
P. N. Deligeorgidis
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
L. Giakalis
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
G. Sidiropoulos
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
M. Vaiopoulou
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
G. Kaltsoudas
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
C. G. Ipsilandis
Institute of Western Macedonia, Branch of Florina, Department of Crop Production, Terma Kontopoulou, 53100 Florina, Greece
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 1 | Page No.: 61-69







ABSTRACT
The biological cycle, the duration of preoviposition-oviposition-postoviposition periods, female adults` longevity, reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) of Frankliniella occidentalis (Pergande) and Thrips tabaci Lindeman (Thysanoptera: Thripidae) were studied on cucumber at 15, 20, 25 and 30°C. The biological cycle of F. occidentalis ranged from 33.99 days at 15°C to 11.99 days at 30°C, while that of T. tabaci ranged from 32.76 to 11.78 days at respective temperatures. The duration of each developmental stage for both thrips species was reduced with temperature increase. Total female longevity of F. occidentalis decreased when temperature increased from 15 to 30°C, while the longevity of T. tabaci was lower and also decreased from 15 to 30°C. For F. occidentalis the maximum reproduction dynamic and reproduction rate was found at 20°C and minimum at 15°C. For T. tabaci the maximum reproduction dynamic was also found at 20°C and the optimum reproduction rate at 25°C, but without statistically significant difference when compared to reproduction rate at 20°C. The minimum values of T. tabaci were also found at 15°C. Cucumber found to be a suitable host plant for survival and reproduction of both thrips species. Temperature may affect thrips population, especially when measurements our out of normal temperature range.
PDF Abstract XML References
How to cite this article
P. N. Deligeorgidis, L. Giakalis, G. Sidiropoulos, M. Vaiopoulou, G. Kaltsoudas and C. G. Ipsilandis, 2006. Longevity and Reproduction of Frankliniella occidentalis and Thrips tabaci on Cucumber under Controlled Conditions. Journal of Entomology, 3: 61-69.
DOI: 10.3923/je.2006.61.69
URL: https://scialert.net/abstract/?doi=je.2006.61.69
DOI: 10.3923/je.2006.61.69
URL: https://scialert.net/abstract/?doi=je.2006.61.69
INTRODUCTION
Many Thysanoptera are phytophagous and considered economically important because they cause damages on cultivated plant species (Bailey, 1938, 1940; Watts, 1936). The polyphagous thrips species, Frankliniella occidentalis (Pergande) and Thrips tabaci Lindeman (Thysanoptera: Thripidae) attack numerous vegetables, ornamentals, cereals, fruit crops and cause serious damages (Wolfenbarger and Hibbis, 1958; Yokoyama, 1977; Anathakrishnan, 1984; Mustafa, 1986; Edelson et al., 1986; Shelton and North, 1987; Kendall and Capinera, 1987; Broadbent et al., 1987; Mantel and Van de Vrie, 1988; Lenteren and Woets, 1988; Terry and Degrandi-Hoffman, 1988). F. occidentalis and T. tabaci can also transmit the TSWV (Mantel and Van de Vrie, 1988; Robb and Parrella, 1987; Cho et al., 1987) General descriptive biology is well documented for many thrips species (Anathakrishnan, 1984; Lewis, 1973). However, comprehensive studies on the duration of preoviposition, oviposition, postoviposition periods, female longevity and reproduction of F. occidentalis and T. tabaci over a wide range of temperatures and particularly on cucumber, have not been conducted. Temperature-dependent growth is an important parameter for explaining fluctuations in population density (Logan et al., 1976; Gaum et al., 1994; Rijn et al., 1995; Deligeorgidis et al., 2005): the population peak of F. occidentalis and T. tabaci in cucumber and tomato crops (in glasshouse) was found between 20/5/2000 and 30/5/2000 (Deligeorgidis, 2005). Quantitative description on rates of development are often being used for predicting biological events in insects (Wagner et al., 1984). Temperature-dependent developmental rates and life-fertility budgets constructed at different temperatures provide quantitative parameters for describing and predicting thrips population dynamics (Gaum et al., 1994; Rijn et al., 1995; Deligeorgidis et al., 2005).
The objective of this study was to determine the biological cycle, the preoviposition-oviposition-postoviposition periods, the total female longevity, reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) of F. occidentalis and T. tabaci, on cucumber at four temperatures and provide quantitative parameters describing population fluctuation.
