
R.K. Gupta
Division of Entomology, Faculty of Agriculture, Sher-e-Kashmir University of Agriculture Sciences and Technology-Jammu, 180002, India
J.C. Raina
Division of Entomology, Faculty of Agriculture, Sher-e-Kashmir University of Agriculture Sciences and Technology-Jammu, 180002, India
M.D. Monobrullah
Division of Entomology, Faculty of Agriculture, Sher-e-Kashmir University of Agriculture Sciences and Technology-Jammu, 180002, India
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 4 | Page No.: 279-288







ABSTRACT
Optimization of virus yield is crucial for minimizing the cost of production and to perfect its large scale-production using in vivo production system. Therefore, separate experiments were conducted to optimize the inoculum dose, larval age, incubation period, and physical status of the harvested larvae for in vivo Helicoverpa armigera nucleopolyhedrosis (HaNPV) mass production. Seven to eight day old larvae treated with an inoculum dose of 1x104 resulted in maximum POBs yield/insect coupled with the desirable proportion of larval harvest and shall therefore be considered as suitable stage for optimum virus production with an incubation period of 7 days. While comparing the HaNPV productivity on the basis of the physical status of the harvested larvae, the maximum POB yield per larva was obtained by harvesting the dead larvae. However, harvesting dead larvae was very difficult and there was high microbial contamination. Thus harvesting moribund larvae was the best option for optimum production of HaNPV.
PDF Abstract XML References Citation
How to cite this article
R.K. Gupta, J.C. Raina and M.D. Monobrullah, 2007. Optimization of in vivo Production of Nucleopolyhedrovirus in Homologus Host Larvae of Helicoverpa armigera . Journal of Entomology, 4: 279-288.
DOI: 10.3923/je.2007.279.288
URL: https://scialert.net/abstract/?doi=je.2007.279.288
DOI: 10.3923/je.2007.279.288
URL: https://scialert.net/abstract/?doi=je.2007.279.288
INTRODUCTION
Insect baculoviruses are particularly attractive as bio-insecticides because of two factors. They are safe for vertebrates and other non-target fauna and they are generally highly pathogenic, host death being the most likely outcome of an infection. Although their use for control of insect pest populations was demonstrated in the 1940s, the first registered commercial product did not appear for another 30 years. Since then, a number of commercial products have been developed (Copping, 1993) but most could not be sustained in market due to the high cost of production. Several factors influence virus production, including: host insect age and sex, rearing environment and nutritional quality of the diet (Shapiro et al., 1981; Kalia et al., 2001; Subramanian et al., 2006). In general, any factor that influences the larval growth rate after virus inoculation will influence the virus yield (Ignoffo and couch, 1981). Optimal production of the viruses is desired for greatest yield of biologically active virus; this requires selection of the appropriate larval age for infection and standardization of inoculum dose to permit maximum larval growth, resulting in higher yield/larva, among other parameters (Shieh, 1989). Besides, it should also confirm to the requisite quality control standards with minimum to negligible contamination of in vivo systems with bacteria and other microbes (Shapiro et al., 1986).
As insect viruses are obligate pathogens, it is necessary to produce them either in host larvae (in vivo) or in cell culture (in vitro). In order to be competitive with traditional pest control methods, baculovirus production must be cost effective. Whereas the in vitro system is not preferred due to its high cost of production (Inceoglu et al., 2001), in vivo production, most often in the homologous host reared on artificial diet, is the only option for large-scale production of the virus in third world countries where inexpensive labour is available and insecticides are expensive (Shieh, 1989; Rabindra et al., 2003). Production in vivo is straight forward, but yields can be highly variable, as can costs (Cherry et al., 1997) which may range from $ 3.33 to 5.33/100 mL of finished product containing 1x1011 Polyhedral Occlusion Bodies (POB)/100 mL (Sathiah and Palaniswamy, 2004). This variability in production cost is critical to the economic success or failure of baculovirus production. The widespread use of Helicoverpa armigera Nuclear Polyhedrosis Virus (HaNPV) in India is well documented as the demand for commercial formulation of this virus is ever increasing, but its commercial use has not percolated to northern parts of country due to lack of suitable regional isolate (Rabindra et al., 2003). In order to promote and popularize its use in north western India, the use of local strains with better adaptability and efficacy has been successfully demonstrated (Rabindra et al., 2003; Gupta et al., 2003). However, for its wide spread multiplication and commercial use as novel biopesticides optimization of in vivo production is very crucial. One of the problems of the original production method, that involved harvesting many infected larvae as dead insects, was that these were often heavily contaminated with bacteria which were found difficult to remove (Grzywacz et al., 1998). Also, alternative inoculation method to obtained higher yield of virus through larval spiracles (Kalia et al., 2001) can be rarely practiced for commercial production. Optimization of in vivo production for Spodoptera litura or other pests is well documented. but the present information on the optimization of HaNPV, is inconsistent. This paper describes the work done to optimise the production of viruses with respect to the most crucial factors and to regain higher levels of production with minimal contamination at a competitive cost.
