania major infection in the sand fly Phlebotomus papatasi (Diptera: Psychodidae). Am. J. Trop. Med. Hyg., 42: 329-334.
Direct Link