
Rogelio Santiago
Campus Universitario de Pontevedra, E-36005 Pontevedra, Spain
Xose C. Souto
Campus Universitario de Pontevedra, E-36005 Pontevedra, Spain
Liliana Monetti
Mision Biologica de Galicia. Spanish Cormcil for Scientific Research (CSIC), Apartado 28, E-36080 Pontevedra, Spain
Bernardo Ordas
Mision Biologica de Galicia. Spanish Cormcil for Scientific Research (CSIC), Apartado 28, E-36080 Pontevedra, Spain
Amando Ordas
Mision Biologica de Galicia. Spanish Cormcil for Scientific Research (CSIC), Apartado 28, E-36080 Pontevedra, Spain
Rosa A. Malvar
Mision Biologica de Galicia. Spanish Cormcil for Scientific Research (CSIC), Apartado 28, E-36080 Pontevedra, Spain
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 4 | Page No.: 281-289







ABSTRACT
The objective of this research was to evaluate the effect of four phenols previously identified at higher concentrations in maize resistant genotypes (p-coumaric acid, ferulic acid, p-hydroxybenzaldehyde and vanillin) on the growth and development of S. nonagrioides larvae. Three different concentrations of each phenolic compound and a mixture of the four were evaluated. Data were recorded on larval and pupal weight, time to pupation and percentage of pupation and mortality. Growth and survival showed no significant differences between the larvae reared on diets with the different treatments and the control. It was concluded that these four compounds, although found in higher concentrations in resistant genotypes, were not toxic to S. nonagrioides in free form under laboratory conditions. Possible ways these compounds could be related with the S. nonagrioides resistance are discussed.
PDF Abstract XML References
How to cite this article
Rogelio Santiago, Xose C. Souto, Liliana Monetti, Bernardo Ordas, Amando Ordas and Rosa A. Malvar, 2006. Effect of Maize Pith Free Phenols on Larval Growth and Development of Sesamia nonagrioides (Lepidoptera: Noctuidae). Journal of Entomology, 3: 281-289.
DOI: 10.3923/je.2006.281.289
URL: https://scialert.net/abstract/?doi=je.2006.281.289
DOI: 10.3923/je.2006.281.289
URL: https://scialert.net/abstract/?doi=je.2006.281.289
INTRODUCTION
Sesamia nonagrioides (Lefèbvre) (Lepidoptera: Noctuidae) is one of the most harmful pests of maize, Zea mays L., in Spain and other Mediterranean regions (Cordero et al., 1998; Albajes et al., 2002; Gianessi et al., 2003). In Northwestern Spain this stem borer has two generations per year. The most important is the second generation larvae, which feed on the pith during plant development, reducing plant growth, grain size and causing indirect yield losses as a consequence of lodging (Larue, 1984). Although variability in resistance to this stem borer has been detected among populations and inbred lines of maize (Malvar et al., 1993; Cartea et al., 1994; Butrón et al., 1999), little is understood about the phytochemical nature of this resistance. Laboratory bioassays are an essential component of investigations of the chemical basis of resistance to insects in crop plants. Plant resistance studies have often focused on the isolation and identification of biologically active compounds in agricultural crops that could inhibit growth and development of insects that feed on plant tissues (Harris, 1979; Schoonhoven et al., 1998). The defense functions of 2,4-dihydroxy-7-methoxy- (2H)-1,4-benzoxazin-3- (4H)-one (DIMBOA) and phenolic acids in maize plants have been well documented for European corn borer Ostrinia nubilalis (Hubner) (Bergvinson, 1993; Bergvinson et al., 1994; Klun and Robinson, 1969), southwestern corn borer Diatraea grandiosella (Dyan), sugarcane borer Diatraea saccharalis (Fabricius) (Ramputh, 2002), pink stem borer S. nonagrioides (Ortego et al., 1998) and the maize weevil Sitophilus zeamais Motschulsky (Arnason et al., 1997).
