
Donald D. Ourth
Department of Biology, The University of Memphis, Memphis, Tennessee 38152, US
Michael S. Parker
Department of Biology, The University of Memphis, Memphis, Tennessee 38152, US
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 2 | Page No.: 95-101







ABSTRACT
The antibacterial response of tobacco budworm (Heliothis virescens) larvae vaccinated with the bacterium Enterobacter cloacae peaked at 6-30 h following vaccination was significantly higher than the antibacterial response of unvaccinated larvae. The effect of immune hemolymph from H. virescens larvae was determined by the growth of Escherichia coli (strain K12 D31) on tryptic soy agar mixed with the immune hemolymph. The range of increase in bactericidal activity seen during this time period was from 49.1 to 71.1%. The greatest bactericidal activity (up to 71.1%) was observed from 12-20 h following the initial infection. A second vaccination (reinfection) with E. cloacae at 48 h following the initial vaccination did not result in any long-term potentiation of the larval antibacterial response, although it did boost short-term bactericidal activity by 3.1% after 12 h. This indicates that an anamnestic (memory) response is likely not elicited. The data demonstrated the time sequence in the antibacterial response of H. virescens larvae.
PDF Abstract XML References
How to cite this article
Donald D. Ourth and Michael S. Parker, 2006. Kinetics of the Antibacterial Response by Heliothis virescens Larvae. Journal of Entomology, 3: 95-101.
DOI: 10.3923/je.2006.95.101
URL: https://scialert.net/abstract/?doi=je.2006.95.101
DOI: 10.3923/je.2006.95.101
URL: https://scialert.net/abstract/?doi=je.2006.95.101
INTRODUCTION
Many insects have an important effect on agriculture, including the lepidopteran Heliothis virescens, the tobacco budworm (TBW) moth. The insect is a pest on many important crops, including cotton, in its larval stage.The humoral immune system of insects consists mainly of cecropins, attacins and lysozyme (Boman and Hultmark, 1987). Insects produce these inducible antibacterial factors in their hemolymph in response to injection of pathogenic bacteria. Cecropins (Mr 4,000-7,000; initially isolated from Hyalophora cecropia pupae) and attacins (Mr 20,000-23,000) have been the best-defined antibacterial insect proteins. Cecropins, cecropin-like peptides and lysozyme have been found throughout the Lepidopteran order, including H. virescens (Boman and Hultmark, 1987; Lockey and Ourth, 1996a, b; Morishima et al., 1990; Ourth et al., 1994; Qu et al., 1982).The purpose of this study was to determine when the H. virescens bactericidal response first began, when it peaked and how long it lasted. We wanted to elucidate the kinetics of the antibacterial response of TBW larvae over a period of 120 h following vaccination with Enterobacter cloacae and also to determine if an anamnestic (memory) response is possibly elicited after a second vaccination. The kinetics of the antibacterial defensive response of TBW larvae has not been previously determined.
MATERIALS AND METHODS
Tobacco budworm eggs were obtained from the Bioenvironmental Insect Control Laboratory, US Department of Agriculture, Stoneville, MS. The eggs hatched after 3 days. The larvae were reared individually in 30 mL plastic cups with lids. Each cup contained approximately 10 mL of diet media (Ourth, 1988; Raulston and King, 1984). The larvae were maintained in a natural photoperiod, at a temperature of 23-25°C, until reaching the fifth instar stage. At this time, the larvae averaged 2 cm in length. The larvae were then injected with 106 bacteria/larva of Enterobacter cloacae (ATCC #23355) using a microinjector. The larvae were divided into 57 groups of five larvae. Three groups of five larvae were then sacrificed at each of 19 time points ranging from 0 to 120 h following the initial bacterial vaccination. The larvae were anesthetized at -4°C for 10 min. Capillary tubes were then used to collect the hemolymph. The average yield of hemolymph was approximately 100-150 μL/larva. The hemolymph was pooled into Eppendorf microcentrifuge tubes containing the anti-melanizing agent 1-phenyl-2-thiourea (PTU). The collected hemolymph was stored at -80°C. An equal number of unvaccinated larvae was sacrificed at the same time points under identical conditions. These larvae had not been vaccinated with E. cloacae. At the 48 h time point, one group out of three groups of each remaining set of initially vaccinated larvae was reinfected with an additional 106 E. cloacae in order to determine whether a second vaccination would result in an increased antibacterial response. This could indicate the possibility of an anamnestic (memory) response in the antibacterial defense of these insect larvae. The reinfected larvae were then sacrificed at the same time points as the larvae which were vaccinated only once. Unvaccinated larvae, which received only a saline injection, were also included as controls.