MATERIALS AND METHODS
Rearing of Host Insect
A disease-free colony of H. armigera was maintained on a semi-synthetic diet (Shorey and Hale, 1965) at 26±2°C and 70±10% relative humidity, 10:14 h (L: D) photoperiod in biocontrol Laboratory of SKUAST-Jammu.
Source of Virus
The HaNPV used in the present study was a purified stock suspension of a local isolate (Samba) which was originally isolated from outer plains of Jammu in 2001 and maintained in the insect Pathology Laboratory. The isolate was given serial passages in laboratory-reared 3rd instar H. armigera larvae that were free from any detectable NPV infection. The virus was propagated for further studies following adherence to strict microbiological procedures by harvesting and collecting larvae individually in sterilized microfuge tubes immediately after death to minimize bacterial contamination followed by storage at -20°C. Larvae that remained intact were harvested and macerated with 3 mL of 0.1% Sodium Dodecyl Sulphate (SDS) and filtered through three-layered muslin cloth several times to remove the insect debris. Further, purification was done by centrifugation of filtrate at 500 rpm for 1 min and discarding the pellet of tissue debris. The supernatant thus obtained was subjected to centrifugation at 2500 rpm for 5 min and supernatant containing lipids and soluble material was discarded. The resuspended pellet containing the occlusion bodies was further purified on 45-60% (w/w) sucrose gradient in a zonal rotor and the fraction containing POBs was diluted in appropriate quantity of 0.1% SDS, pelleted and washed in sterile distilled water. Prior to enumeration, the contents was vortexed to facilitate equal distribution of polyhedra in the suspension and the stock suspension was diluted in sterilized distilled water to the desired level using a micropipette. Ten microliter of the viral suspension was expelled into a haemocytometer chamber and was left undisturbed for 2 min. Counting of POBs was done under a phase contrast microscope (Nikon Eclipse 600) using doubled ruled Neubauer Haemocytometer (Weber scientific) with a 0.1 mm depth. Ten replicated counts were made systematically across the grid after appropriate settings for each sample. The quantified stock suspensions were stored at 4°C for further use.
Optimization Procedure
All experiments were conducted at 26±2°C temperature under 12:12 h dark: Light photoperiod. For viral inoculation a modified diet, without formaldehyde was prepared. A cube of diet, calculated to be consumed in 24 h was contaminated with a specific amount of virus. Insects were placed individually in 50 mL sterilized plastic containers and a single virus-contaminated diet disc was added for 24 h. The larvae were then transferred to clean uncontaminated diet free of formaldehyde and incubated for the required period. Larvae were monitored at 24 h interval for mortality due to HaNPV infection until pupation. Virus-infected cadavers were collected and frozen immediately. The samples were then homogenized with pre-cooled pestle and mortars and then transferred to pre sterilized vials and the final volume adjusted to 10 mL with distilled water. The numbers of Polyhedral Occlusion Bodies (POBs) were assessed for each replicate for all treatments and means were worked out.
The percent harvest was calculated as the ratio between total no of larvae harvested with typical NPV infection in each group/total no of larvae inoculated. The total yield was calculated as POB yield/larvae x total no of larvae harvested.
