
Norazizah Shafee
Department of Medical Microbiology, Faculty of Medicine, Universiti Malaya Medical Center, 50603 Kuala Lumpur, Malaysia
Sazaly AbuBakar
Department of Medical Microbiology, Faculty of Medicine, Universiti Malaya Medical Center, 50603 Kuala Lumpur, Malaysia
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 1 | Page No.: 89-94







ABSTRACT
Potential occurrence of dengue 2 virus (DENV-2)-induced apoptosis in mosquito larvae was investigated. Heads of Toxohrynchites splendens mosquito larvae were inoculated with DENV-2 inoculum at multiplicity of infection of 0.0005-0.001 plaque forming unit per cell. After 7 days of incubation, the larvae heads were severed and squashed onto slides. Following immunofluorescence staining with mouse polyclonal antibody against DENV-2, antigens were detected in 17.5% of the total cells counted. Counterstaining of the cells using colorimetric apoptotic staining method revealed that 21.7% of the antigen-positive cells were in apoptotic state. Statistical analyses of the data showed that the induction of apoptosis was directly correlated to the DENV-2 infection and not merely a metamorphic programmed cell death response.
PDF Abstract XML References
How to cite this article
Norazizah Shafee and Sazaly AbuBakar, 2006. Effects of Dengue 2 Virus Inoculation on Toxorhynchites splendens Larvae. Journal of Entomology, 3: 89-94.
DOI: 10.3923/je.2006.89.94
URL: https://scialert.net/abstract/?doi=je.2006.89.94
DOI: 10.3923/je.2006.89.94
URL: https://scialert.net/abstract/?doi=je.2006.89.94
INTRODUCTION
Mosquito-borne diseases contribute significantly to human health problems. Their incidence has increased significantly within the last decade particularly in the tropics and subtropics (Reiter, 2001; Malavige et al., 2004). Mosquito-borne diseases include Murray Valley encephalitis, St. Louis encephalitis, lymphatic filariasis, malaria, West Nile fever, yellow fever and dengue fever. Dengue fever is caused by dengue viruses that are transmitted by mosquitoes of the genus Aedes. Dengue virus is a positive single-stranded RNA virus first isolated by Sabin and Schlesinger (Sabin and Schlesinger, 1945). The virus belongs to the Flaviviridae family of the genus Flavivirus. There are four distinct serotypes of dengue virus, dengue 1, 2, 3 and 4. These viruses cause a spectrum of disease severity ranging from the mild dengue fever to the more serious dengue hemorrhagic fever or dengue shock syndrome.
Dengue viruses are maintained in a cycle that involves humans and Aedes aegypti. This domestic, day-biting mosquito prefers to feed on humans which lead to viral transmission to humans through bites of infected female mosquito. The adaptation of the virus to human hosts was suspected due to human settlement near forest where the virus was transmitted through peridomestic mosquitoes such as Aedes albopictus or the Stegomyia species (Rudnick, 1978). Human migratory activities might have facilitated the movement of virus into towns and cities where the virus was sporadically transmitted by the two species of mosquito vectors. Early documentations of Aedes aegypti species showed that the vector might have originated from Africa and introduced to the rest of the world through slave tradings (Anonymous, 2000). The species then became highly adapted to humans and urban environments and ultimately spread as urbanization expended.
Dengue virus has been reported to induce apoptosis in various cell lines (Thongtan et al., 2004; Lin et al., 2002; Su et al., 2001). However, no studies have reported the occurrence of apoptosis in mosquito cells infected with dengue virus. In the present study we investigated the occurrence of apoptosis in T. splendens mosquito larvae following intrathorachic inoculation of DENV-2.
MATERIALS AND METHODS
Virus Inoculum
Virus inoculum used in the study was prepared by infecting C6/36 cell cultures at 70-80% confluency with DENV-2 (New Guinea C strain) to give an estimated multiplicity of infection of about 0.01 plaque forming unit per cell. Once the infected cell cultures shown about 90% cytopathic effects, the infected cells were then frozen and stored at -70°C. The virus inoculum was then prepared by freeze-thawing the infected cell cultures and sedimenting the supernatant at 40 000 x g to remove the cell debris. The Cell culture supernatant obtained from the centrifugation was later used as the virus inoculum. Infectivity of the inoculum was determined by performing viral plaque assays using porcine spleen cells (Rao, 1976).
DENV-2 Inoculation
Intrathoracic inoculation of DENV-2 into T. splendens mosquito larvae was performed according to the method described by Rosen and Gubler (1974) with minor modifications. The method used compressed air through drawn out, calibrated glass microcapillary tubes to push liquid into the T. splendens larvae heads. DENV-2 inoculum at multiplicity of infection of 0.0005-0.001 (2-3 μL) was injected into heads of T. splendens larvae immobilized on ice for 5 min. Inoculated larvae were held at 24°C in a container half-filled with water and subjected to a 12 h daylight cycle. Throughout the incubation period, the predaceous larvae were fed with the smaller Aedes aegypti larvae. After 7 days, the larvae heads were severed using sterile scalpels and squashed onto glass slides precoated with 0.01% poly-L-lysine solution. Slides were fixed in 4% paraformaldehyde for 25 min, permeabilized in 0.5% Triton X-100 for 5 min, washed in PBS and finally in distilled water. Larvae injected with media alone were included as controls. All the procedures were performed on ice. The slides were then air-dried and kept at 4°C until needed.