A bactericidal assay study was done to determine the percent bactericidal activity of the hemolymph samples of the 57 groups of larvae. For each time point, 25 μL of pooled hemolymph from each group of five larvae was mixed with an equal volume of physiological saline solution containing Escherichia coli (K12 D31 strain) bacteria. The concentration of E. coli was approximately 4000-6000 bacteria/mL. The mixture of hemolymph and E. coli in physiological saline solution was incubated for 90 min at 30°C. It was then mixed with 10 mL of molten tryptic soy agar in a standard Petri dish. The plates were incubated at 37°C for 24 h. The number of E. coli colonies growing on each plate was then counted. A proportional equation was then used to calculate the percent bactericidal activity of the hemolymph samples (Ourth and Wilson, 1981).
The percentage of bactericidal activity for each time point was calculated by comparing the number of bacterial colonies growing on plates containing hemolymph from larvae vaccinated with E. cloacae to the number of colonies growing on plates with hemolymph from unvaccinated larvae (saline controls) bled at the same time point. The same procedure was used for the hemolymph collected from twice-vaccinated larvae. The differences in the percentages of bactericidal activity between the vaccinated and unvaccinated larvae, between the once-vaccinated and twice-vaccinated larvae and between twice-vaccinated and unvaccinated larvae were determined by calculating a comparison of the two groups in each case. In the two cases where the unvaccinated larvae were compared with the other two treatments, the number of bacterial colonies growing on the plates containing hemolymph from unvaccinated larvae was taken as the base value. The number of colonies growing on the plates containing hemolymph from the once-vaccinated or twice-vaccinated larvae was then proportionally compared to this base value. When comparing once-vaccinated and twice-vaccinated larvae, the number of bacterial colonies growing on the plates containing hemolymph from once-vaccinated larvae was taken as the base value. The number of colonies growing on the plates containing hemolymph from twice-vaccinated larvae was then obtained by subtraction.
RESULTS
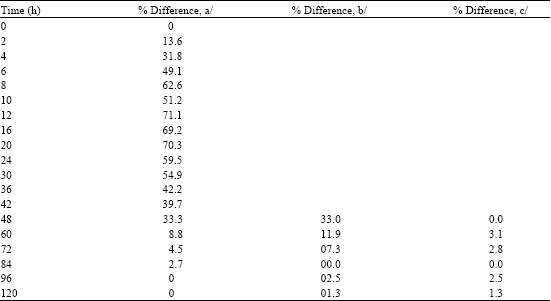
Hemolymph from vaccinated larvae contained much greater bactericidal activity than hemolymph from unvaccinated larvae (Fig. 1, 2 and Table 1). The bactericidal activity of vaccinated larvae was significantly greater than that of the unvaccinated larvae at all time points ranging from 2-48 h post-infection (range of increase 14-71%). The number of E. coli bacteria killed correlated with the peak bactericidal activity found at 12-20 h. Starting at 60 h, antibacterial activity remained at approximately the same level as the response of the unvaccinated larvae (Fig. 1, 2 and Table 1). The greatest bactericidal activity of 71.1 and 70.3% was seen at 12 h and 20 h, respectively, after the initial vaccination. The percentages of bactericidal activity (increase of 3.1% at 60 h and increase of 2.8% at 72 h) of the twice-vaccinated larvae were greater than those of the larvae that were vaccinated only once, likely indicating no anamnestic (memory) response. Starting at the 84 h time point, the bactericidal activity of the twice-vaccinated larvae reverted back to approximately the same levels as the bactericidal activity of the once-vaccinated groups (Fig. 1, 2 and Table 1).
