
G.A. Foko Dadji
Laboratory of Zoology, Faculty of Science, University of Yaounde I, P.O. Box 812 Yaounde, Cameroon
J.L. Tamesse
Laboratory of Zoology, Higher Teacher�s Training College, University of Yaounde I, P.O. Box 47 Yaounde, Cameroon
J. Messi
Laboratory of Zoology, Faculty of Science, University of Yaounde I, P.O. Box 812 Yaounde, Cameroon
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 4 | Page No.: 299-307







ABSTRACT
The developing trend of resistance in Anopheles gambiae toward synthetic mosquitocidal agents makes their management extremely difficult. A new approach in the fight against malaria vectors consist of using native plants with insecticidal value. Two varieties of Capsicum annuum the red and the yellow varieties were evaluated for their toxicities to An. gambiae. Ovicidal, larvicidal and pupal mortality effects of C. annuum fruit have been noted. The results showed that for 60 eggs, embryonic mortality was 100% when treated with the yellow variety of C. annuum. In addition, this variety was also detrimental to the embryonic development of An. gambiae. In contrast, at a concentration of 1 g L-1 of the red variety of C. annuum 19% of egg-hatched and 93% of total mortality rate from eggs to emergence. At a concentration of 0. 25 g L-1 of the red variety of C. annuum 44% of egg-hatched and 67% of total mortality rate from eggs to emergence. The embryonic development duration and first instars larvae development duration were longer (82 and 77 h, respectively) in media containing 1 g L-1 of C. annuum. The total mortality rate and the quantity of C. annuum in breeding media were highly correlated (r = 0.964; p< 0.01). The number of males, females and the number of eggs laid per female were negatively correlated with the quantity of C. annuum in the breeding medium. This study shows that C. annuum fruits are significantly toxic on eggs, larval and pupal stages of An. gambiae. Future tests in the field will help determine if C. annuum may be effective for malaria control in tropical region.
PDF Abstract XML References Citation
How to cite this article
G.A. Foko Dadji, J.L. Tamesse and J. Messi, 2007. Insecticidal Effects of Capsicum annuum on Aquatic Stages of Anopheles gambiae Giles under Laboratory Conditions. Journal of Entomology, 4: 299-307.
DOI: 10.3923/je.2007.299.307
URL: https://scialert.net/abstract/?doi=je.2007.299.307
DOI: 10.3923/je.2007.299.307
URL: https://scialert.net/abstract/?doi=je.2007.299.307
INTRODUCTION
The malaria morbidity and mortality rates in Africa began an upward trend in the 1970s probably due to an increasing of parasite resistance to chloroquine and resistance of the vector mosquitoes to chemical insecticides (Carter and Mendis, 2002; Korenromp et al., 2003; Terkuile et al., 2004). Most human infections of Plasmodium are due to the infecting sting of mosquito of the Anopheles genera. Anopheles gambiae, Anopheles funestus and Anopheles arabiensis are the most important vectors of malaria in Africa (Coluzzi, 1984; Fondjo et al., 1992; Mouchet and Carnevale, 1991). In Cameroon, An. gambiae is the main vector of malaria in the rural and urban regions (Manga et al., 1992). Currently, Insecticide-Treated Nets (ITNs) represent the most practical and economical methods of controlling vectors (Copeland et al., 1995).
