
A.D. Banjo
Department of Plant Science and Applied Zoology, Olabisi Onabanjo University, Ago Iwoye, P.M.B. 2002, Ogun State, Nigeria
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 4 | Page No.: 308-316







ABSTRACT
Some aspects of the bioecology and behaviour of Gasteroclisus rhomboidalis Boh. at the Olabisi Onabanjo University were studied from March to September 2003. A survey of the speciation of the curculionid pest on Amaranthus plot of the University was carried out. The result revealed that three species occurred at study sites. The relative abundance of the pest, Gasteroclisus rhomboidalis earlier thought as a major pest of Amaranthus and Celosia is infact a minor one and that it does not occur abundantly. Infestation was all the year round on wild Amaranthus (Amaranthus spinosus) at the University site and possibly Amaranthus is the primary alternate host of this pest. The life history of G. rhomboidalis was studied using A. cruentus and result showed that there are three instar larval stages and take between 32 to 42 days (
PDF Abstract XML References Citation
How to cite this article
A.D. Banjo, 2007. Bio-Ecology and Life History of Gasteroclisus rhomboidalis Boh.(Coleoptera: Curculionidae) a Pest of Amaranthus cruentus (L.) Sauer. Journal of Entomology, 4: 308-316.
DOI: 10.3923/je.2007.308.316
URL: https://scialert.net/abstract/?doi=je.2007.308.316
DOI: 10.3923/je.2007.308.316
URL: https://scialert.net/abstract/?doi=je.2007.308.316
INTRODUCTION
Vegetables play a very important role in the human diet. They are important in neutralising the acid substance produced in the course of digestion of meat, cheese and other foods and are of value as roughages which promote digestion and helps to prevent constipation. They are important sources of mineral elements needed by the body being especially rich in calcium and iron. They are also valuable sources of vitamins. Green and yellow vegetables are important sources of vitamin A. These leafy, green and yellow vegetables contribute about 33% of the vitamin A supplied by the major food groups: they also supply about 25% of the Ascorbic acid and appreciable quantities of Thiamine, Niacin and Folic acid (Thompson and Kelley, 1957).
One of the main features of the diet taken by the inhabitants of the tropics and subtropics of the developing world is the deficiency in the supply of Protein and Vitamin. One of the measures recommended for the alleviation of this state is the increased production and consumption of cheaper sources of high quality protein and vitamin. The vegetables provide such a source, the potential of which can be immediately released (Egwuatu, 1975).
According to Tindall (1983) the leaves of Amaranthus (African spinach) per 100 g of edible portion contains Water, Protein, Fat, Mineral Salts like Calcium, Phosphorus Iron and Vitamin such as Carotene, Thiamin, Riboflavin, Niacin, Ascorbic acid.
Oke (1980) stated that Amaranthus is one of the most popular leafy vegetables in Southern Nigeria where it is grown for its leaves rather than seed. Indeed Amaranthus is also a major crop grown in the Fadama regions in the northern part of Nigeria (personal observation).
Kamalanathan et al. (1973) stated that among the different leafy vegetables grown in India, Amaranthus is the most popular on account of its earliness to mature, high nutritive value and palatability.
However, one of the greatest limiting factors in increasing the productivity of vegetable is the range of insect pests with which they are associated and the level of losses suffered in unimproved and improved agriculture.
There are numerous insect pests that have been recorded to attack this plant. Akinlosotu (1977) reported the followings as major pest of Amaranthus hybridus which is synonymous with A. cruentus: Sylepta derogata F. (Pyralidae: Lepidoptera) Hypolimnas missipus L. (Nymphalidae: Lepidoptera), Sjostedlina sp (Coreidae: Heteroptera). Cletus fuscescens Wlk, Cletus unifasciata Blote (Coreidae: Heteroptera) Anoplocnemis curvipes F (Coreidae: Heteroptera) Lygaeus festivus Thumb and Nysius albidus (Lygaeidae Heteroptera) Acrosternum acutum Dall, Aspavia armigera F., Menida maculiventris Dall in the family Pentatomidae order Heteroptera, Creontiades pallidus Ramb, Helopeltis schoutedeni. Reut (Mirirdae: Heteroptera) Aphis craccivora Koch, Aphis aspiraecola Patch (Aphidae Homoptera) Lagruia villosa F. (Lagriidae: Coleoptera), Oothetca mutabilis Sahl. (Galarucidae: Coleoptera) Epilachana chrysomelina var capensis Th. Coccinellidae: Coleoptera Gasteroclisus rhomboidalis Boh., Hypolixus nubilolus Boh., Alcidodes albolineatus Boh. and Nematocerus acerbus Fst. (Curculionidae: Coleoptera) Zonocerus variegatus Orthoptera: Pyrgomorphidae. He also mentioned the following: Hotea gambiae Wstw. (Pentatomidae: Heteroptera). Centrotus sp. (Membracidae: Homoptera) Homorocoryphus vicinus Wlk (Tettiganidae: Orthoptera) Cassida sp. (Cassidae: Coleoptera) as minor pests of A. cruentus in South Western Nigeria.