DIMBOA is found in maize as a glycoside at high levels during the early stages of plant development; however DIMBOA concentration decreases as the plant grows, so this compound fails to protect the plants from the attack of the second generation of some lepidopteran pests (Mihm, 1985). Phenolic compounds are present in plant cells in four different forms: insoluble cell wall-bound phenolics, soluble ester-bound phenolics, soluble glycoside-bound phenolics and free phenolics. Most studies on maize resistance mechanisms have been focused on bound phenolics due to their role on cell wall cross-linking, providing a physical barrier to diseases and insect feeding (Bergvinson et al., 1994; Ramputh, 2002; Fry et al., 2000; Bily et al., 2003). On the other hand, free phenolic acids have low molecular weights and are partially soluble in many solvents, including water. These characteristics are important properties of plant toxins because they must pass through cell membranes to be readily available (Rhoades and Cates, 1976). Some phenols which act as toxins to some species can also act as phagostimulants to others. For example, cinnamic acid stimulates feeding by Scolytus multistriatus (Marsham) (Norris, 1977) but inhibits pupation in Agrotis ipsilon (Hufnagel) (Reese, 1977). p-Coumaric and ferulic acid were phagostimulants for the spotted stem borer, Chilo partellus (Swinhoe) (Torto et al., 1991), while they inhibit feeding of S. zeamais (Serratos et al., 1987). p-Coumaric acid was toxic to the two spotted spider mite, Tetranychus urticae Koch (Leszczynski et al., 1988), as ferulic acid was toxic to the dried fruit beetle, Carpophilus hemipterus (L.) (Dowd, 1990). Chorogenic acid is considered to be allelopathic (Einhellig, 1985), but it stimulates feeding by Leptinotarsa decemlineata (Say) (Hsiao and Fraenkel, 1968) and Lema trilineata Olivier (Kogan and Goeden, 1971). Furthermore, other free phenols were toxic to different insect species (Campbell et al., 1984, Fischer et al., 1990).
Early studies suggested the possibility that some natural acids with phenolic structure could be related with resistance to S. nonagrioides; at the same time, they showed inhibition of the fungal genera Fusarium (Anglade and Molot, 1967). Furthermore, recent studies put forward a possible role of free phenols in S. nonagrioides resistance due to the higher level of these compounds in the pith of genotypes resistant to this pest (Santiago et al., 2005). To determine if these compounds have an antibiotic effect, insect growth and development must be assessed. The goal of this study was to determine the possible effects of selected phenols on S. nonagrioides larval growth and development.
MATERIALS AND METHODS
Selected Compounds
Four phenolic compounds identified and quantified in previous studies in the pith of maize were selected for laboratory feeding trials (Fig. 1). p-Coumaric (pCA) and ferulic acid (FA) were the most abundant free phenols found in resistant genotypes in extensively studies with thirteen maize inbred lines (Santiago et al., 2005). Early studies showed p-hydroxybenzaldehyde (pHBA) and vanillin (VA) at low, but higher concentrations in resistant genotypes during some trials, so these compounds were also incorporated into meridic diets. Levels detected in the pith tissue and higher and lower concentrations were tested.
Laboratory Bioassays
Laboratory tests were conducted to investigate the antibiotic effect of the four phenols on the growth and development of S. nonagrioides larvae. Commercial phenolics purchased from Sigma-Aldrich Química, SA (Madrid, Spain) were dissolved in water with a sonicator and added to the artificial diet (Eizaguirre, 1989). Sixteen treatments were tested.
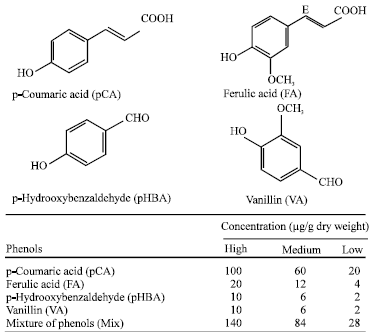 | |
| Fig. 1: | Chemical structure and concentrations of the different phenols added to meridic diets in the laboratory bioassays |
Twelve consisted of diets containing just one of the four phenols at low, medium or high concentration (Fig. 1). Besides the effect of a particular compound, it is also important to consider the combinations and interactions among compounds, which can be responsible for the appearance of a toxic effect (Cates, 1996; Nelson and Kursar, 1999). A mixture (Mix) of the four compounds at low, medium and high concentration was also tested, adding three treatments to the experiment (Fig. 1). Finally, artificial diet with distilled water was used as control (Control).
All treatments and control were evaluated in tests of ten replications each, arranged in a randomized complete block design. Approximately 10 g portions of diet per treatment and replication were placed into 9 cm-diameter Petri dishes. A 10-day-old larvae was placed into each dish, the dishes were sealed with parafilm and placed in a environmental controlled room at 26°C with a photoperiod of 16:8 (L:D). Fresh diet was provided every week so that all larvae ate as much as they wanted. Larval weight and the number of dead and missing larvae (larvae that escape from the dishes), were recorded weekly until pupation. Pupal weight and time to pupation were also registered.