MATERIALS AND METHODS
In 2003, from April to June, the biological cycle, the duration of preoviposition-oviposition-postoviposition periods, female longevity, reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) of F. occidentalis and T. tabaci were studied on cucumber (Cucumis sativus L. F1 Kamaron) under controlled laboratory conditions: at four temperatures 15, 20, 25 and 30°C, 65±3% RH, 14 L: 10 D photoperiod and light intensity 9000 Lux. Thirty two cucumber plants planted in small pots (12x10 cm) were used for each thrips species (64 in total), 8 plants at each temperature, on which sprays with fungicides Ridomyl MZ 63.5 WP and Benlate 50 WP were applied at a rate 15 g and 6 g/10 L water, for protection of the plants from mycological diseases. Each cucumber plant was infested with one female adult (one day-old) of F. occidentalis. Similarly, the same process was followed with T. tabaci. Thrips were collected from rearing on cucumber kept in the laboratory. Each plant was covered with plastic (0.4 mm) cylindrical cage 11x40 cm. The openings on the top and the sides of these cages were covered with dense muslin (0.06 mm opening) for ventilation. The number of eggs oviposited by each female was measured by examining the tissues of each cucumber leaf under a stereoscope every day. The newly emerged larvae I, after being recorded on plants, were discarded every day. For the study of the biological cycle there were used 10 eggs from each pot at the four temperatures, for each thrips species. Each larva I individual, was placed into a plastic cylindrical cage 1.5x5 cm in which there was a piece of cucumber leaf 4 cm long by 1 cm wide, which was renewed every day until the development of adults, used to calculate the duration of immature stage total biological cycle. ANOVA across temperatures was conducted (8 replications/thrips species/temperature) and means were compared according to Tukey-Kramer multiple range test. Because the values (in days) of the duration of the immature stages were very low, a transformation was applied according to the formula (Snedecor and Cochran, 1980):
For the estimation of duration of preoviposition-oviposition-postoviposition periods, female longevity, reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) at each temperature, there were used the first 14 female adults (for both thrips species) that were developed, because this was the least number of individuals that completed full development from egg to adult stage, in order to meet ANOVA requirements.
RESULTS
Temperature affects significantly the duration of each stage and the length of these periods decreased as temperatures increased. The biological cycle of F. occidentalis ranged from 33.99 days at 15°C to 11.99 days at 30°C, while that of T. tabaci ranged from 32.76 to 11.78 days at respective temperatures (Table 1).
In Table 2, it is clearly shown that temperature importantly affects the duration of preoviposition-oviposition-postoviposition periods of F. occidentalis and T. tabaci. The length of these periods decreased as temperatures increased. The female adult longevity of both thrips species was closely related to temperature. Total female longevity of F. occidentalis ranged from 60.86 days at 15°C to 26.86 at 30°C, while the longevity of T. tabaci ranged from 54.12 days at 15°C to 20.99 days at 30°C. Preoviposition of F. occidentalis lasted more at 15°C (8.8 days) and at 20, 25 and 30°C was estimated 2.73, 2.41 and 2.33 days, respectively with no significant differences among them. The oviposition and postoviposition periods as well as female adult longevity of F. occidentalis were found to be significantly different across the four temperatures. Results of T. tabaci were alike. The preoviposition period at 20, 25 and 30°C found 2.60, 2.26 and 2.13 days, respectively but there were no significant differences among them. Only at 15°C (8.06 days) there was found a significant increase. The oviposition and postoviposition periods and female adult longevity of T. tabaci were found to be significantly different across temperatures.
In general, biological cycle of both thrips species and female longevity tend to increase at lower temperatures. These tensions are statistically significant and predictable by second-degree models (departure from linearity was significant) shown in Fig. 1. Total biological cycle (in days) of immature stages (BC) and Female Longevity (FL) for F. occidentalis (FO) and T. tabaci (TT) can be predicted with accuracy (p<0.001) by the following equations:
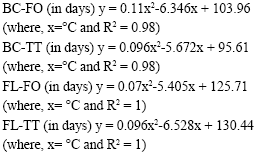 |
The statistical analysis of data of reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) for F. occidentalis showed significant differences among the four temperature treatments (Table 3). The highest reproduction dynamic (71.86 eggs/female) and reproduction rate (1.94 eggs/female/day) have been observed at 20°C, differing significantly in comparison to the rest of the temperatures. The temperature increase at 25 and 30°C reduced the reproduction dynamic and the reproduction rate. The values of both parameters were even lower at 15°C. Similarly, for T. tabaci there were significant differences across the four temperatures. The optimal temperature for reproduction dynamic (53.33 eggs/female) was found at 20°C and maximum reproduction rate (1.74 eggs/female/day) at 25°C, which was not significantly different from that at 20°C (1.72 eggs/female/day). The minimum reproduction dynamic and reproduction rate were observed at 15°C.
