
J.N. Malinga
KAR1-Njoro, P.O. Private Bag, 20107, Njoro, Kenya
M.G. Kinyua
KAR1-Njoro, P.O. Private Bag, 20107, Njoro, Kenya
A.W. Kamau
Ministry of Agriculture, P.O. Box 30028, 00200, Nairobi
J.K. Wanjama
Department of Crops and Soil Science, Egerton University,
P.O. Box 536, 20107, Njoro, Kenya
J.O. Awalla
KAR1-Njoro, P.O. Private Bag, 20107, Njoro, Kenya
R.S. Pathak
Department of Crops and Soil Science, Egerton University,
P.O. Box 536, 20107, Njoro, Kenya
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 5 | Page No.: 350-361







ABSTRACT
Recent development of virulent (Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae) (RWA) biotypes is a challenge to insect resistance breeding programs. The aim of this study was to detect the existence of biotypes within two Russian wheat aphid populations sampled from Njoro and Timau locations of Kenya. Ten clones were raised from each population and used in population increase studies, virulence tests on different wheat cultivars and Amplified Fragment Length Polymorphic (AFLP) DNA fingerprinting assays. Results of two population increase tests indicated that aphid survivorship was high on Njoro biotype with mortality being concentrated at the end of the maximum life span. Timau biotype had low survivorship with mortality concentrated early in the cycle. Timau biotype had fewer progeny and lower estimates of intrinsic rate of natural increase on susceptible bread-wheats. Results of virulence tests showed that no clear virulence trend was recorded on the biotypes but Njoro biotype was appreciably more virulent on some resistant wheats than Timau biotype. One AFLP primer pair, E-AAC/M-CAC, gave amplified and polymorphic DNA fingerprints and more than one RWA genotype were identified among clones of Njoro and Timau biotypes. The study shows the existence of more than one biotype and/or genotype present within tropical RWA populations.
PDF Abstract XML References Citation
How to cite this article
J.N. Malinga, M.G. Kinyua, A.W. Kamau, J.K. Wanjama, J.O. Awalla and R.S. Pathak, 2007. Biotypic and Genetic Variation Within Tropical Populations of Russian Wheat Aphid, Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae) in Kenya. Journal of Entomology, 4: 350-361.
DOI: 10.3923/je.2007.350.361
URL: https://scialert.net/abstract/?doi=je.2007.350.361
DOI: 10.3923/je.2007.350.361
URL: https://scialert.net/abstract/?doi=je.2007.350.361
INTRODUCTION
Russian wheat aphid (Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae: Macrosiphini) has become the major pest of wheat and barley in Kenya. Economic importance of Russian wheat aphid also known as RWA is attributed to reduced grain, loss of kernel weight, quality and increased costs of production due to application of insecticides (Marasas et al., 1997). Even with the use of contact insecticides the yield losses attributed to Russian wheat aphids in farmers’ fields in Kenya may reach 40% (Kinyua et al., 2003). Systemic aphicides are effective but costly and lead to loss of environmental purity and quality. The aphid attacks the plant by infesting the young growing tip deep in the leaf whorl where it feeds from the phloem of longitudinal veins. Symptoms of its attack appear as chlorotic spots that coalesce to form white, yellow or purple streaks running parallel to the midrib of leaves (Botha, 2004). In young plants, heavy infestation leads to prostrate tillers while adult plants show trapped ears within the flag leaf looking like a fishhook. Severe infestations may lead to head sterility and death of the host plant.