Statistical Analysis
Repeated measures analysis was used to analyze the weekly weight measurements of larvae. A growth curve of weight on time was estimated for each treatment and homogeneity of linear and quadratic coefficients were tested for each pair of treatments. The analysis was made using the MIXED procedure of SAS (SAS Institute, 2000). All factors were considered random except for treatment which was considered fixed. The covariance was calculated following Littell et al. (1996). Additionally, analysis of variance and comparisons of means among treatments were calculated for larval weight. Larval weight means was adjusted by initial larval weight by analysis of covariance (SAS Institute, 2000). An analysis of variance and comparison of means among treatments were also calculated to analyze larval survival (proportion of total larval mortality, mortality during the first week and proportion of larvae that reach pupae stage).
Finally, to analyze larval survival and time to pupation, the Kaplan-Meier estimates of the survival and time to pupation functions were calculated for each treatment and curves were compared using the log rank test (Ordás et al., 2002; Cantor, 1997). Survival functions were significantly different if they deviated from the expected values of the null hypothesis (that survival functions are equivalent in all treatments). The statistic determines whether differences between survival functions are significantly different at any probability level, thus indicating that larvae have significantly different survival in some treatments than in others (LIFETEST procedure of SAS). Missing larvae were censured for larval survival analysis, which means that the analysis considered that larvae lived at least until they disappeared. When larvae reached pupal stage, the larvae survival was scored as reaching 42 days. Missing and dead larvae were removed before analyzing time to pupation data.
RESULTS
The growth of the larvae reared on each treatment fitted a parabolic function (Table 1). The linear parameters were positive and higher than the quadratic ones, the latter being negative. This means that the weight of larvae increased quickly in the first days, but larval weight remained almost invariable for many days thereafter. The coefficient of determination (r2) varied from 0.62 to 0.75 (data not shown).
Larvae fed on diets containing pCA and FA showed similar growth patterns. Larvae reared on diet containing low concentrations of both phenols had a linear growth, while the quadratic coefficient was not significant (Table 1). The linear coefficient for diet containing the lowest concentration of FA was significantly smaller than the linear coefficient for the highest concentration. However, the highest concentration showed a significant negative quadratic coefficient. In general, for each week, weight did not show significant differences among diets containing pCA or FA (Table 1).
The linear and quadratic coefficients of larval growth curves for pHBA at different concentrations were homogeneous. However, larval weights at 7 and 14 days were lower on diets with high than with low pHBA concentrations (Table 1) and larval weight at 21 days on diets with the highest pHBA concentration was significantly lower than on diets with the lowest pHBA concentration. However, weight of larvae fed on this compound was not significantly different from the control.
| Table 1: | Larval weight at five dates adjusted by initial larval weight and linear and quadratic coefficients±SD of larval weight curves on time (mg) for each phenol at different concentrations |
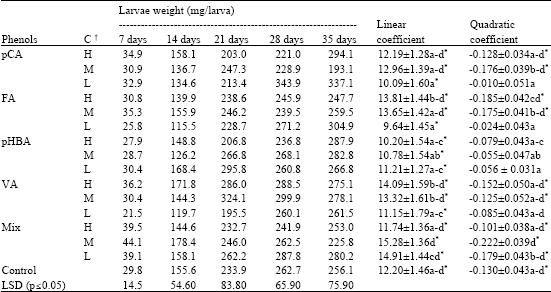 | |
| †, High (H), Medium (M) and Low (L) concentrations (Fig. 1), Coefficients within a column followed by the same letter are homogeneous (p≤0.05), | |
Although regression coefficients were homogeneous for the three VA concentrations on diets, larvae fed on them showed the opposite trend to larvae reared on pHBA diets (Table 1). Larval weight was less for the lowest concentration than for the higher ones. The larval weight at 21 and 28 days were significantly smaller for the low concentration than for any other concentration (Table 1).
The mixture of phenols showed the highest linear coefficients at medium concentration, having the highest increase on the larval weight just during the first week (Table 1).