Although the fight against malaria vectors is the most efficient preventative way to reduce the incidence of this endemic illness (Carnevale and Mouchet, 1990); chemical insecticides (e.g., DDT, dieldrine…) used in the past proved inefficient due to the detrimental effect to the environment, the development of pesticide resistant mosquitoes and the escalating cost of application. Products used were inefficient for the vector control in various regions of Africa South of Sahara. The control of populations of Anopheles by plants extract with insecticidal effects is one of the new tracks of research (Rey et al., 2001). Some plants are source of substances used to repulse or to kill mosquitoes and other insect pests. Odalo et al. (2005) showed that Croton pseudopulchellus Pax, Mkilua fragrans Verdc, (Annonaceae), Endostemon tereticaulis (Poir.) Ashby, Ocimum forskolei Benth., Ocimum fischeri Guerke, Plectranthus longipes Baker (Labiateae) and Cymbopogon nardus Melissa (Curtis et al., 1989) have been demonstrated to exhibit good repellent activities against Anopheles gambiae. The recent plant- insect interactions studies reveal that some secondary metabolites of plants could have an effect on the growth, the development and the behavior of larvae (Muller and Su, 1999). Among these, several phenolic compounds are known for their toxicity to insects (Mercer and Anderson, 1994) including carvacrol, 4-isopropylbenzenemethanol, phytol, thymol 1-methylpyrrole which exhibited high individual repellencies to An. gambiae (Odalo et al., 2005).
Capsicum annuum (Solanaceae) is cultivated in all regions of Cameroon. Its fruits that are used as spices are very rich in phenolic compounds (Diaz et al., 1998). The most abundant of these compounds are the capsaicinoids, including capsaicine and dihydrocapsaicine which represent 77 to 90% of secondary compounds. These compounds have been used to control bugs and birds (Diaz et al., 1998). In addition, these compounds are biodegradable without any harmful effect on the environment (Isman, 1999).
In the study reported here, we determined the possible effects of C. annuum at varying concentrations on mosquito growth and development. We examined the inhibition effect of two varieties of C. annuum on egg-hatching, larval and pupal development of An. gambiae in an effort to find a biological control agent that is benign to the environment, but will effective by reduce the multiplication of the malaria vector.
MATERIALS AND METHODS
Mature fruits of Capsicum annum Linne, commonly called “thick pimento” were obtained from the Agronomic Research Institute for Development (Yaounde-Cameroon). Fruit of both the red and yellow varieties were obtained dried and crushed with a moulinex to form a fine powder. The resultant powder was added to the mosquito breeding medium.
Breeding of An. gambiae
Eggs were obtained from laboratory strain of An. gambiae maintained at ambient rearing condition. All the bioassays were conducted at 26-28°C, 70-80% RH, 12:12(L:D), in 2003 and in Yaoundé1 University (Yaoundé, Cameroon).
Sixty eggs of An. gambiae were added to each of three plastic trays (18 cm Ø, 5 cm deep) containing 1, 2 and 4 L of spring water from Simbock (locality near Yaounde) with a sufficient quantity of Tetrababy fish food “R” (0,3 g/100 larvae/d). This food is rich in proteins and usually used for the nutrition of young fishes of less than 1 cm of length.
Three rearing media were constituted containing the powder of Capsicum annuum:
| • | Medium 1: One liter of spring water containing 1 g of the powder |
| • | Medium 2: Two liter of spring water containing 1 g of the powder |
| • | Medium 3: Four liter of spring water containing 1 g of the powder |
Control mediums consisted of (1, 2 and 4 L) of spring water without C. annuum powder. The number of eggs hatched was determined daily by counting the number of first instar larvae observed in each tray until no hatching occurred for 10 days. The duration of embryonic development was determined using the method reported by Dempster (1961). Embryonic development was calculated by the time it takes for 2/3 of the eggs to become first instar larvae. The experiment took 20 days and was replicated 10 times in order to eliminate possible tray effects or cage effects.
Pupae and larvae of An. gambiae are counted daily. At emergence, adults were counted and their sex was determined by visual inspection. Sex ratio was determined (number of female per the number of male that emerged). A sugar solution (10%) was provided ad libitum. Three days after emergence, females were allowed to blood feed on a rabbit and kept under laboratory conditions to lay eggs. Eggs laid by females were counted and the average fecundity for each female was estimated (total number of eggs laid over the total number of females). The total duration of development (from eggs to adults) was determined by considering embryonic development duration, larval development duration and pupal development duration. Throughout our experiment, mortality rate was determined at each developmental stage (number of death over the total number of larvae or pupae at the developmental stage). Larvae were considered dead when they did not react to touching. Adult mortality rate was determined 5 days after emergence by divided number of death at the punter with the total number of adult recorded immediately after emergence.