Forsyth (1966) reported that Hypolixus rhomboidalis (Coleoptera: Curculionidae) as an important pest of Amaranthus species in Ghana. Tindall (1983) also stated that Gasteroclisus rhomboidalis is one of the pests encountered on Amaranthus although he did not categorically say if it is a minor or major pest.
Most studies on the Amaranthus have been more or less on the consumption levels and nutrition value, cultivation practices centers of origin and diversity, germ plasm, collections, breeding and priorities for collecting and taxonomic classification and description (Grubben and Van Sloten, 1981).
Akinlosotu (1977), Forsyth (1966) and Tindall (1983) all these authors also list pests of Amaranthus whilst very little has been done on the various aspects of their bioecology.
According to Kumar (1987) knowledge of a pest’s basic biology and ecology is essential to gain an insight into the interaction of the pest with the components of the ecosystem in which it thrives. Also Wheatley (1972), a good understanding of the biology and behaviour of pest and disease organism usually reveal logical ways to attack them. Classically, the role of biologists have included identification of causal organism, elucidation of their lifecycles, habits, host ranges, modes of dispersal, migration and epidemology, their physiology and innate behaviour, host plant relationship and ultimately an understanding of the factors controlling their incidence.
The present study was undertaken to study some aspects of the bioecology and habit of Gasteroclisus rhomboidalis a curculionidae pest of Amaranthus, since lack of accurate biological information can have serious consequences in the development of integrated pest management programs.
Therefore the aim of this study is to investigate (i) the development of immature stages with a view to establishing the number of instar larvae and instar period. (ii) the food and host preference (iii) the feeding behaviour (iv) Boring activities and adult distribution per plants.
MATERIALS AND METHODS
An experimental plot of 12 vegetable beds 6x1.5 m each was established at the Applied Zoology Unit, Olabisi Onabanjo University Ago-Iwoye, Nigeria (6°48’N, 3°5’E). After the ground had been prepared, a part of the seed of A. cruentus was mixed with 1,000 parts of humus soil and sprinkled over the beds in rows thirty centimeters apart. The seedlings were watered in March before the commencement of rainfall when the experiment started but watering was stopped at the onset of rainfall late march.
For the laboratory studies of soil and 5 L plastic pots were filled with humus soil mixed with NPK fertilizer before transplanting some plants from the bed into the pots.
Seedlings of 3 weeks old were transplanted into 220 polythene bags, a plant per bag They were kept in the screen house for three weeks before inoculation and after inoculation.
Curculionid Pest Survey
A continual monitoring of curculionids found on Amaranthus was conducted on the experimental plot from March to September 2003.
Biology Studies of G. Rhomboidalis
A culture of G. rhomboidalis was set up in cages placed on the open verandah of the Laboratory, of the University where the temperature was approximate to air shade temperature for the University and fluctuated between 20.1 and 35.0°C and relative humility fluctuated between 50 and 96.7%.
Potted a Cruentus Plant Were Put in the Cages 1x0.7x0.7 m (L x B x H)
Adults beetle of both sexes were hand picked from the foliage of Amaranthus stand on the experimental plot.
To inoculate host plants with eggs, these beetles were confined in pairs, in the cages. Eggs were successfully laid and new potted plant replaced each day. Treated plants were promptly labeled, dated and transferred into another screen house to prevent reinoculation. This was done for a period of 40 days. Destructive sampling was done for the plant after the first set of adult emerged.
Artificial Rearing
Eggs reared artificially by dissecting the host plant for eggs found carefully removed with camel hair brush and placed on moist filter paper laid on wet cotton wool inside Petri dishes as used for Lixus camerunus (Eluwa, 1979). Egg dishes were kept on shelves in the laboratory at temperature fluctuating between 20 and 30°C during the study.
The above method was used to rear larvae and pupae dissected out from the plant to adult.
Morphometric
Morphometric studies and description of the immature stages of G. rhomboidalis was carried out on the various stages obtained and preserved in 70% alcohol.
Measurements of the egg length and width measurement of the larvae length, width, lead width, were done using micrometer eyepiece.