| Table 1: | Duration of immature stages and biological cycle (in days) of F. occidentalis and T. tabaci under controlled conditions, across temperatures |
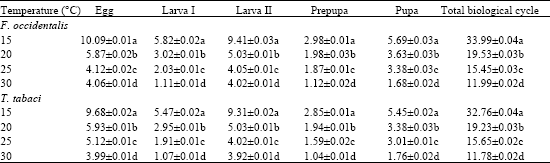 | |
| Means followed by the same letter within each column are not significantly different (p<0.05) according to Tukey-Kramer Multiple Range Test | |
| Table 2: | Means of pre-oviposition, oviposition, post-oviposition periods and total female adults longevity (in days) of F. occidentalis and T. tabaci under controlled conditions, across temperatures |
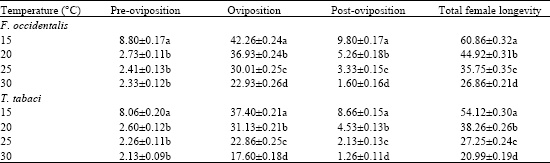 | |
| Means followed by the same letter within each column are not significantly different (p<0.05) according to Tukey-Kramer Multiple Range Test | |
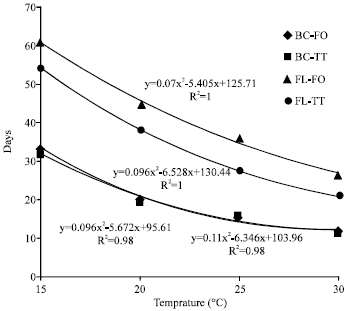 | |
| Fig. 1: | Total biological cycle (in days) of immature stages (BC) and Female Longevity (FL) for F. occidentalis (FO) and T. tabaci (TT) in relation to temperature (°C) |
| Table 3: | Reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) mean values of F. occidentalis and T. tabaci under controlled conditions, across temperatures |
 | |
| Means followed by the same letter within each column are not significantly different (p<0.05) according to Tukey-Kramer multiple range test | |
DISCUSSION
The data of this study indicated that F. occidentalis and T. tabaci may successfully survive and reproduce on the cucumber plant. The duration of immature stage cycle, the length of preoviposition-oviposition-postoviposition periods and thus the female adult longevity of these two species was decreasing as temperature was increasing. The reproduction dynamic and reproduction rate, also, of both thrips species have been influenced significantly by temperature. Second-degree models may predict with accuracy, female longevity and immature stage cycle in relation to temperature. These models, are similar to those found by Gaum et al. (1994) and Rijn Van et al. (1995). Differences between present findings and the studies of the above mentioned researchers are due to different cucumber varieties used, different age calculation that lead to lower developmental and ovipositional rates Gaum et al. (1994) and different number of entries (N). Also, there are slight differences between the model used in this study and the model proposed by Rijn Van et al. (1995) because developmental rate is more or less linearly depended to temperature between 12 and 28°C for F. occidentalis (between 17 and 28°C for T. tabaci (Edelson and Magaro, 1988)), but biological cycle and female longevity in this study showed a departure from linearity and temperatures used refer to a different interval (from 15 to 30°C) and are equally spaced (5°C step) in purpose, for best fitting a model. Calculations for female longevity were more or less independent from mortality, because of the procedure followed after the immature stage and female adults development.
For F. occidentalis, the length of preoviposition lasted for a longer period at the lower temperature of 15°C, delaying the maturing of the females for oviposition. On the contrary, the length of this period from 20 to 30°C was much shorter than at 15°C. As a consequence the maturation of the female for oviposition occurs much earlier from 20 to 30°C than at 15°C. The oviposition and postoviposition periods of the females at four temperatures exhibited significant differences. As temperature increases the duration of oviposition and postoviposition was significantly reduced. The longer duration (in days) of these periods was recorded at 15°C as it was also observed in the preoviposition period, while the shorter found at 30°C. Thus, females’ longevity of F. occidentalis was decreasing as the temperature was increasing from 15 to 30°C. Therefore, it could be concluded that female adults tends to live longer at the lower temperatures and less in the higher ones.