Optimum Inoculum Dose
To determine the optimum dosage of inoculum for virus production, 5, 7 and 9 day-old larvae were individually inoculated with different doses: 1x102, 1x104, 1x106 and 1x107 POBs per larvae. Twenty larvae that completely consumed diet disc were selected in each group and reared on uncontaminated diet. Larval mortality was recorded every 24 h and larvae found dead or moribund due to polyhedrosis were harvested individually up to 7 days after incubation. The harvested larvae were stored individually in microfuge tubes at -20°C. The experiment was replicated four times and the mean number of POBs per larva, % larval harvest and total yield of POBs for each treatment were calculated.
Optimum Larval Age for Inoculation
Four replicates of 5, 7, 8, 9, 10 and 11 day-old larvae were individually allowed to feed on artificial diet contaminated with the optimized inoculum dose (1x104 POBs/insect). Within each age group larval weights were restricted to as uniform as possible. Larval mortality was recorded every 24 h. Larvae attaining the pupal stage were reared until eclosion. It was assumed that emerging adults did not contain POBs, so they were discarded, whereas non-emerging pupae were examined microscopically for the presence of POBs. All the dead larvae and pupae of each age group containing POBs were collected and the mean number of POBs per insect as well as the total number of POBs for each treatment was calculated.
Optimum Incubation Period
To optimize the incubation period, seven day- old larvae were individually inoculated with a virus dose of 1x104 POBs/larva (optimized inoculum dose). They were allowed to feed and develop as in the previous experiments. After each of 5, 7 and 9 days of incubation, over 15 intact larvae which represented both moribund and recently dead specimen were harvested at specific time period and stored at -20°C. The larvae were later macerated and number of POBs per larva was determined. Pupae and ruptured/disintegrated larvae were discarded. To determine the percent harvest 100 larvae were inoculated individually for each period of incubation and harvest potential was assessed on each sampling day.
Optimum Physical Status of Harvested Larvae
To compare the production potential depending on the physical status of harvested larvae, 7-8 days old larvae of H. armigera were individually inoculated with an inoculum dose of 1x104 POBs/larva. Ten individuals from each group of larvae namely living infected, moribund and dead larvae were harvested on 7th day after incubation. Thereafter, the mean number of POBs per larva was determined as in the earlier experiments. The experiment was replicated 5 times and each replicate consisted of 50 individuals. For enumeration of bacterial contamination the viral suspension were serially diluted from 10 to 106 fold using sterile micro pipettes. The Petri dishes containing nutrient agar medium were marked into six equal segments for each dilution. Two drops of 10 μL of suspension of different dilutions were then poured on specific areas marked for each dilution. The plates were then left for about 25 min as such, thereafter incubated at 35±2°C in incubator overnight. Five replicates of each sample were maintained and Colony Forming Unit (cfu) in each segment were counted in colony counter twice every 24 h. The total bacterial population/larvae were calculated as
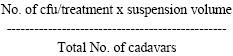 |
Statistical Analysis
The data recorded were statistically analysed by one way ANOVA (SPSS 10.0 Version) and means were compared with Tukey HSD a test (p<0.05).
RESULTS
Optimum Inoculum Dose
The results of testing different HaNPV inoculum doses on progeny virus produced in 5, 7 and 9 days old larvae revealed that the mean POBs yield per harvested larva followed an increasing trend initially and peaked at an inoculum dose of 1x104 POBs/larva in all the ages tested (Fig. 1). However, the increase of the inoculum dose beyond this range led to significant decrease in percentage of intact larvae harvested.
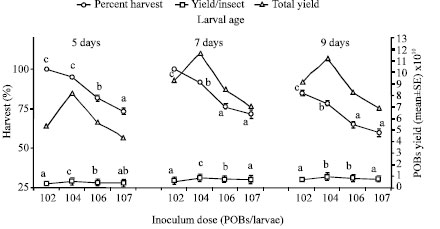 | |
| Fig. 1: | Production of HaNPV in 5, 7 and 9 days old larvae at different inoculum doses of virus *Vertical bars represent standard error of means, *Mean values followed by a common letter in respective age do not differ significantly; One way ANOVA, Tukey HSDa (p<0.05). The data were analyzed individually for each age but presented in a combined figure |
It was found that a virus inoculum dose of 1x104 POBs/larva resulted in the significant higher production of virus over all the other dosage tested with a respective yield of 0.54x1010, 0.82x1010 and 0.94x1010 OBs/insect in 5 (p<0.05, F = 14.92), 7 (p<0.05, F = 19.15) and 9 (p<0.05, F = 20.58) days old larvae.