Development of resistant varieties appears to be the most sustainable approach in the management of RWA. However, a successful insect resistance breeding program requires an understanding of insect biotypes occurring in the locality. Biotypes may overcome resistant plants and failure to recognize their existence may lead to severe infestations of formerly resistant cultivars. Biotypes can be detected using several approaches and among these are phenotypic and molecular markers. Phenotypic markers in aphids have commonly used population increase tests that measure (survival, growth, reproduction) and virulence tests that measures (cultivars reaction to insect attack). Workers postulate that differences in population increase parameters or in reaction of the lines to the insect isolates constitute differences in virulence and indicate biotypes (Haley et al., 2004; Basky, 2003). Puterka et al. (1992) used both population increase and virulence tests. The workers postulated that units identified as biotypes may be different clones, populations or phenotypically similar groups composed of unknown genotypes. Other workers have employed molecular techniques to characterize RWA populations and indicated that band differences at DNA level show biotypic and/or genotypic variation but could not link genotypic variation to biotypic variation (Robinson et al., 1993; Black et al., 1992). Castro et al. (2005) demonstrated that differences in antixenosis tests were linked to DNA markers and could be used to identify RWA biotypes. Biotypes would not be a concern in the tropics as RWA populations occurring in mild climates until recently were thought not to consist of biotypes. The RWA populations are said to reproduce predominantly by parthenogenesis with females giving birth to live young from unfertilized eggs. However, recent reports suggest that RWA biotypes do occur under mild climates. Haley et al. (2004) demonstrated differential virulence patterns between originally identified RWA and a new isolate within mild climates of the United States of America (USA). To date there are very limited reports on biotypic variation in tropical D. noxia populations although differences in varietal performance between USA and Kenya D. noxia populations (Kiplagat, 2005) and differences in DNA fingerprints between USA and Ethiopia (Smith et al., 2004) have been noted. The objective of this study was to detect biotypic variation within tropical RWA populations using population increase, virulence tests and AFLP-PCR markers with a view to inform on RWA biotypes in Kenya. AFLP-PCR markers were selected as they have higher reproducibility, apply stringent PCR conditions and detect even single nucleotide polymorphism (Vos et al., 1995).
MATERIALS AND METHODS
Sampling and Rearing Aphid Clones
Two aphid colonies were collected from symptomatic bread-wheat at two different locations in Kenya; Njoro, Kenya (0 20' S; 35° 56' E; 2166 m above sea level (asl) and Timau, Kenya (0 05' S; 37° 20' E; 2640 m (asl). The aphids were taken to the Kenya Agricultural Research Institute, Njoro; where the experiments were conducted. Njoro is located in the lower highlands (LH3) with a mean annual rainfall of 931 mm and temperatures of 7.9 to 21.9°C and a mean of 14.9°C (Jaetzold and Schmidt, 1983). The aphids were identified with the aid of a 10X magnifier according to descriptions given by Hein et al. (1989). Viviparous adult female RWA aphids were separately settled on clean barley seedlings, in pots within plexi-cages (50x55x48 cm) in the greenhouse at Njoro. The barley seedlings had been grown under screen cages in the greenhouse to keep them clean with temperatures that fluctuated between 10 and 31°C (Night: Day) under natural light conditions. The plants were watered regularly to ensure that they did not suffer moisture stress. The two aphid colonies were denoted Njoro and Timau biotype consistent with the location where they were collected. Inoculation was done by gently placing a single aphid using a paintbrush within the leaf whorl and allowing it to multiply to form a clone. In all, 10 different clones of each biotype were set up and were used in virulence and population increase tests on one wheat cultivar Kenya Mbuni. To ensure that no crossovers occurred between the two biotypes, we collected fresh RWA colonies from the same locations. The new clones were used in a broader population increase test by assaying half of the clones on five bread-wheat entries and the other half, using DNA fingerprinting assays.
Production and Maintenance of Test Plants
Throughout the study period a continuous supply of clean (aphid free) wheat seedlings for the experimental work was maintained and leaf excised from these for the experiment. Ten 0.75 L pots per entry each containing five plants were planted in sterilized soil/manure mixture at the ratio of 3:1 and containing 5 g Di-ammonium phosphate fertilizer and 2 g copper oxychloride every three days to provide a fresh supply. The pots were caged at planting with screen cloth supported on a wire frame. When leaves attained five to six leaf stages, the largest leaves were excised and used for leaf sections.
Rearing Same Age Females for the Study
To obtain females of the same age for the experiment, a single 3rd to 4th instar aphid was taken from the clones, settled on a 6 cm long leaf section, placed in moist sand for 24 h and allowed to larviposit nymphs. After 24 h, one nymph was selected while the rest (adult and the nymphs) were killed. The retained nymphs on the leaf sections were approximately the same age with 24 h as maximum difference. A single aphid of known age was then settled on an excised leaf of 6 cm length and cut 6 cm long up from the base of the sheath and inserted in moist sand in plastic petri-dishes. The petri-dishes were placed in controlled environment chambers in a complete randomized design using 24 clones for Njoro biotypes and 45 clones for Timau biotypes, respectively. Each biotype was put in a separate conviron maintained at temperatures of 25:18°C photoperiod 12:12 [L: D] and relative humidity varying between 60-80%. The aphids of known age were then left to larviposit and nymphs born to them counted and removed daily until death occurred. The leaf section was changed every two to three days while sand was moistened daily.