The reproduction of F. occidentalis on cucumber plants has been proved to be closely depended on temperature. Females reared at 20°C reproduced at a significantly higher rate than those reared at 15, 25 and 30°C. Thus, the optimal temperature for reproduction dynamic (eggs/female) and reproduction rate (eggs/female/day) is considered to be 20°C. Bryan and Smith (1956) showed that the duration of preoviposition of F. occidentalis was 3 days, postoviposition 30 days, females’ longevity 90 days and reproduction 40-65 eggs/female on the radish/bean pods at 15°C. Bryan and Smith (1956) also reported that the reproduction dynamic (eggs/female) at 20°C was the same with that at 15°C, while at 26.7°C was 20-40 eggs/female and 0.66-1.63 eggs/female/day. Lublinkhof and Foster (1977) reported a preoviposition period of 10.44, 2.43, 2.39 days, females longevity of 70.80, 56.75, 27.50 and reproduction of 24.2, 95.5 and 43.83 eggs/female on bean pods at 15, 20 and 30°C, respectively. Rearing of thrips on cotton leaves or cotton leaves plus pollen, at 27°C showed females’ longevity 30.0 or 38.3 days and 53.0 or 193.3 eggs/female, respectively (Trichilo and Leigh, 1988). Bene and Gargani (1989) reported that the females lived 30 days and had oviposited 35 eggs/female on chrysanthemum leaves at 25°C. Rijn Van et al. (1995) found 12.39 days total immature stage biological cycle at 25°C and 22.1 eggs/female net reproduction. Female adults of F. occidentalis when fed on chrysanthemum plus pollen at 25°C lived 31.4 days, which is about 4 days less than the respective value of our study and laid 135.6 eggs/female which is about 2.5 times greater than our findings (Robb, 1989). The addition of chrysanthemum pollen in the diet of F. occidentalis may provide the nutrients needed for increased reproduction. On the chrysanthemum, at 27.2°C, females lived 34 days and produced 125 eggs/female (Robb, 1989), while on cucumber leaves at 25°C, 38.7 eggs/female (Mollema et al., 1990) and on bean leaves at 25°C the life duration of female was estimated at 13.2 days and producing 32.4 eggs/female (Bünte et al., 1990). According to the results of Lowry et al. (1992) females of F. occidentalis on peanut, at 20 and 25°C, produced 7.7, 9.94 eggs/female and lived 2.2, 2.33 days respectively. The survival and reproduction of F. occidentalis was apparently restricted by the host used (peanut), which does not seem to be suitable for F. occidentalis. Food suitability is considered an important parameter for the longevity and reproduction of various insect species (Andrewartha, 1935; Gawaad and Shazil, 1969a; Teulon and Penman, 1991; Omkar and Srivastava, 2003). Brødsgaard (1994) reported that in the case of F. occidentalis on bean leaves at 25°C under different photoperiods 4:20, 8:16 and 16:8 (L:D), the preoviposition period was 1.9, 1.53, 1.65 days, female longevity 13.32, 9.63, 10.8 days and produced 22, 24.95 and 29.6 eggs/female, respectively. The study of F. occidentalis also on bean leaves at 25°C has shown that the oviposition period lasted 24.45 days, the females lived for 47.96 days, the reproduction was found 43.17 eggs/female and 1.76 eggs/female/day (Gerin et al., 1994), while on chrysanthemum the female longevity was 16 days (Arzone et al., 1989).
In comparison to the respective results of other researchers the results of our study showed a few differences. Probably, these differences are due to different rearing techniques, the different host-plants and the different conditions used in each experiment. Taking into account the differences of the results among the various studies, the most suitable host plants for F. occidentalis survival, development and reproduction, should be chrysanthemum and cotton when offered together with pollen. According to present results, F. occidentalis development and reproduction on cucumber are most favored at 20°C. Consequently, its population could increase with higher rate in glasshouses at temperatures around 20°C.