Analyses of Data
Mean and standard errors of different parameters were calculated and compared using SPSS (SPSS, 1999; Tripathi et al., 2003) notably tests of comparison non parametric Z of Wilcoxon (1945), U of Mann Whitney (1947) and χ2 of Kruskal Wallis (1952) and the test of interrelationship of Spearman.
RESULTS
Effect of Capsicum annuum on Egg-hatching, Larval and Pupal Development Duration
No egg hatching was recorded for each quantity of C. annuum (yellow variety) (1, 0.5, 0.25 g L-1). This variety didn’t permit us to obtain any larvae during the ten series of experiments in which 60 eggs were tested. Each experiment took 7 days.
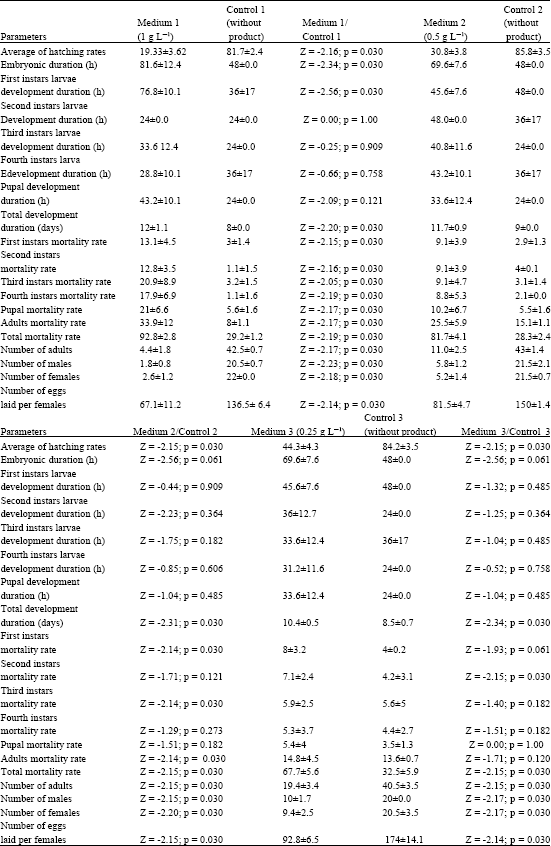
The egg-hatching rate varied within the quantity of C. annuum of red variety. It could be seen from Table 1, that percentage of hatched eggs was 19.33% in the medium with 1 g L-1 of C. annuum (so called medium 1), 30.83% in the medium with 0.5 g L-1 of C. annuum (medium 2) and 44.33% in the medium with 0.25 g L-1 of C. annuum (medium 3). The embryonic development duration was about 81.6 h in the medium 1; 69.60 h in the medium 2 and 69.6 h in the medium 3.
The larval and pupal development duration was as followed: first instars larvae: 76.8 h in the medium 1; 45.6 h in the medium 2 and 45.6 h in the medium 3; second instars larvae: 24 h in medium 1; 48 h in medium 2 and 36 h in medium 3; third instars larvae: 33.6 h in medium 1; 40.8 h in medium 2 and 33.6 h in medium 3; fourth instars larvae: 28.8 h in medium 1; 43.2 h in medium 2 and 31.2 h in medium 3; pupa: 43.2 h in medium 1; 33.6 h in medium 2 and 33.6 h in medium 3. The pre-adult development duration was 12 days in medium 1; 11.7 days in medium 2 and 10.4 days in medium 3 (Table 1).