Behavioural Studies
Mating and oviposition were made on the adult both on the field and in the laboratory. Ten pairs were observed and observation carried out for 12 h on each.
Observations were also made on the periodicity of oviposition among the laboratory specimens. Choice of oviposition sites was observed using 15 pairs of G. rhomboidalis kept in cages.
Food Preference Tests
Preference of Amaranthus cruentus, Celosia argentes and Talinum triangulare for food was conducted for two weeks among 14 newly emerged adults.
RESULTS
Survey of Curclionid on Amaranthus Cruentus
In the survey carried out on the experimental plot at the University. Three species of curculionids were found. Gasteroclisus rhomboidalis Hypolixus nubilosus and unidentified very small curculionid were found.
The unidentified curculionid was most abundant followed by G. rhomboidalis 920, 679, 605 for the six month period.
Biology Studies of G. Rhomboidalis
Eggs
Table 1, summarizes the incubation periods of eggs and duration of larva instars of G. rhomboidalis under uncontrolled temperature and humidity conditions. Temperature fluctuating between 20.1 and 35.0°C and relative humidity between 50 and 96.77%. The eggs hatched in 7 days and the total period of development of egg to adult in 36±4 days. Percentage of egg hatched was low 67 eggs out of 128 eggs (52.34% hatchability).
Eggs placed in Petri dishes containing moistened cotton wool and filter paper hatched within 5 or 6 days. Thirty five eggs out of 40 eggs (87.5%) hatched within this period. Larva, there are three instar larval instars.
The duration of Pupa stage whilst the mean development period was 9 to 15 days in the shoot, it was 8 to 10 days in those reared in petri dishes.
Morphometric
Morphometric studies and description of the stages of G. rhomboidalis . EGG: The eggs are usually oval, partially white, measured 0.8 mm (±0.05 mm) long, (Table 2) 0.6±0.03 m wide. The head region of larva becomes clearly visible at the third day.
The egg is smooth with no architectural design. The egg colour changed from white when laid to creamy yellow before eclosion of G. rhomboidalis.
| Table 1: | Incubation period of eggs and duration of larval stages of G. rhomboidalis boh |
 | |
| Table 2: | Mean length of each of development stages of G.rhomboidalis |
 | |
| Table 3: | Mean head capsule width of larvae instars of G.rhomboidalis |
 | |
Larval Stages
There are three larval stages using the mean measurement of the head capsule of all representative larvae.
The three larval instars all looked alike and not significantly different in morphology to the first instar larvae.
1st Instar
On emergence it is white in colour, the head is brown, with prominent compound eyes and mandibles. The larva is apodous and eruciform. The abdomen has 10 segments, Head width 0.875±0.149 mm and the head is prognathous (Table 3).
Pupa
It is adecticous of the execrate type. Length 11.3±0.42 mm pupation is within the food plant. It is light brown at first, darkening later and heavily cuticularised at exposed surface like the appendages and rostrum.
Adult
It is uniformly black with distinct regular yellow deposit in bands. The rostrum is curved slender and subequal to the length of the thorax dorsally. Length 13.5±0.9 mm usually elongated oval and moderately robust. The lateral margin of the elytra is parallel medianly and broadly rounded apically. The elytra are often individually rounded apically.
The surface of the elytra is crenulate and the intervals consistently dull. The intervals have shallow punctures and numerous pubescences.
Behavioural Studies
Mating and Oviposiiton in G. rhomboidalis
Courtship and mating takes place anywhere on the plant, on top of leaf surface, stem and the inflorescence, in fact anywhere on the shoots as well exposed sites.
The female appears passive to both courtship and mating. Male mounts the female by approaching from behind. The male then straighten its forelegs elevating the anterior part of its body which assumes an angle of approximately 30° with that of female. After this, the male begins to stroke the head of the female with it rostrum. At the same time tarsi of the male start a rhythmic tapping on the thorax and abdomen of the female. The duration of this stroking varied and often occurs at irregular interval while mating occurs.
Once the male had induced the female to open her pygidium and overt the tip of her genitalia, the male places his aedeagus into the opening.
Copulation last several hours and have been observed to last for up to 12 h usually 9 h (n = 20 ![]() 8.55±2.37 h). Copulation among the insect take place frequently and at all times of the day.
8.55±2.37 h). Copulation among the insect take place frequently and at all times of the day.
It was observed that females do carry the males on their back without copulating and sometimes after copulating with the aedeagus withdrawn for hours.