The experiment was repeated on five more different wheat entries (one moderately resistant line (KRWA-9) and Doubled haploid (DH) progeny (DH NJBW1/KRWA-9 and NJBW1/KRWA-9) and two susceptible lines Kenya Kwale and Kenya Pasa). Estimates of intrinsic rate of natural increase (rm) were calculated using the method of Wyatt and White (1977) as shown;
where, the development time (t) in days for D. noxia from birth to onset of reproduction, the number of progeny (Mt) subsequently produced by each aphid in a span equivalent to its development time (t) were recorded. Cohort generation time (Tc) was calculate as Tc = 4t/3. Data was analyzed on both surviving cohort attaining reproduction and total cohort, using SAS statistical package and the means transformed using √(x+1) to standardize the variance.
Phenotypic Characterization of Aphid Biotypes Using Virulence Tests
Thirty-six wheat lines Kenya Heroe, Kenya Duma, Kenya Kwale, Kenya Chozi, Kenya Pasa, Kenya Fahari, KRWA-4, KRWA-8, KRWA-9, KRWA-16 and their F1 crosses were planted in the greenhouse on evaluation flats (95x23.5x7.5 cm). Their pedigree is given in Table 1. The test lines were planted in sterile soil/manure mixture (3:1) that was added with 2 g copper oxychloride and 5 g Diammonuim phosphate (18-46-0). Two seeds of each of the test plants were planted in 10x3.725 cm plots and caged with a wire cage 60 cm high and polyester screen mesh (68 meshes per square cm). Eight days after emergence, test plants were infested with two apterous aphids by placing them at the base of the plant and allowing them to multiply undisturbed for 21 days. Aphids were taken from a mixture of clones from Njoro and Timau biotypes, respectively. Twenty one days after infestation, damage on infested test plants was assessed on a modified scale of (1-9) for resistance against RWA (1,2,3 resistant, 4,5-moderately resistant, 6 moderately susceptible, 7,8,9-susceptible (modified scale of Tolmay, 1995). Data on damage rating scores was transformed to standardize variance then analysis of variance was carried out on it using SAS package.
| Table 1: | Nomenclature and sequences of AFLP primers used in the study according to Keygene http://www.keygene.nl/htll/nomenclature.htm |
 | |
DNA Extraction from Individual RWA Aphids
The remaining clones used in the previous study had 24 individual aphids isolated per biotype. The aphids were examined carefully for predators or parasites and genomic DNA extracted from them. Fresh aphid tissue plus sand and proteinase K was ground in liquid nitrogen. The mixture was extracted using a protocol based on Promega wizards genomic DNA purification and extraction mini-kit and following the given manufacturers instructions (Wizard Plus mini-preps (DNA purification systems CAT # A7500; www.promega.com). The DNA concentration per sample was quantified using a spectrophotometer and resolved on a 2% agarose gel through electrophoresis to determine the quality. Presence of the DNA was confirmed by visualizing the bands on the gel on the trans-illuminator.
AFLP-PCR Protocol
The extracted DNA was stored on dry ice for DNA quantification and PCR analysis. The DNA was adjusted accordingly to give a concentration of 10 ηg. The DNA was first restricted to completion with EcoRI and MseI enzymes in a total reaction volume of 25 μL at 37°C for 2 h. The restriction enzymes were heat inactivated at 70°C for 15 min. The restricted DNA samples were then ligated to adapter ligation solution containing EcoRI and MseI adapters using T4 DNA ligase in a reaction volume of 50 μL at 20°C for 2 h. The resultant DNA template was amplified with EcoRI (end labelled with [P33] dATP) and MseI selective primers with one pair of nucleotides at the 3 primer end complementary to the adapters ;) E-AAC/M-CAC.