Exposure of An. gambiae eggs and different larvae development stages to capsicum annuum powder of red variety showed a significant difference compares to the results obtained from control media. Indeed, results obtained in medium 1 compared to those of control medium showed a significant difference for the egg-hatching rate (Z = -2.16; p = 0.03), embryonic development duration (Z = -2.34; p = 0.03), first instars larvae development duration (Z = -2.56; p = 0.03) and the total development duration (Z = -2.207; p = 0.03). The comparison of the developmental parameters of An. gambiae obtained in medium 2 and its control medium and also results from medium 3 and its control medium permitted us to reveled that only the hatching rate (Z = -2.15; p = 0.03) and the total development duration (Z = -2.31; p = 0.03, Z = -2.34; p = 0.03, respectively) showed the significant difference (Table 1).
| Table 1: | Effects of C. annuum (red variety) on the development and mortality of An. gambiae and comparison between experimental media and their corresponding controls (non parametric test of wilcoson; p<0.05) |
 | |
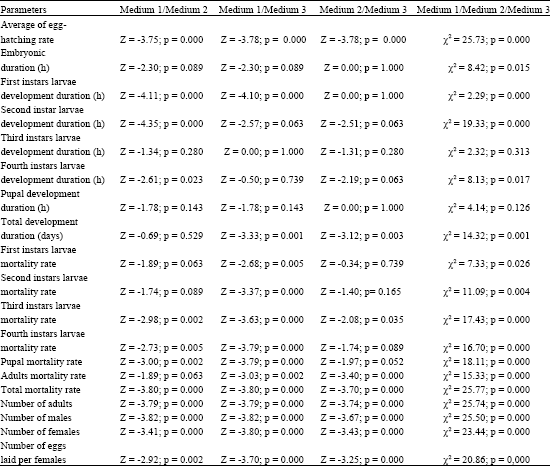
When comparing the effect of different quantities of Capsicum annuum of red variety on egg-hatching and development duration of larval and pupal stages until the adult reared in medium with those reared in the medium 2, we noted a significant difference: egg-hatching rate (Z = -3.75; p = 0.000); first instars larvae development duration (Z = -4.11; p = 0.000); second instars larvae development duration (Z = -4.35; p = 0.000); fourth instars larvae development duration (Z = -2.61; p = 0.023). The comparison of results obtain from medium 1 to those of medium 3 shows a significant difference for egg-hatching rate (Z = -3.78; p = 0.000), first instars larvae development duration (Z = -4.1; p = 0.000) and total development duration (Z = -3.33; p = 0.001). On the other hand, when we compared the results gotten in medium 2 to those of medium 3, only the egg-hatching rate (Z = -3.78; p = 0.000) and the pre-adult development duration (Z = -3.12; p = 0.003) showed a significant difference. Comparison between parameters of the development of this insect in the three media (medium 1, medium 2, medium 3) showed that only the third instars larvae development duration and the pupal development duration showed a non significant difference (Table 2).
The interrelationship test of Spearman achieved between the eggs hatching, the development duration of different larval stages and the quantity of Capsicum annuum of red variety in the rearing medium revealed a significant correlation between the hatching rate (r = -0.961; p <0,01), the embryonic development duration (r = 0.714; p = 0.01), the first instars larvae development duration (r = 0.754; p <0.05), the pre-adult development duration (r = 0.808; p <0.01) and the quantity of Capsicum annuum in the rearing medium (Table 3).
| Table 2: | Effects of C. annuum (red variety) on the development and mortality of An. gambiae and comparison between several experimental media containing different quantities of powder. (non parametric tests of wilcoson. Mann whitney and Kruskal wallis; p<0.05) |
 | |
| Table 3: | Interrelationship test of Spearman between egg-hatching rate, development duration, mortality rate of developmental stages of An. gambiae and the quantities of C. annuum powder in breeding media (p<0.05) |
 | |
| * Non significant, **: Significant | |
Effect of Capsicum annuum on Larval and Pupal Mortality
The mortality rate of An. gambiae larvae and pupae have been evaluated in the different rearing media. The number of males, females and the number of eggs laid a female who survived from pupae was also evaluated. The mortality rate of larvae was as followed: first instars larvae, 13.1% in medium 1, 9.1% in medium 2 and 8% in medium 3; second instars larvae, 12.8% in medium 1, 9.1% in medium 2 and 7.1% in medium 3; third instars larvae, 20.9 in medium 1; 9.1 in medium 2 and 5.9% in medium 3; fourth instars larvae, 17.9 in the medium 1, 8.8 in medium 2 and 5.3 in medium 3. The mortality rate of pupae was as follow: 21% in medium 1, 10.2% in medium 2 and 5.4% in the medium 3. The adult’s mortality rate was 33.9% in the medium 1, 25.5% in medium 2 and 14.8% in medium 3. The total mortality rate at emergence, known as the number of adults compared to the number of eggs at the beginning of each experiment, was 92.8; 81.7 and 67.7% in media 1, 2 and 3, respectively. Thus, the mortality rate is higher in medium 1 follow-up with medium 2 and then medium 3. At the end of the experiment the number of adults were 1.8 males and 2.6 females in medium 1; 5.8 males and 5.2 females in medium 2; 10 males and 9.4 females in medium 3. The number of eggs laid a female was, respectively 67.1; 81.5 and 92.8 in media 1, 2 and 3 (Table 1).