Female oviposit just immediately after separating from the male. The female looks for a suitable site on the stem or a stout petiole wherein to oviposit. The female excavate or drill into the petiole or stem with its rostrum until the whole length of the rostrum is sunk into the stem, it then turns around, locate the hole with the tip of her abdomen and oviposits an egg.
| Table 4: | Mean tunnel length of immature stages of G.rhomboidalis |
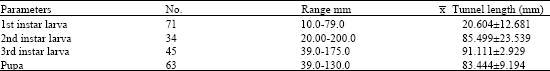 | |
The female again turns around pushes the egg down the hole with her rostrum and plugged with pulverished plant tissues and some secretions from its salivary gland.
Both laboratory and field observation showed that G. rhomboidalis prefer laying more of its eggs on the stem rather than the leaf petiole under 2% were found on the petiole ( 3 out of 193 punctures i.e., 1.5% observed on the petiole) of course these were thick petiole and no eggs were deposited at the inflorescence region. Eggs were out and often female starts laying its eggs by making the excavation or egg holes form the lower portion of the shoot about two thirds (2/3) from the terminal.
Feeding Habit
The adults feed on the leaf margin and leave a characteristic crescent scooped out sign or pattern at interval.
From field observation at the vegetable plots where Amaranthus cruentus was planted in close proximity to Celosia argentea on infestation of G. rhomboidalis was found on C. argentea. Likewise A. cruentus was preferred food of G. rhomboidalis on the experimental plot at the University where Talinum triangulare was left as weed among A. cruentus.
However, in a multiple choice test where in 14 newly emerged adults starved for 24 h were given a free choice of A. cruentus, T. triangulare and C. argentea in a cage, G. rhomboidalis showed a preference to C. argentea by first defoliating the two potted plants of C. argentea within 4 days T. triangulare did not feed on A. cruentus plant at all.
Boring Activities
Table 4 shows the mean tunnel length occupied measured for the larval to develop to adult. On hatching, the larva bores directly up into pith of the stem. When the eggs are laid in a leaf petiole, the larva on hatching tunnels down the petiole into the stem. Larvae bore in both directions up and down the stem. Often, those near the base bore towards the inflorescence.
Dissection of infected stems showed that developing larvae mined and consumed most of the pith of their host plants and in the process produced larval galleries in which the resulting pupae ultimately came to lie.
At the time of emergence of the adult, the individual larval gallery was about 32 to 150 mm long (n = 280, ![]() 48.914±12.94 mm). Dissection also revealed that within the stem, developing larvae usually mined upwards and downward thus leaving all discarded materials as scum at both ends of the larval chamber.
48.914±12.94 mm). Dissection also revealed that within the stem, developing larvae usually mined upwards and downward thus leaving all discarded materials as scum at both ends of the larval chamber.
Multiple developments of larvae do occur readily in individual host shoots and up to twelve developing larvae and pupae (m = 26 ![]() 6.46±3.71) have been found in a shoot.
6.46±3.71) have been found in a shoot.
Dissection revealed that no two finite larval chambers however close were in practice confluent, they were always separated by either a narrow strip of intact host tissues or at the worst, by a thick plug of woody fragments produced by the larvae. Thus no two advanced larvae or the succeeding pupae were ever found in one larval chamber.
DISCUSSION
The investigations undertaken in this work have attempted to elucidate some aspects of bioecology and behaviour of G. rhomboidalis on A. cruentus.
Apart from a brief description of the adult by Epenhuijsen (1974) and mention of it as pest of Amaranthus and Celosia by Akinlosotu (1977) and Tindall (1983), no detailed study of the behaviour nor the bio-ecology of G. rhomboidalis seems to have been published in Nigeria.
The survey showed that 3 curculionids, H. nubilosus, G. rhomboidalis and unidentified curculionid occur at the University vegetable plots. It is likely that these are all stem borers, as only the adults were encountered during the survey, much work need to be done on other two curculionids.
In all localities and varieties investigated, G. rhomboidalis were found on and collected from tender shoots of the Amaranthus leaf or on the inflorescence when the plant is fully matured. Literally, the species appeared to be constant inhabitant of this host plant. The distribution of adult beetle amongst host plant shoot was zero to four per shoot.
From the table on the mean tunnel (Table 4) there seems to be no relationship between tunnel length and stage of development, rather, the length of plant, the number of eggs deposited and the number hatched played a vital role in delimiting the length of the tunnel.
Attempts to establish the exact number of larval moults undergone by this weevil through a count of moulted head capsule found in the larval galleries as in other boring beetle fail as found by Eluwa (1979). This was because thorough examination of gallery debris revealed no definite head capsule except that of the penultimate larva mixed with brownish continuous plate presumably disintegrated moulted head capsules of the other instars as in the of Lixus camerunus Eluwa (1979).