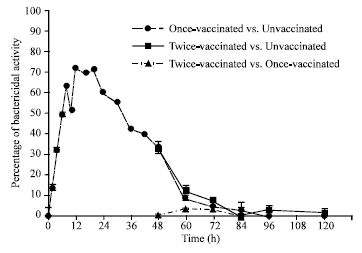 | |
| Fig. 1: | Bactericidal assay in Heliothis virescens larvae |
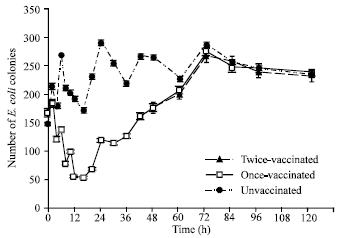 | |
| Fig. 2: | Effect of immune hemolymph from Heliothis virescens larvae on bacterial growth |
| Table 1: | Bactericidal activity of Heliothis virescens larvae | ||||||
 | |||||||
| |||||||
DISCUSSION
The main purpose of this study was to determine how quickly a lepidopteran larva, Heliothis virescens, can respond to an initial bacterial infection and when the peak in bactericidal activity occurs in relation to time after the initial infection. At the same time, an attempt was made to determine whether an insect larva has the immune capacity for an anamnestic (memory) response to a previous infection.
A comparison of the kinetics in vaccinated versus unvaccinated larvae showed an increase of bactericidal activity in larvae vaccinated with E. cloacae. The effect of immune hemolymph from H. virescens larvae was determined by the growth of E. coli on tryptic soy agar mixed with the immune hemolymph (Fig. 2). This is the first study of the kinetics of the bactericidal response over an extended period of time in an insect. At 2 h post-vaccination, the larvae were showing a nearly 14% increase in bactericidal activity, indicating that they were responding rapidly to the bacterial challenge (Fig. 1, 2 and Table 1). Hoffmann et al. (1993) found that the transcription rate of several antibacterial protein RNAs in Drosophila increased over a period varying between 6 h and 24 h, depending on the gene and then either stopped or leveled off. Morishima et al. (1995) found by Northern blot analysis that cecropin A and lysozyme mRNAs were both first detected in the larval fat body of the silkworm (Bombyx mori) one hour after injection of soluble peptidoglycan. We first detected bactericidal activity in H. virescens at 2 h following bacterial vaccination indicating transcription of mRNA. Our data corroborates their findings in that a definite increase in bactericidal activity and protein synthesis was seen very early at 2 h in the H. virescens immune response (Fig. 1, 2 and Table 1).
The H. virescens bactericidal activity then increased and remained elevated for 48 h following the initial vaccination with live bacteria. The greatest increases (49.1-71.1%) in bactericidal activity occured in hemolymph samples collected at 6-30 h after the initial vaccination by comparison with the unvaccinated controls. The greatest peaks in bactericidal activity occurred at 12 h (71.1%) and at 20 h (70.3%) (Fig. 1, 2 and Table 1).
Ourth and Smalley (1980) found that peak bactericidal activity, using a disk diffusion assay with hemolymph on inoculated agar, occurred at 24 h following vaccination of the adult cotton boll weevil (Anthonomus grandis) with heat-killed Serratia marcescens. Bactericidal activity of the boll weevils was nearly as great at 12 h as at 24 h, but no bactericidal activity was detected at 36 h post-vaccination.
Briggs (1958) initially found bactericidal activity present in hemolymph of 11 species of lepidopteran larvae. Live attenuated bacteria were used to vaccinate the 11 different species of lepidopteran larvae. Their antibacterial responses were followed from 8 h to 15 days post-vaccination (Briggs, 1958). All of the 11 species combined demonstrated an 8-24 h range of antibacterial activity. Stephens (1962) vaccinated wax moth (Galleria mellonella) larvae with dead Pseudomonas aeruginosa. Maximum bactericidal activity was seen between 16 and 24 h after vaccination. Bactericidal activity correlated well with protective immunity. The interval between 4 and 72 h was followed. No bactericidal activity was detected with the assay at 4 h and the bactericidal activity was in decline at 48 h with about half the bactericidal activity remaining when compared with the bactericidal peak time from 16 to 24 h. Very little bactericidal activity remained at 72 h following vaccination and the bactericidal activity had nearly returned to pretreatment levels. These results reflect the immune response to a dead bacterial preparation rather than the immune response to a live bacterial vaccine as was done in our experiments with H. virescens. Our results with H. virescens also corroborate those of Briggs (1958) and Stephens (1962) in that no bactericidal activity was detected prior to vaccination (Fig. 1, 2 and Table 1). The time occurrence of the H. virescens bactericidal response indicates the rapidity of the defensive response following an initial Gram-negative bacterial infection or live vaccination as was done here.