The mortality rate of the different development instars, the number of males and females and the number of eggs laid per female of An. gambiae showed a significant difference when we compared the results gotten in medium 1 to those of the control. In the other hand, between medium 2 and the control, we noted a significant difference for the first instars larvae mortality rate, the third instars larvae mortality rate, the adult mortality rate, the total mortality rate, the number of males and females and the number of eggs laid per female. Between the medium 3 and control we noted a significant difference for second instars larvae mortality rate, the total mortality rate, the number of males and females and the number of eggs laid per female (Table 1).
The comparison between the mortality rate of mosquitoes larvae and pupae reared in medium 1 to those observed in medium 2, showed a significant difference for the third instars (Z = -2.98; p = 0.002), fourth instars (Z = -2.73; p = 0.005), pupae (Z = -3.00; p = 0.002) and total mortality rate of adults at emergence (Z = -3.8; p = 0.000). Mortalities rates observed in medium 1 are meaningfully different from those observed in medium 3 (p = 0.000). The comparison between the mortality rate of mosquitoes larvae, pupae and adults reared in medium 3 to those of medium 2, show a significant difference for the third instars larvae (Z = -2.08; p = 0.03), adults (Z = -3.40; p = 0.000) and the total mortality rate of adults at emergence (Z = -3.7; p = 0.000). The comparison of mortalities rates is meaningfully different in the three media (p = 0.001). The number of adults, (males and females) and the number of eggs laid per female were meaningfully different from one medium to another (Table 2).
The interrelationship test of Spearman between the An. gambiae mortality rate and the quantity of C. annuum in the rearing medium was meaningfully significant for the first instars larvae mortality rate (r = 0.692; p = 0.001), the second instars larvae mortality rate (r = 0.741; p = 0.01), the third instars larvae mortality rate (r = 0.803; p = 0.01), the fourth instars larvae mortality rate (r = 0.776; p = 0.01), the pupal mortality rate (r = 0.776; p = 0.01), the adults mortality rate (r = 0.784; p = 0.01) and the total mortality rate of adults at emergence (r = 0.964; p = 0.01) (Table 3).
DISCUSSION
Protection against mosquitoes is generally obtained by the use of synthetic chemical products with the culminating problems associated with the development of resistance in mosquitoes and toxicity to man and its environment (Tripathi et al., 2003). However, natural product provided useful future alternative means for mosquito control. Some plant extracts or phytochemicals products are known to possess ovicidal, larvicidal, repellent, antifeeding and insecticidal activities against various insect species (Isman, 1999). Results of the current study revealed that egg-hatching rate decrease with the concentration of Capsicum annuum in the medium, meanwhile the total larval development duration increase with the concentration of C. annuum in the medium.
Secondary plant metabolic compounds (polyphenols) are known to have adverse effects on the midgut epithelial barrier of Lepidoptera and Orthoptera larvae (Barbehenn and Martin, 1994). Phenolic components are known to have ovocidal and insecticidal properties against different species of insects (Isman, 1999). Yang et al. (2003) showed that terpenoids and alkaloids destroy eggs and females of P. humanus capitis.