Detection of Antigen-positive Cells
The presence of DENV-2 viral proteins was detected by direct fluorescence antibody technique using pooled mouse anti-DENV-2 hyperimmune sera at 1:40 dilution in 1 x PBS for 1 h at 37°C. Cells were later washed twice for 5 min each with 1x PBS followed by incubation with FITC (1:50)-conjugated anti mouse IgG (Sigma, USA) at 37°C for 1 h. Slides were then washed three times in 1 x PBS, once with distilled water and continued with colorimetric staining of apoptotic cells prior to viewing under a fluorescent microscope (Axiolab; Zeiss, Germany).
Detection of Apoptotic Cells
TdT FragELTM DNA Fragmentation Detection Kit (Calbiochem, USA) employing the TUNEL principle was used to stain apoptotic cells following protocols provided by the manufacturer. The system utilizes biotin-labeled deoxynucleotides and detected with streptavidin-horseradish peroxidase (SA-HRP) conjugate. Upon staining, cells were air-dried and observed under a light microscope (Telaval 31; Zeiss, Germany).
Cell Counting
The percentages of antigen-positive and apoptotic cells were determined as previously detailed (Shafee and AbuBakar, 2002). Statistical analyses using student's t-test and Pearson correlation were performed using the SPSS for Windows version 10.5A (SPSS Inc., USA).
RESULTS AND DISCUSSION
Mosquito inoculation is a widely used method for dengue virus isolation (Vaughn et al., 1997; Kuno et al., 1985; Gubler and Sather, 1988). Mosquito species commonly used for virus isolation includes A. aegypti, A. albopictus, T. amboinensis and T. splendens (Gubler, 1998). In the present investigation, effects of DENV-2 inoculation into heads of T. splendens mosquito larvae were investigated. Fluorescence microscopic analyses of stained T. splendens mosquito larvae head cells injected with DENV-2 inoculum showed the appearance of DENV-2 antigen-positive cells (Fig. 1a). This observation is in agreement with previous reports which indicated that dengue viruses replicate in most mosquito tissues, including the brain (Rosen and Gubler, 1974; Gubler, 1998).
Following staining of apoptotic cells by colorimetric detection method, a number of the antigen-positive cells were shown to be in apoptotic state (Fig. 1b). This observation appeared to be the first to show the occurrence of apoptosis in heads of T. splendens mosquito larvae inoculated with DENV-2. In vitro, however, sindbis virus, another mosquito-borne virus, was shown to induce low level of apoptosis in A. albopictus cell line. Karpf and Brown (1998) reported that the occurrence of apoptosis in Sindbis virus-infected A. albopictus cells did not correlate with a large induction of apoptosis. Another study showed that persistent Sindbis virus infection induces distinct cytological effects on A. albopictus cells including an increase in apoptosis (Karpf et al., 1997). Recently, Hopwood et al. (2001) reported that malarial parasites induced apoptosis in the ovaries of Anopheles stephensi mosquitoes and inhibit oogenesis in these mosquitoes. Another study reported the induction of apoptosis by Bacillus thuringiensis subsp. israelensis in polytene midgut cells of Culex pipiens larvae (Smouse and Nishiura, 1997).These studies which suggested that apoptosis do occur in infected mosquito larvae tissues are in agreement with data obtained from our present investigation.
To calculate the total number of cells present, the samples were counter stained with propidium iodide which stained up all the cells nuclei (Fig. 1c). Following determination of the number of apoptotic and antigen positive cells (Shafee and AbuBakar, 2002), it was found that 17.5% of the cells stained positive for DENV-2 antigens (Table 1). A significant number (21.7%) of the antigen-positive cells appeared to be apoptotic. This mean value was found to be significantly different (p<0.001, two-tailed student's t-test) than the mean value of samples injected with media alone which served as the controls. It was also noted that the increased percentage of apoptotic cells was directly correlated (r = 0.75, p<0.001, Pearson) with the number of antigen positive cells, suggesting that dengue virus infection could result in apoptotic cells in mosquito larvae head cells. Previous studies have shown that metamorphic programmed cell death occurred in some insects (Nishiura and Smouse, 2000; White et al., 1994). In the present investigation, results of the statistical analyses suggested that DENV-2 induced apoptosis in T. splendens larvae and the apoptosis was not due to metamorphic programmed cell death.