The product was mixed with AFLP forward and reverse primers and PCR was carried out according to basic PCR protocol. PCR cycle conditions included 94°C for 2 min followed by 30 cycles repeat of denaturation at 94°C for 1 min, annealing at 50°C for 1 min and extension at 72°C for 60 sec followed by 30 cycles (94°C for 30 sec, 50°C for 30 sec, 72°C for 1 min) and a final extension of 72°C for 10 min. For each sample, 5 μL of reaction mix was run in 6% polyacrylamide gel and the reaction products visualized using radioactivity.
RESULTS
Population Increase of Russian Wheat Aphid Biotypes Raised on Wheat
Figure 1 shows data pertaining to survivorship of apterous RWA clones of Njoro and Timau biotypes raised on wheat in the growth chamber under controlled temperatures (18-25), photoperiod 12:12 h) n = 24 for Njoro Biotype (Type I curve), n = 45 for Timau biotype (Type III curve). The figure shows that Njoro biotypes experienced high survivorship with mortality being concentrated at the end of the maximum lifecycle. This gave a Type I response curve while Timau biotypes experienced low survivorship with mortality being concentrated early in the lifecycle. This gave a Type III response curve (Begon et al., 1990). Despite the high mortality observed on Timau biotypes the few survivors lived longer (45 days) than Njoro aphids (40 days) (Data not shown). However, Njoro biotypes had appreciably higher cohort generation time and subsequently higher estimates of intrinsic rate natural increase than Timau biotypes (Fig. 2). Percent survivorship, development time, progeny size and intrinsic rate of natural increase of Njoro and Timau biotypes raised on five wheat entries (susceptible K.Pasa and K.Kwale and resistant KRWA-9 and its progeny) is reported in Fig. 3.
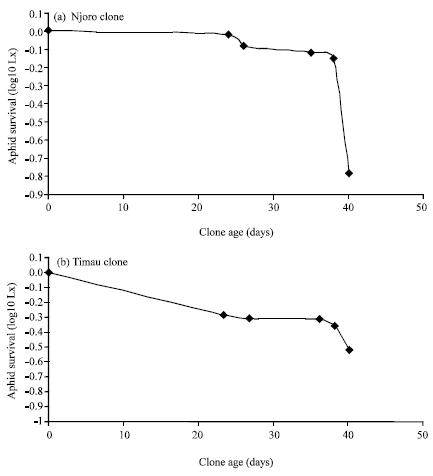 | |
| Fig. 1: | Survivorship of apterous RWA clones of Njoro and Timau biotypes raised on wheat in the growth chamber under controlled temperatures (18-25), photoperiod 12:12 h) n = 24 for Njoro biotype (Type I cure), n = 45 for Timau biotype (Type III curve) |
Figure 3 shows that Timau biotypes suffered significantly higher mortality than Njoro biotypes on all wheat cultivars including the susceptible ones. However, the study showed that the two biotypes did not differ in development times required to reach reproduction. The survivors of Timau biotype reproduced appreciably fewer progeny on all of the wheat entries except on Kenya Pasa than that produced by Njoro biotypes. Similarly, estimates of intrinsic rate of natural increase were lower for Timau biotypes than on Njoro biotypes. Wheat entries (Table 2) containing the unknown RWA gene in entry KRWA-9, recorded lower progeny size and estimates of intrinsic rate of natural increase than that recorded on susceptible wheat entries (Kenya Kwale and Kenya Pasa). The susceptible wheats (Kenya Kwale and Kenya Pasa) were significantly (p<0.05) more suitable as a host for colonization and reproduction than the resistant KRWA-9 and its progenies.
Virulence Tests
Data pertaining to means±SD of RWA damage (scale 1-9; 1-resistant; 9-susceptible) on selected wheat cultivars by Njoro and Timau RWA biotypes in the greenhouse at Njoro in 2004 are reported in Table 3. The two biotypes showed differential reaction on the wheat lines but the differences were not significant with no clear trend recorded. The susceptible wheats were damaged by infestation of both Njoro and Timau biotypes with degree of susceptibility dependant on the biotype used.