The capsaicin, major phenolic components contained in the fruits of C annuum although its insecticide activity have not yet been well defined, it has been suggested that it would inhibit the growth of insects and would be implied in certain reactions of oxidization (Diaz et al., 1998). In this study, pre-adult development duration is positively correlated to the quantity of C. annuum in the medium (r = 0.808). Our results showed that C. annuum contained the poisonous compounds responsible for the retardation of egg-hatching, larval and pupal development of An. gambiae. It can therefore be use for larval control. However this approach was neglected and malaria control policy shifted toward domestic adulticid methods. Nevertheless, it is important to remind that Brazil, Egypt an Zambian have successfully suppressed malaria for over 10 years by using larval control program (Killeen et al., 2002).
The mortality rate of An. gambiae at different developmental stages was higher when the concentration of C. annuum in the medium was 1 g L-1 and low when this concentration was 0.25 g L-1. The mortality rate thus recorded should be due to the chemical compounds of the C. annuum fruits. Rey et al. (2001) showed that this plant could produce few polyphenol and mainly capsaicinoids. Capsaicinoids are implied in the biosynthesis of the lignin (polyphenol). Larvae raised in a medium containing polyphenol ingest these poisonous compounds that accumulate in their intestine and destroy their partition epithelial (David et al., 2000b). The plant polyphenol showed a more elevated insecticide activity compared to the conventional insecticides (Rey et al., 2001; David et al., 2000a). This statement can explain the increase of their mortality rate and the affect of the punter of females (Tuno et al., 2004). Larvae of An. gambiae used to swim on to surface; when the medium is deep, they are obliged to flood.
With the yellow variety of C. annuum, we got 0% of egg-hatching rate. The main characteristic of pepper is their pungent flavor due to capsaicin and a related molecules, capsaicinoid. The quantity of these compounds varied within varieties of C. annuum, 1 to 4% (Rey et al., 2001). So the most efficient varieties are those, which have a high rate of capsaicinoids that would be the case of the yellow variety of C. annuum.
The total mortality is greatly correlated to the quantity of C. annuum in the medium of breeding. Although the World Health Organization considers that the larval control in Africa should have an applicability limited in Africa. Strategies of control of aquatic stages of An. gambiae have been the biologic struggle. Then for this control we recommend that the drainage or the drying up of larva habitats and or the immersion in these larval resting places of poisonous compounds of plant origins as C. annuum. According to our result C. annuum can be considered as a good material for the control of malaria vectors with no poisonous effect on human being and the environment.
CONCLUSIONS
The study of the development duration and the mortality rate of An. gambiae in a medium containing the powder of Capsicum annuum permitted to note that the development duration were longer when higher quantity of C. annuum were added in the breeding medium. The mortality rate was positively correlated to the concentration of C. annuum. Therefore, the immersion of powder of this plant in the resting places would be an efficient method for malaria vector control, particularly in the developing countries where mosquitoes larvae developed resistances facing the chemical insecticides. Larval control methods should now be reprioritized for research, development and implementation as an additional way to roll back malaria.
REFERENCES
- Barbehenn, R.V. and M.M. Martin, 1994. Tannin sensitivity in larvae of Malacosoma disstria (Lepidoptera): Roles of the peritrophic envelope and midgut oxidation. J. Chem. Ecol., 20: 1985-2001.
Direct Link - Carnevale, P. and J. Mouchet, 1990. [Vector control and malaria control]. Medecine Tropicale, 50: 391-398.
PubMed - Carter, R. and K. Mendis, 2002. Evolutiony and historical aspects of the burden of malaria. Clin. Microiol. Rev., 15: 564-594.
Direct Link - Coluzzi, M., 1984. Heterogeneities of the malaria vectorial system in tropical Africa and their significance in malaria epidemiology and control. Bull. World Health Organ., 62: 107-113.