However, since it is often desirable in insect life history work to know the period of such larval instar, that is the length of the several stadia, the width of the head capsule of all representative larvae found after hatching were measured. In fact, Bedard (1933) as quoted by Eluwa (1979) stated that since there is no anatomical characters whereby the various instars can be distinguished from one another and, the size of the body is so variable, the capsule dimensions are the only criteria for identification.
Therefore, using the head capsule measurement of G. rhomboidalis and incorporating it with the larval length measurement there seems to be only three instars.
This is similar to the number of instars larvae obtainable in other curculionid such as in the genus Baria and in Neochetina bruchi (Deloach and Cordo, 1976) and in Tryophloeus striatus (Furmiss, 2004) but in contrasts to five to seven instars in Hylobitelus xiaoi by Wen et al. (2004).
From the result therefore, the beetle spends 7 days in the egg stage 10 to 21 days as larva (![]() = 12.55±5.85 day) and 9 to 15 days pupa (12±2) and the life history stages completed in 32 to 40 days from oviposition to emergence.
= 12.55±5.85 day) and 9 to 15 days pupa (12±2) and the life history stages completed in 32 to 40 days from oviposition to emergence.
From observation, the onset of rain which marks the termination of the dry season encourages the emergence of some weeds among which is Amaranthus spinosus and abandoned farms of fallow field thus flourish for several weeks before sufficient moisture in the soil allow for cultured planting. These probably allow easy build up of the insect which survive on this plant where it survives as alternative host.
At present, Amaranthus is grown during the wet season of the year except near the river banks or where it can be irrigated. Thus its time of planting needed to be studied as early planting during the beginning of wet seasons may result in concentration of early migrants or early emerging stem borers on the early planted crops as found in the Rice stem borer by Akinsola (1979).
Rearing these insects artificial decreases the period of development and seen that it took 5 to 6 days for eggs to hatch in the Laboratory while 7 days in the shoot likewise took pupa 9 to 15 days before emergence in the shoot where as, in the Laboratory took 8 to 10 days and from the treatment given the Laboratory (i.e., moisten cotton wool) high humidity plays a vital role in the rapid development of the insect. Indeed the eggs of Lucilia sericata Meig (Bursell, 1970) and Ptinus tectus Boisd (Chapman, 1972) demonstrate a type of relationship in which high relative humidities accelerate development.
This stem borer of Amaranthus do not usually destroy all stem within a plot and for this reason, it is often difficult to accurately estimate yield losses as found in the Rice Stem borer (Akinsola, 1979). Complete death of plant is caused by infestation at the early stage of growth when the girth of plant is small and infestation in form of ovipositioning is high on the plant.
From the ecological studies on this beetle, it seems G. rhomboidalis may be referred to as an indirect pest as Ruesink and Kogan (1975) stated that the indirect pest, damage parts that may not affect yield, according to the role of the part in the physiology of the plant vis a vis the development of desired product. Thus when the desired product is the leaf, the beetle consumption is very low especially when it is realized that their number per matured plant is zero to four.
On the other hand, it may be referred to as a direct pest when the larval tunneling activities is taken cognizance of as the profuse tunneling toward the terminal and oft the shoot resulted in shoot break over and hence production of grain (seed) is seriously affected.
It is persistent pest in that development of this insect still continue in the plant host weeks after these are uprooted and the aging and withering host plant do serve readily for the development of larval stages.
Amarathus in Nigeria is largely under a low level technology and apart from large scale government farms, insecticides are hardly used. Even the effectiveness of insecticide on this stem borer that securely shut off its immatured stages from external influence needs much to be determined.
Therefore a study of this nature can only be a background information and the study needs to carried out on longer period of time and it would be of great value to continue to study the incidence and relationship of stem borers and their natural enemies as the planed extensification and intensification of vegetable production in Nigeria progresses.This study therefore provide the much needed background information for integrated control of this pest.
REFERENCES
- Furmiss, M.M., 2004. Biology of Trypophloeus striatulus (Coleoptea Scolytidae) in felt leaf willow in Interior Alaska. Environ. Entomol., 33: 21-27.
Direct Link - Wen, X., Y. Kuang, M. Shi, H. Li, Y. Luo and R. Deng, 2004. Biology of Hylobitelus xiaoi (Coleoptera Curculionidae), a new pest of slash Pine, Pinus elliottii. J. Econ. Entomol., 97: 1958-1964.
Direct Link