Optimum Larval Age
When the larvae of different ages were inoculated with the optimized inoculum dose (1x104 POBs/larva), it was found that virus yield per insect increased with an increase in age of larvae tested (Fig. 2). In all the younger stages of larvae inoculated viz., 5, 7 and 8 days old, the mean virus yield/larva remained low in comparison to older larvae, the difference being significant (p<0.05, F = 46.23). However, in the older stages of larvae i.e., 9, 10 and 11 days, a significant decrease in per cent larval harvest was noticed (p<0.05, F = 11.13). Among the various ages of larvae inoculated, 11 days old larvae gave a maximum virus yield (1.01x1010 POBs/insect). However, at this stage only 65% of larvae could be harvested. The highest total yield of virus was recorded in 7 days old larvae. Nevertheless, the virus yield/insect as well as larval harvest at this age of inoculated larvae was at par with 8 days old larvae.
Optimum Incubation Period
No doubt, the increase in incubation period resulted in significant increase in yield of POBs/insect up to 7 days but it simultaneously led to significant reduction in larval harvest (p<0.05, F= 243.92) (Fig. 3). While cent percent larval harvest was observed in the larvae incubated for 3 and 5 days, it was highly reduced to as low as 45% in the larvae incubated for 9 days. Seven days post incubation led to desirable larval harvest (88.33%) coupled with a maximum POBs yield per larvae (0.84x1010).
Optimum Physical status of Harvested Larvae
While comparing the HaNPV productivity on the physical status of harvested larvae (Table 1), it was found that all the stages yielded different number of POBs per larva, the differences being significant (p<0.05, F = 9.75). The maximum POBs yield (8.74±0.21x109) per larva was obtained by harvesting the dead larvae, followed by 8.30±0.30x109 POBs/larva in moribund larvae and 7.34±0.14x109 POBs/larva in infected healthy larvae.
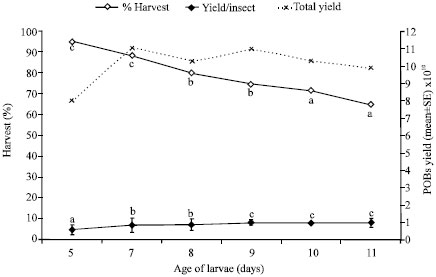 | |
| Fig. 2: | Production of HaNPV from larvae of different ages inoculated with optimized inoculum dose Vertical bars represent standard error of means, Mean values followed by a common letter in a line do not differ significantly; One way ANOVA,Tukey HSDa (p<0.05) |
 | |
| Fig. 3: | Production of HaNPV in 7 days old larvae fed with optimized viral dose (1x10 POBs/larva) and subjected to varied post incubation periods. Vertical bars represent standard error of means, Mean values followed by a common letter in a line do not differ significantly; One way ANOVA,Tukey' HSD (p<0.05) |
| Table 1: | Comparison of HaNPV productivity depending on physical status of harvested larvae |
 | |
| *Mean ±SE with in a column superscripted by different letter(s) are significantly different (p<0.05, Tukey HSDa) **Calculated by averaging 5 replicates of 10 infected larvae | |
However, harvesting the dead larvae was complicated by the disintegration of larval cuticle, which results in loss of POBs coupled with high microbial contamination (Table 1). The minimum bacterial count was observed in infected healthy larvae.