| Table 2: | Wheat entries used in this study, their origin and pedigree |
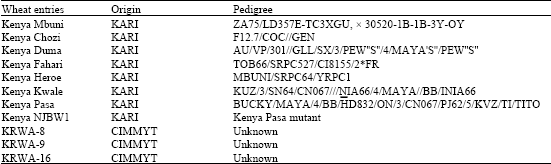 | |
| *KARI-Kenya Agricultural Research Institute | |
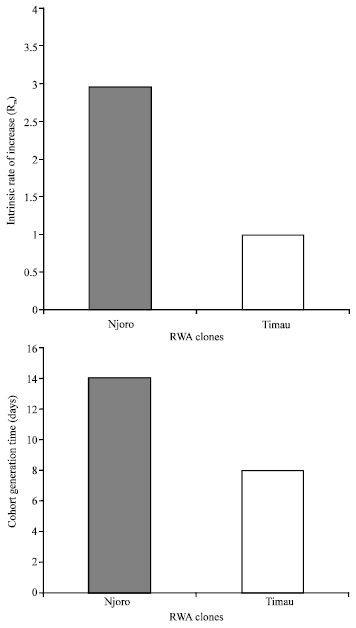 | |
| Fig. 2: | The intrinsic rate of natural increase and cohort generation time of RWA clones of Njoro and Timau biotypes raised on wheat under controlled temperatures (18-25°C), photoperiod 12:12 h) n = 24 for Njoro Clone, n = 45 for Timau clone |
| Table 3: | Means±SD of RWA damage (scale 1-9; 1-resistant; 9-susceptible) on wheat cultivars by Njoro and Timau RWA biotypes in the greenhouse at Njoro 2004 |
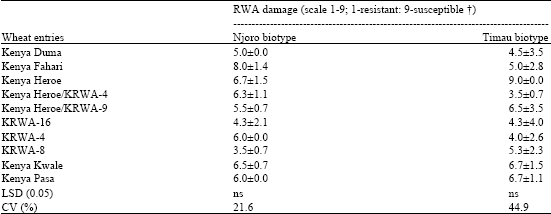 | |
| †RWA scale 1-3: resistant, 4-5; moderate resistance; 6-9 susceptible, ns: Non-significant | |
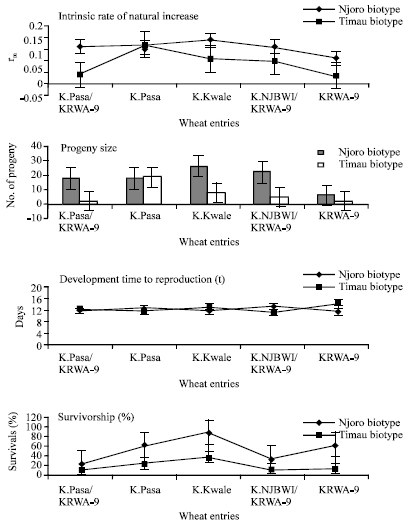 | |
| Fig. 3: | Percent survivorship, development time, progeny size and intrinsic rate of natural increase of Njoro and Timau biotypes raised on five wheat entries (susceptible K. Pasa and K. Kwale and resistant KRWA-9 and its progeny) |
| Table 4: | AFLP-PCR bands showing polymorphism in 12 RWA clones taken from of Njoro (N) and Timau (T) biotypes and representing bands amplified with E-AAC/M-CAC primer pair. The presence of a band is denoted (+) while absence of a common band is denoted (-) |
 | |
The resistant wheats identified by a prefix of KRWA-, had one of the entries KRWA-8, selected as resistant in preliminary screening tests (Malinga et al., 2001) being severely more damaged by Timau than Njoro biotype. KRWA-4 and its progeny (Kenya Heroe/KRWA-4) were more damaged by Njoro biotype while KRWA-16 remained unaffected maintaining moderate resistance to both biotypes.
AFLP-Map of Njoro and Timau Biotypes
Results of DNA profiles of individual aphid genomic DNA is presented in Fig. 4 and Table 4 and 5. Results from the AFLP analysis showed that the DNA was of high quality. The band pattern produced from the products of AFLP differed with the different primer pairs used. Not all the primer pairs were useful in distinguishing fragment profiles. In this study, the AFLP fingerprints derived from primer pair E-AAC/M-CAC produced the best band pattern that was distinct and showed polymorphic bands on randomly selected clones (Fig. 4). The E-AAC/M-CAA primer was monomorphic although a single faint polymorphic band was observed on one of the Njoro clones and which was not analyzed. The E-AAC/M-CAG primer combination produced the poorest band pattern with unclear bands and it was not possible to distinguish between fragment profiles so it was not analyzed. The AFLP fingerprints observed on primer pair E-AAC/M-CAC showed that there were at least two distinct types of clones (Fig. 4). Most of Njoro and Timau clones shared fingerprints and had few polymorphic bands.