Dunn (1990) found that antibacterial proteins were first seen at 6-8 h post-infection in Manduca sexta. Spence et al. (1992) found that all major inducible proteins were seen 24 h after infection in M. sexta. For many of these antibacterial proteins, the response was strongest at 48-72 h post-infection.
Sumida et al. (1992) immunized Bombyx mori (silkworm) larvae with formalin-treated E. coli K-12. Using a well diffusion inhibition assay, a peak of antibacterial activity was seen in 5th-instar larvae (day 3-4 of 5th instar) from 12-24 h post-immunization. Cecropin-like A and B activity bands were detected by acid gel electrophoresis. Ichimori et al. (1992) found that only live E. coli and not formalin-treated E. coli induced antibacterial activity in 5th-instar B. mori larvae (day 1 of 5th instar) after oral immunization. This antibacterial activity began 48 h post-immunization and lasted for 120 h post-immunization. They interpreted this to mean that multiplication of E. coli in the larval midgut was required for induction of an antibacterial response.
In the present study, a second vaccination at 48 h also showed a small increase of 3.1% in bactericidal activity when compared with the once-vaccinated larvae. Reinfection at 48 h of once-vaccinated larvae resulted in a bactericidal response that increased bactericidal activity by 3.1% at 60 h and 2.8% at 72 h. Although a small increase in bactericidal activity was seen following the second vaccination, this probably does not indicate an anamnestic or memory response. With the second vaccination, only a small increase in bactericidal activity was seen, indicating that the larvae had probably reached their maximum capacity for a bactericidal response. An anamnestic response was reported for the American cockroach, Periplaneta americana (Karp and Rheins, 1980). Cockroaches were given a primary injection of honeybee venom toxoid. Two weeks later, a second injection of honeybee toxoid was given and a secondary or anamnestic response was exhibited by the cockroaches.
We tried to determine if H. virescens could have a memory response to the first vaccination, perhaps represented by a large increase in bactericidal activity after a second injection of bacteria. Although there was a small percentage increase in bactericidal activity (3.1%) with the twice-vaccinated larvae, this very small increase probably does not represent an anamnestic (memory) response (Fig. 1, 2 and Table 1) when compared with the large increase in the memory immune response of a vertebrate. It is therefore our conclusion that lepidopteran larvae likely do not have memory capacity to a bacterial infection analogous to the vertebrate memory response. The data demonstrate the economics of innate immunity by larvae in that the data reflect the fitness costs and maximum survival from an infection under non-starvation conditions (Moret and Schmid-Hempel, 2000; Read and Allen, 2000).
ACKNOWLEDGEMENT
The authors thank Dr. Hans Boman, University of Stockholm, Sweden, for providing the Escherichia coli K12 D31 bacterial strain used in this study.
REFERENCES
- Moret, Y. and P. Schmid-Hempel, 2000. Survival for immunity: The price of immune system activation for bumblebee workers. Science, 290: 1166-1168.
CrossRefDirect Link - Morishima, I., S. Suginaka, T. Ueno and H. Hirano, 1990. Isolation and structure of cecropins, inducible antibacterial peptides, rom he silkworm, Bombyx mori. Comp. Biochem. Physiol., 95: 551-554.
Direct Link - Sumida, M., H. Ichimori, S. Johchi, A. Takaoka, T. Yuhki, H. Mori and F. Matsubara, 1992. Antibacterial activity inducible in the haemolymph of the silkworm, Bombyx mori, by injection of formalin-treated Escherichia coli K-12 during the fifth larval instar and pharate adult development. Comp. Biochem. Physiol., 101: 165-171.