DISCUSSION
Mass production of the virus at reasonable costs is an important factor in its development into a marketable product. In vitro systems are the preferred option for recombinant baculovirus production but the techniques have not yet been sufficiently developed for large-scale production of either recombinants or natural isolates. The production in cell culture can also lead to reduction of biological activity of insect viruses coupled with variety of mutations or defective viral populations due to passage effect (Chakraborty and Reid, 1999; Chakraborty et al.,1999; Sun et al., 2005). Moreover, their cost of production is quite high in comparison to in vivo production which is highly practiced in developing nations for mass production of viruses. The cost of in vivo production can be further reduced if the production parameters are standardized with respect to inoculum dose, age of larvae, post incubation period and harvesting stage. An inoculum dose of 1x104 POBs/larva could be considered as optimum dose for inoculation, as it resulted in the highest virus yield/insect coupled with a desirable proportion of larval harvest in all the three ages of insect incubated for a period of seven days. Furthermore, at this dose the total yield of POBs harvested from treated larvae of different age in different set of experiments was highest. The increase in dose of inoculum from 1x102 to 1x104 raised the mean virus yields, but beyond this dose POBs yield per larvae started decreasing. At the lower inoculation doses, very few larvae were infected successfully while at the optimum dose most were infected. It seems probable that for a given size of larvae there is a threshold dose of NPV to overcome host resistance and initiate a successful infection. At lower dose e.g., at 1x102 POBs/larvae in this study, a high proportion of the larvae were able to restrict infection to the initial site of entry, the mid gut epithelium. The mechanism of resistance could be the rapid shedding of primary infected cells known to occur in other species (Engelhard and Volkman, 1995; Naryanan, 2004) and/or by an apoptotic response to cellular infection (Chejanosky and Gershburg,1995). Thus the larvae may prevent the development of secondary infection to other body tissues or even clear the infection entirely. Unless the NPV infection spread from the primary site in the mid gut epithelium to the other tissues of the body that are the main site of viral replication, there is no significant production of polyhedron. The present investigation indicated that inoculum dose retarded larval growth in a dose-dependent manner in each age group studied, a result, which is similar to that of Shapiro et al. (1981) and Grzywacz et al. (1998). At the highest dose of 1x107 POBs the yield per larvae was less than that obtained at the lower doses. This is related to the observation that at the high doses the growth of larvae was impaired. Similar severe growth impairment at higher doses has also been reported in Trichoplusia ni treated with Autographa californica NPV (Van Beek et al., 1990). Grzywacz et al. (1998) ascribed it to the combined effects of partial starvation resulting from the destruction of the midgut by a massive primary infection and the early establishment of secondary infection in other tissues through NPV virions passing directly into the haemocoel. This early establishment of secondary infection and resultant metabolic disruption prevents the larvae from growing to full size. Such higher doses of NPV may also have a serious negative effect on NPV productivity due to rapid mortality at higher doses. With very large oral doses of NPV, virions from the inoculum are able to pass across the midgut epithelia directly into haemocoel to initiate the secondary infection in other body tissues without the need for a primary replication cycle, thus speeding up the development of infection (Flipsen, 1995). The reduction in the production of POBs when the virus was used at a higher dose of 1x107 POBs/larva was in conformity with the report of Hedlund and Yendol (1974) in gypsy moth, Lymantria dispar larva. Shapiro et al. (1981) reported that if the inoculum dose is too high, the larval growth might be retarded, resulting in less than optimal yield. If the inoculum dose is too low, infection may not be uniform with in the host population, resulting in sub-optimal yield.
The virus yield/larva remained quite low in all the earlier stages of inoculated larvae i.e. 5, 7 and 8 days old larvae as compared to older larvae. However in the older stages of the larvae significant decrease in per cent larval harvest was noticed when compared to younger ones. The overall total yield of POBs is a multiple of virus yield/insect and number of intact larvae harvested. Although, the highest total yield of virus was recorded in 7 days old larvae, but the virus yield/insect as well as larval harvest at this age of inoculated larvae was at par with 8 days old larvae, therefore, the larvae of both these ages can be used for maximizing the total POBs yield. The increase of POBs yield/larva with the larval age was in conformity with the findings of Monobrullah and Nagata (2000), wherein maximum yield was obtained in 4th instar larvae, which was significantly higher than the yield in 1st instar. With increase in age the decrease in larval harvest after inoculation with optimized inoculum dose (1x104 POBs/larva) is due to the fact that old larvae are more resistant and complete pupation earlier. Traditionally this development of resistance to NPV infection has been attributed to increase in larval age or biomass (Briese, 1986), but recently stronger evidences were put forth wherein active sloughing of the mid gut cells was explained as defensive phenomenon (Washburn et al., 1996). Nevertheless, these viruses have evolved methods to bypass the defence phenomenon of apoptosis by directly blocking this response with the possession of P35 and Iap genes, thereby monitoring their own survival by suppressing apoptosis of host cells (Narayanan, 2004). The decrease in larval harvest in old larvae is attributed to maturation immunity (Whitlock, 1977). In healthy and old larvae infection of midgut and goblet cells and their replacement by differentiating regenerative cells is a natural process and may be another means for limiting infection. The observed increase in size of larvae, with low and intermediate doses may be attributed to the presence of egt gene in the baculoviruses (O’Reilly and Miller, 1989).