In the case of T. tabaci temperature had also a significant effect (similar to F. occidentalis) on duration of immature stage cycle, preoviposition-oviposition-postoviposition periods, on total female longevity as well as on reproduction dynamic and reproduction rate. Harris et al. (1936) reported that female longevity of T. tabaci was 19.9 days at 30°C, which is one day shorter than the respective value of our study. At mean temperature 21°C, preoviposition period lasted 3 days, oviposition 50 days, postoviposition 6 days and total females longevity of T. tabaci was 59 days (Sakimura, 1937). Gawaad and Shazli (1969b) in a relative study found that the preoviposition, oviposition, postoviposition and female longevity of T. tabaci lasted 21.1, 12.9, 3.7 and 18.7 days, respectively at 22°C, when fed on castor oil. At 25°C, RH 70% on the onion, the duration of females longevity was 16 days (Gawaad and Shazil, 1969a). Federov (1929), Franssen and Van Heurn (1933), Watts (1934), Ileri (1947), Lall and Singh (1960) reported that longevity of females ranged from 15 to 25 days. Lall and Singh (1968) recorded that female longevity of T. tabaci was 18.8 and 20.2 days at 15 and 30.8°C, respectively. Sakimura (1937) reported 80 eggs/female and 1.8 eggs/female/day at 18°C. On the castor oil plant at 22°C, T. tabaci produced 28.7 eggs/female, while at 25°C there were 13.4, 15.2, 16.0 and 24.0 eggs/female for stored onion bulbs, new onion bulbs, garlic bulbs, green leaves of stored onion bulbs and castor oil seedling, respectively (Gawaad and Shazil, 1969a,b). Rijn Van et al. (1995) found 12.90 days total immature stage biological cycle at 25°C and 27.5 eggs/female net reproduction. The highest reproduction rate 55 eggs/female of T. tabaci on the onion was observed during March with mean temperatures 23.4°C, while the lowest reproduction rate of 28.2 eggs/female was found during April with mean temperature was 30.8°C (Lall and Sing, 1968).
Similarly to data from F. occidentalis, differences were observed between the results on T. tabaci of our study with these of other researchers (Edelson and Magaro, 1988; Ileri, 1947) that can be obliged on the different conditions, the host plant or the different technique of rearing.
As it was shown from the results of this study, cucumber is a suitable host plant for survival and reproduction of both F. occidentalis and T. tabaci. Temperature was proved to be a very important factor affecting population densities of both F. occidentalis and T. tabaci. Decreased reproductivity at 15 and 30°C suggests that, normal early season and summer temperatures may inhibit their population increase. Conversely, warm periods in early spring and cool summer weather may trigger population build-up. Temperature affects in the same way both thrips species used, while total biological cycle (in days) of immature stages found almost the same at each temperature and female longevity of T. tabaci was proportionally always less than F. occidentalis.
REFERENCES
- Deligeorgidis, P.N., C.G. Ipsilandis, C. Fotiadou, G. Kaltsoudas, L. Giakalis and A. Garsen, 2005. Fluctuation and distribution of Frankliniella occidentalis (Pergande) and Thrips tabaci Lindeman (Thysanoptera: Thripidae) populations in greenhouse cucumber and tomato. Pak. J. Biol. Sci., 8: 1105-1111.
CrossRefDirect Link - Van Lenteren, J.C. and J. Woets, 1988. Biological and integrated pest control in greenhouses. Annu. Rev. Entomol., 33: 239-269.
CrossRefDirect Link - Mantel, W.P. and M. van de Vrie, 1988. De Californische thrips, Frankliniella occidentalis, een nieuwe schadelijke tripssoort in de tuinbouw onder glas in Nederland. Entomologische Berichten, 48: 140-144.
Direct Link - Omkar, G. and S. Srivastava, 2003. Influence of six aphid prey species on development and reproduction of a ladybird beetle, Coccinella septempunctata. BioControl, 48: 379-393.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Ames, Iowa, USA., ISBN-13: 9780813815602, Pages: 507.
Direct Link - Trichilo, P.J. and T.F. Leigh, 1988. Influence of resource quality on the reproductive fitness of flower thrips (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am., 81: 64-71.
Direct Link - Wagner, T.C., H. Wu, P.J. Sharpe, R.M. Schoolfield and R.N. Coulson, 1984. Modeling insect development rates: A literature review and application of a biophysical model. Ann. Entomol. Soc. Am., 77: 208-225.
Direct Link