| Table 5: | Percentage (%) polymorphic bands observed between selected aphid clones taken from Njoro and Timau biotypes using primer combination E-AAC/M-CAC |
 | |
| *N-Njoro biotype, T-Timau Biotype | |
 | |
| Fig. 4: | AFLP-PCR profile map of genomic DNA of single clones of RWA Njoro (N) and Timau (T) biotypes amplified and resolved on a 6% polyacrylamide gel assayed against E-AAC/M-CAC primers. Band (a) and (C) are examples of well amplified bands showing polymorphism. Band b are sporadic bands that are also well amplified and were scored |
However, a few clones had missing bands at a common banding site, denoted by band (a) and (c). Among such clones was N11 from Njoro biotype and this was considered to be of a different genotype from all other clones. A few clones had sporadic but well amplified bands as denoted by band (b) (Table 4). They were considered to belong to another genotype. At least two clones (N5 and N22) belonging to Njoro biotype showed a high percentage of polymorphism when compared to other clones of Njoro and Timau and were considered to belong to a different genotype (Table 5). The observed variation within (N5 and N22) belonging to Njoro biotype was larger than that observed between some of the clones of Njoro and Timau biotypes.
DISCUSSION
Differences in life history parameters result in different population growth rates, which are an important measure for differentiating insect biotypes (Basky, 2003). In the present study, it was shown that Njoro biotypes experienced higher survivorship, progeny and estimates of intrinsic rate of natural increase than Timau biotypes. This indicated that Njoro biotypes were able to colonize and reproduce on the wheat crop more successfully than the Timau biotypes indicting higher virulence or aggressiveness. The low survivorship and reproduction recorded on Timau biotype could result in slow buildup of RWA infestation and inability to attain economic thresholds before the wheat plant attains maturity. The differences observed may be attributed to locational niche environments that may arise from adaptation or mutation within the insect. The results would suggest that the aphids may experience speciation and ecological adaptation resulting in geographical parthenogenesis (Wilson et al., 2003). Despite the detected differences, the virulence tests did not confirm the hypothesis that Njoro biotypes were more virulent than Timau clones. Both biotypes had clones which were virulent on some wheat entries indicating the possible existence of more than one biotype within each of the Njoro and Timau population. The lack of a clear trend observed on the two biotypes may also have resulted from the use of a mixture of clones in each biotype, leading to sampling error with some the clones possibly being virulent. The differences could also be attributed to differences in host plant–aphid interactions. New virulence has been found to constitute a biotype when the new pest populations severely damages already identified resistant varieties (Haley et al., 2004). The present application of AFLP-PCR markers revealed polymorphic fingerprints observed on primer pair E-AAC/M-CAC. The observed variation within Njoro biotypes is larger than is observed between some of the clones of Njoro and Timau biotypes indicating that the biotypes are not reproductively isolated and gene flow probably occurred. The findings indicate that more than one genotype could be present within Njoro populations. This could belong to a rare or poorly fit genotype. The rare genotypes possibly accounted for the differences observed on fecundity, survivorship and intrinsic rate of natural increase between the two populations. The band differences observed in the present study reflect genetic variation within tropical aphid population occurring in Kenya. However, genetic variation is thought to be uncommon in tropical or mild climate environments as RWA undergo anholocyclic life cycles, reproduce by parthenogenesis and lack hybridity. Some workers argue that clonal variation is a common phenomenon allowing the build up of low frequency genotypes into new populations even under mild climates (Wilson et al., 2003). This may account for genotypes observed from the molecular markers. The present study validates the work of Black et al. (1992) who demonstrated that DNA polymorphisms may be present within RWA population occurring in different countries under mild