Present observation of DENV-2-induced apoptosis in T. splendens is an important finding which implies the mechanism of vector responses to dengue virus infections. In the present investigation T. splendens larvae were used. The use of this predacious Toxorhynchites mosquitoes is safe since it avoids the possibility of accidental transmission of the virus by escaped mosquitoes (Rosen and Gubler, 1974; Gubler and Sather, 1988).
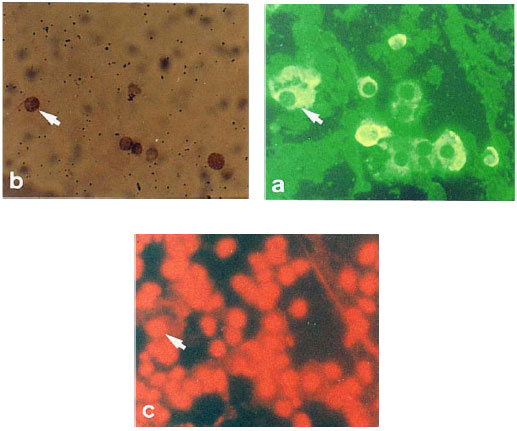 | |
| Fig. 1: | Detection of antigen-positive and apoptotic cells in T. splendens mosquito larvae heads injected with DENV-2 inoculum. Panels a, b and c show the same view-field observed using different filters. Arrows indicate a cell which expressed dengue viral antigens and also apoptotic. Detection of dengue virus antigens using mouse hyperimmune sera developed using anti-mouse FITC conjugated antibodies (a). Similar sample was stained with TdT FragEL DNA Fragmentation Detection Kit to detect apoptotic cells (b). Sample was then counter-stained with propidium iodide to stain all cells© |
| Table 1: | The influence of DENV-2 inoculation on T. splendens mosquito larvae. Harvested larvae head cells were stained for DENV-2 antigens, apoptotic cells and then counterstained with propidium iodide. The number of antigen-positive cells was determined and the percentage over the total number of cells was calculated. The percentage of antigen-positive cells which appeared apoptotic was determined by the number of apoptotic cells over the number of antigen positive cells. Results are presented as mean±SD. |
Since mosquitoes are the natural vector, it is probable that less selection of virus strains takes place than with suckling mouse or tissue culture inoculation. Further studies, nevertheless, is crucial to investigate such phenomenon in Aedes aegypti or Aedes albopictus mosquitoes, which constitute the main vector in dengue virus spread. In summary, results presented in this investigation provide an important insight into the mechanism of dengue virus replication in mosquito vectors.
ACKNOWLEDGEMENT
This research project was supported by IRPA Grants No. 06-02-03-0303 and 06-02-03-0618.
REFERENCES
- Gubler, D.J., 1998. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev., 11: 480-496.
PubMedDirect Link - Hopwood, J.A., A.M. Ahmed, A. Polwart, G.T. Williams and H. Hurd, 2001. Malaria-induced apoptosis in mosquito ovaries: A mechanism to control vector egg production. J. Exp. Biol., 204: 2773-2780.
Direct Link - Karpf, A.R., J.M. Blake and D.T. Brown, 1997. Characterization of the infection of Aedes albopictus cell clones by sindbis virus. Virus Res., 50: 1-13.
CrossRef - Kuno, G., D.J. Gubler and N.S. Santiago-de-Weil, 1985. Antigen capture ELISA for the identification of dengue viruses J. Virol. Methods, 12: 93-103.
Direct Link - Nishiura, J.T. and D. Smouse, 2000. Nuclear and cytoplasmic changes in the Culex pipiens (Diptera: Culicidae) alimentary canal during metamorphosis and their relationship to programmed cell death. Ann. Entomol. Soc. Am., 93: 282-290.
Direct Link - Reiter, P., 2001. Climate change and mosquito-borne disease. Environ. Health Perspect., 109: 141-161.
CrossRefDirect Link - Sabin, A.B. and R.W. Schlesinger, 1945. Production of immunity to dengue with virus modified by propagation in mice. Science, 101: 640-642.
PubMed - Su, H.L., Y.L. Lin, H.P. Yu, C.H. Tsao, L.K. Chen Y.T. Liu and C.L. Liao, 2001. The effect of human bcl-2 and bcl-X genes on dengue virus-induced apoptosis in cultured cells. Virology, 282: 141-153.
PubMed - Thongtan, T., S. Panyim and D.R. Smith, 2004. Apoptosis in dengue virus infected liver cell lines hepG2 and Hep3B. J. Med. Virol., 72: 436-444.
Direct Link - Vaughn, D.W., S. Green, S. Kalayanarooj, B.L. Innis and S. Nimmannitya et al.., 1997. Dengue in the early febrile phase: Viremia and antibody responses. J. Infect. Dis., 176: 322-330.
Direct Link - White, K., M.E. Grether, J.M. Abrams, L. Young, K. Farrell and H. Steller, 1994. Genetic control of programmed cell death in drosophila. Science, 264: 677-683.
Direct Link