Direct Link - Copeland, R.E., T.W. Walker, L.L. Robert, J.I. Githure, R.A. Wirtz and T.A. Klein, 1995. Response of wild Anopheles funestus to repellent-protected volunteers is unaffected by malaria infection of the vector. J. Am. Mosquito Contr. Assoc., 11: 438-440.
Direct Link - David, J.P., D. Rey, G. Marigo and J.C. Meryran, 2000. Larvicidal effect of a cell-wall fraction isolated from alder decaying leaves. J. Chem. Ecol., 26: 901-913.
Direct Link - Dempster, J.P., 1961. The analysis of data obtained by regular sampling of an insect population. J. Anim. Ecol., 30: 429-432.
CrossRefDirect Link - Diaz, J., A. Bernal, F. Merino and A. Ros Barcelo, 1998. Phenolic metabolism in Capsicum annuum L. Recent Res. Devel. Phytochem, 2: 155-169.
Direct Link - Killeen, G.F., U. Fillinger, I. Kiche, L.C. Gouagna and G.J. Knols, 2002. Eradication of Anopheles gambiae from Brazil: Lessons for malaria control in Africa?. Lancet Infect. Dis., 2: 618-627.
CrossRefPubMedDirect Link - Korenromp, E.L., B.G. Williams, E.C. Gouws, C. Dey and R.W. Snow, 2003. Measurement of drends in childhood malaria mortality in Africa: An assessment of progress toward targets based on verbal autopsy. Lancet Infect. Dis., 3: 349-358.
Direct Link - Kruskal, W.H. and W.A. Wallis, 1952. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc., 47: 583-621.
CrossRefDirect Link - Mann, H.B. and D.R. Whitney, 1947. On a test of whether one of 2 random variables is stochastically larger than the other. Ann. Math. Stat., 18: 50-60.
CrossRefDirect Link - Mercer, D.R. and I.R. Anderson, 1994. Tannins in treehole habitats and their effects on Aedes sierrensis (Diptera: Culicidae) production and parasitism by Lambornella clarki (Ciliophora: Tetrahymenidae). Ann. Entomol. Soc. Am., 31: 159-167.
CrossRefDirect Link - Mulla, M.S. and T. Su, 1999. Activity and biological effects of Neem products against arthropods of medical and veterinary importance. J. Am. Mosq. Control Assoc., 15: 133-152.
PubMedDirect Link - Odalo, J., M. Omolo, H. Malebo, J. Angira, P. Njeru, I. Ndiege and A. Hassanali, 2005. Repellency of essential oils of some plants from the Kenyan coast against Anopheles gambiae. Acta Trop., 95: 210-218.
CrossRef - Rey, D., J.P. David, G. Besnard, J.L. Jullien, C. Lagneau and J.C. Meyran, 2001. Comparative sensitivity of larval mosquotoes to vegetable polyphenols versus conventional insecticides. Entomol. Exp. Appl., 98: 361-367.
Direct Link - Ter Kuile, F.O., M.E. Parise, F.H. Verhoeff, V. Udhaya-Kumar and R.D. Newman et al., 2004. The burden of co-infection with human immunodeficiency virus type 1 and malaria in pregnant woman in sub-Sahara Africa. Am. J. Trop. Med. Hyg., 71: 41-54.
Direct Link - Tripathi, A.K., V. Prajapati, A. Ahmad, K.K. Aggarwal and S.P. Khanuja, 2004. Piperitenone oxide as toxic, repellent and reproduction retardant toward malarial vector anopheles stephensi (Diptera: Anophelinae). J. Med. Ent., 41: 691-698.
Direct Link - Tuno, N., K. Miki, N. Minakawa, A. Githeko, G. Yan and M. Takagi, 2004. Diving ability of Anopheles gambiae (Diptera: Culicidae) Larvae. J. Med. Entomol., 41: 810-812.
Direct Link - Yang, Y.C., H.S. Lee, J.M. Clark and Y.J. Ahn, 2004. Insecticidal activity of plant essential oils against Pediculus humanus capitis (Anoplura: Pediculidae). J. Med. Entomol., 41: 699-704.
CrossRefPubMedDirect Link