The virus yield per insect increased with the increase in incubation period up to 7 days beyond which a gradual decrease was observed. It can be attributed to the fact that in the infection cycle, the mean POBs yield per harvested larvae fell because the heavily infected larvae had dried earlier. On the other hand, the proportion of infected intact larvae for harvesting fell sharply from 100 per cent (3 and 5 days) to as low as 45%. The maximum virus yield/insect (8.65x109) coupled with highest total POBs yield (7.1x1010) could be achieved after 7 days of incubation because the desirable degree of percentage of larvae harvested (83.33%) decreased after 7 days of incubation. However, incubation beyond 7 days led to sharp decline in percent larvae harvested thereby leading to lower total POBs yield. The insect harvested at 3 and 5 days of incubation yielded low POBs along with the reduction of size of polyhedra. This period actually correspond to the primary infection phase in the midgut wherein virus replicate before producing progeny virions which subsequently spread in to body for secondary infection in various tissues. The optimum harvesting time found here was in conformity with the results of Bell (1991) who found optimum production of Helicoverpa armigera NPV in Heliothis zea 7 days after infection and very similar to that found in the other lepidopteran insects in which NPV production has been studied (Grzywacz et al., 1998; Kumar et al., 2005).
The maximum POBs yield per larva was obtained by harvesting the dead larvae. However, harvesting dead larvae was very difficult and there was high microbial contamination. Thus harvesting moribund larvae is the best option for optimum production of HaNPV. The results on optimum physical state of larvae were in conformity with the results of Monobrullah and Nagata (2000) in case of Spodoptera litura. However, Jones et al. (1998) found infected healthy larvae of H. armigera as optimum stage for production of maximum uncontaminated POBs. The improved productivity in term of POBs yield/insect in the current investigations may be attributed to the strain of virus used as the selection of improved strain of the virus lead to enhancement of productivity in in vivo systems (Grzywacz et al., 1998). However, further work is required to ascertain the exact reason for the improved productivity.
The result of present study clearly indicated that adjusting the inoculum dose and harvesting time had significant impact on NPV productivity. Correct selection of larval age for inoculation and physical status of larvae to be harvested were also found as crucial factors in maximizing the productivity, avoiding the losses from death and disintegration of larvae and minimizing the bacterial contaminations.
ACKNOWLEDGMENTS
We thank Dr. R.J. Rabindra, Director, PDBC, ICAR, Bangalore for helpful suggestions during Project Review Workshops for carrying out this research work. Thanks are also due to Dr. Phyllis Weintraub, Gilat Research Center and Nor Chejanovsky, Institute of Plant Protection, Volcani Center, ARO, Israel for their helpful comments on an earlier version of the manuscript. Laboratory assistance by Mr. Kamlesh Bali and Raghubir Lal is also duly acknowledged. This study was funded by the National Agriculture Technology Project, ICAR, India (CGP-II-108).
REFERENCES
- Bell, M.R., 1991. In vivo production of nuclear polyhedrosis virus utilizing tobacco budworm and a multicellular larval rearing container. J. Entomol. Sci., 26: 69-75.
Direct Link - Chakraborty, S. and S. Reid, 1999. Serial passage of Helicoverpa armigera nucleopolyhedrovirus in H zea cell cultures. J. Invertebr. Pathol., 73: 303-308.
Direct Link - Chakraborty, S., C. Monsour, R. Teakle and S. Reid, 1999. Yield, biological activity and field performance of a wild type Helicoverpa armigera nucleopolyhedrovirus production in H zea cell culture. J. Invertebr. Pathol., 73: 199-205.
Direct Link - Chejanovsky, N. and E. Gershburg, 1995. The wild type Autographa californica nuclear polyhedrosis virus induced apoptosis of Spodoptera littoralis. Virology, 209: 519-525.