climates. Similar polymorphism has been demonstrated on parthenogenetic Sitobion avenae (F.) and Rhopalosiphum padi (L.) (Martinez et al., 1992; Carvalho et al., 1991). Polymorphism has also been reported by different workers in RWA populations collected in the USA where the aphid has exclusive parthenogenesis mode of reproduction (Lapitan et al., 2006; Baker et al., 1996; Robinson et al., 1993). The genetic variation in the present study may have arisen from cyclic or obligate parthenogenesis, spontaneous mutability of quantitative traits, alary polymorphism, geographical parthenogenesis and induction through adaptation to abnormal environments, selection or other factors yet undetermined (Wilson et al., 2003; Papura et al., 2003; Black et al., 1992). Cyclic or obligate parthenogenesis is speculated as there is a possibility that sexual forms may occur along the snow peaked slopes of Mt Kenya located in an important wheat zone of Kenya. Parthenogenetic organisms often harbour substantial genotypic diversity to form mixed clones. This diversity may be the result of recurrent formations of new clones, or it may be maintained by environmental heterogeneity acting on ecological differences among clones (Vorburger, 2006). It has been shown that there are biotypic differences with regard to fecundity, survivorship, reproduction and intrinsic rate of natural increase between Njoro and Timau biotypes. There are also band differences at DNA level between and within the clones of the two RWA biotypes. The study has not been able to demonstrate a clear relationship between the phenotypic and molecular markers. Genotyping RWA populations by utilizing more colonies from different locations and using both phenotypic and more AFLP markers and sequencing unique bands will contribute to better characterization of Russian wheat aphid populations. The present study shows that RWA breeding programs in the tropics should undertake biotypic variation studies before embarking on crop improvement in development of resistant wheat.
ACKNOWLEDGMENT
The authors thank Egerton University, Kenya, the World Bank through Kenya Agricultural Research Institute (KARI) and International Atomic Energy Agency (IAEA) for facilitating the study.
REFERENCES
- Basky, Z., 2003. Biotypic and pest status differences between Hungarian and South African populations of Russian wheat aphid, Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae). Pest Mgt. Sci., 59: 1152-1158.
Direct Link - Black, W.C. IV., N.M. DuTeau, G.J. Puterka, J.R. Nechols and J.M. Pettorini, 1992. Use of the random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) to detect DNA polymorphisms in aphids (Homoptera: Aphididae). Bull. Entomol. Res., 82: 151-159.
Direct Link - Botha, C.E.J. and B. Matsiliza, 2004. Reduction in transport in wheat (Triticum aestivum) is caused by sustained phloem feeding by the Russian wheat aphid (Diuraphis noxia). South Afr. J. Bot., 70: 249-254.
Direct Link - Castro, A.M., A. Vasicek, M. Manifesto, D.O. Gimenez and M.S. Tacaliti et al., 2005. Dobrovolkaya, M.S. Roder, J.W. Snape and A. Boner, 2005. Mapping antixenosis genes on chromosome of wheat to greenbug and to a new biotype of Russian wheat aphid. Plant Breed, 124: 229-233.
Direct Link - Papura, D., J.C. Simon, F. Halkett, F. Delmotte, J.F. Le-Gallic and C.A. Dedryver, 2003. Predominance of sexual reproduction in Romanian populations of the aphid Sitobion avenae inferred from phenotypic and genetic structure. Heredity, 90: 397-404.
Direct Link - Smith, C.M., T. Belay, C. Stauffer, P. Stary, I. Kubeckova and S. Starkey, 2004. Identification of Russian wheat aphid (Homoptera: Aphididae) populations virulent to the Dn4 resistance gene. J. Econ. Entom., 97: 1112-1117.
Direct Link - Vos, P., R. Hogers, M. Bleeker, M. Reijans and T. van de Lee et al., 1995. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res., 23: 4407-4414.
CrossRefPubMedDirect Link - Vorburger, C., 2006. Temporal dynamics of genotypic diversity reveal strong clonal selection in the aphid Myzus persicae. J. Evol. Biol., 19: 97-107.
Direct Link - Wilson, A.C.C., P. Sunnucks and D.F. Hales, 2003. Heritable genetic variation and potential for adaptive evolution in asexual aphids (aphidoidea). Biol. J. Linnean Soc., 79: 115-135.
Direct Link - Wyatt, I.J. and P.F. White, 1977. Simple estimation of intrinsic increase rates for aphids and tetranychid mites. J. Applied Entomol., 14: 757-766.
Direct Link