Direct Link - Cherry, A.J., M.A. Parnell, D. Grzywacz and K.A. Jones, 1997. The optimization of in vivo nuclear polyhedrosis virus production in Spodoptera exempta (Walker) and Spodoptera exigua (Hubner). J. Invertbr. Pathol., 70: 50-58.
Direct Link - Cherry, A.J., R.J. Rabindra, M.A. Parnell, N. Geetha, J.S. Kennedy and D. Grazywacz, 2000. Field evaluation of Helicoverpa armigera nucleopolyhedrovirus formulation for control of the chickpea pod borer, H. armigera (Hubn.) on chickpea (Cicer arietinum var Shoba) in Southern India. Crop Prot., 19: 51-60.
Direct Link - Engelhard, E.K. and L.E. Volkman, 1995. Developmental resistance in fourth instar Trichoplusia ni orally inoculated with Autographa californica nuclear polyhedrosis virus. Virology, 209: 384-389.
Direct Link - Grazywacz, D., D.J. McKinley, G. Moawad and A. Cherry, 1998. The in vivo production of Spodoptera littoralis nuclear polyhedrosis virus. J. Virol. Methods, 71: 115-122.
Direct Link - Inceoglu, A.B., S.G. Kamita, A.C. Hinton, Q. Huang, T.F. Severson, K. Kang and B.D. Hammock, 2001. Recombinant baculoviruses for insect control. Pest Manage. Sci., 57: 981-987.
Direct Link - Jones, K.A, B. Zelazny, U. Ketunuti, A. Cherry and D. Grzywacz, 1998. A Survey of Virus Control of Insect-pests in South-east Asia and the Western Pacific. In: Insect Virus and Pest Management, Hunter-Fujita, F.R.H., P.F. Entwistle, M.F. Evans and N.E. Crook (Eds.). John Wiley and Sons, NY., USA., pp: 244-248.
- Kalia, V., S. Chaudhari and G.T. Gujar, 2001. Optimization of Production of nucleopolyhedrovirus of Helicoverpa armigera throughout larval stages. Phytoparasitica, 29: 23-28.
Direct Link - Kumar, C.M., N. Sathiah and R.J. Rabindra, 2005. Optimizing the time of harvest of nucleopolyhedrovirus infected Spodoptera litura (Fabricius) larvae under in vivo production system. Curr. Sci., 88: 1682-1684.
Direct Link - Monobrullah, M.D. and M. Nagata, 2000. Optimization of Spodoptera litura (Fab.) nucleopolyhedrovirus production in homologous host larvae. Insect Sci. Applied, 20: 157-165.
Direct Link - Narayanan, K., 2004. Insect defence: Its impact on microbial control of insect pests. Curr. Sci., 86: 800-814.
Direct Link - O`Reilly, D.R. and L.K. Miller, 1991. Improvement of baculovirus pesticide by deletion of EGT gene. Biotechnology, 9: 1086-1089.
Direct Link - Rabindra, R.J., N. Sathiah, K.P. Jayanth and R.K. Gupta, 2003. Commercial Scale Production and Formulation of Nucleopolyhedroviruses of Helicoverpa armigera and Spodoptera litura. In: Recent Advances in Integrated Pest Management, Singh, A., N. Bombawale and T.P. Trivedi (Eds.). NCIPM, New Delhi, pp: 32-39.
- Subramanian, S., G. Santharam, N. Sathiah, J.S. Kennedy and R.J. Rabindra, 2006. Influence of incubation temperature on productivity and quality of Spodoptera litura nucleopolyhedrovirus. Biol. Control, 37: 367-374.
Direct Link - Sun, X., X. Sun, B. Bai, W.V.D. Werf, J.M. Vlak and Z. Hu, 2005. Production of polyhedral inclusion bodies from Helicoverpa armigera larvae infected with wild type and recombinant HaSNPV. Biocontrol. Sci. Technol., 15: 353-366.
Direct Link - Van-Beek, N.A.M, P.H Flore, H.A. Wood and P.R. Hughes, 1990. Rate of increase of Autographa californica nuclear polyhedrosis virus in Trichoplusia ni larvae determined by DNA: DNA hybridization. J. Invertebr. Pathol., 55: 85-92.
Direct Link