Larval mortality did not significantly differ between treatments (Table 2). The range varied from 15-17% of dead larvae when fed on the mixture of phenols (Low: 28 μg g-1 dry weight) and FA diets (High: 20 μg g-1 dry weight) to 37-39% of dead larvae on the highest concentrations of pHBA (High: 10 μg g-1 dry weight) and 1 diets (Low: 20 μg g-1 dry weight). A total larval mortality is an important trait to analyze, however most of the deaths took place during the first week, when the larvae are most susceptible to toxic compounds. During this period, there was higher mortality when they fed on pHBA diets than when they fed on FA or Mixed diets, especially for the highest pHBA concentration (Table 2), although non significant differences were observed.
Survival data for larvae fed on different treatments were not equivalent. Table 2 shows the values of the log rank statistic in each treatment as well as the comparisons between treatments. The positive values of the statistic indicate a number of dead larvae larger than the number expected under the null hypotheses of equivalent survival distributions, while negative values indicate there were fewer deaths than expected under the null hypothesis. For example, there would be about 4 to 7 deaths above expected among larvae fed on pHBA diets, if survival functions were homogeneous. In conversely, there were around 2 to 5 deaths below that expected among larvae reared on Mix and FA diets (Table 2). Finally, in the comparisons between treatments the highest value of the log rank statistic was obtained for larvae reared on diets with the highest concentration of pHBA.
Time to pupation was also studied using a survival analysis. Survival data often consists of a variable response that measures the length of time until a specified event occurs, in this case the event was pupation. Although the lowest value of time to pupation was obtained in larvae reared on diets with 100 μg g-1 dry weight of pCA, the lowest values of log rank statistic at any concentration were shown in larvae fed on pHBA diets. In contrast, the highest values were in larvae reared on VA diets (Table 2). However, even though we could detect some possible trends in the survival and time to pupation log rank data, these values were not significantly different from the control.
| Table 2: | Proportion of total larval mortality, mortality during the first week and larvae that reach pupae state (±SE) and values of the log-rank statistic for homogeneity of survival and time to pupation curves for each phenol at different concentrations |
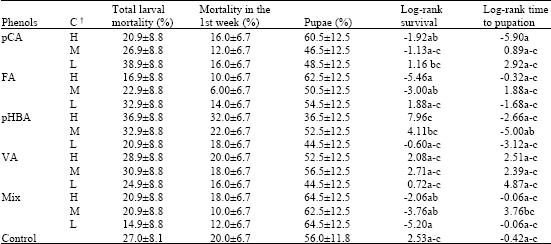 | |
| †, High (H), Medium (M) and Low (L) concentrations (Fig. 1), Log-rank statistic within a column followed by the same letter means that curves are homogeneous (p≤0.05), Phenols: p-coumaric (pCA), ferulic acid (FA), p-hydroxybenzaldehyde (pHBA) and vanillin (VA) | |
Pupal weight did not show significant differences among larvae reared on the different diets (data not shown). Pupation occurs when larvae reach around 300 mg independently of larval feeding. Proportion of larvae that reach the pupae stage was lower for the larvae fed on diets with the highest concentration of pHBA than larvae fed on diets with the highest concentration of pCA or FA; or any concentration of the Mix (Table 2).
DISCUSSION
Todd et al. (1971) found many phenolic acids present in higher plants to be toxic to the greenbug, Schizaphis graminum (Rodani) and many of them were present in resistant barley strains. Lesczynski et al. (1988) confirmed a toxic effect of pCA to the spider mite, T. urticae and later, Lege et al. (1995) found higher concentrations of this compound in cotton resistant genotypes to this pest. In maize, Serratos et al. (1987), studying the feeding response of the maize weevil, S. zeamais, clearly demonstrated that pure phenolic substances have significant antifeedant properties at a concentration close to the phenolic content in the grain. In this study, although a negative trend could be suggested for the highest concentration of pHBA, present findings indicate that none of the phenolics or mixtures incorporated into artificial diets had any statistically significant effect on growth, final weights or survival, compared to control.
These results are surprising, in that such an extensive body of evidence has been accumulated indicating that these compounds are toxic when incorporated into artificial diets (Dreyer et al., 1981; Serratos et al., 1987; Arnason et al., 1992), as well as previous studies showed higher concentration of free phenols in maize resistant genotypes (Santiago et al., 2005).
It is possible that the phenolic toxicity could be highly susceptible to modification by other dietary constituents. Different studies have shown that the presence of other plant oxidative enzymes, the type of dietary protein and the presence of antioxidants are determinants of phenolic toxicity (Felton et al., 1992; Appel, 1993; Summers and Felton, 1994). However, Warnock et al. (2001), studying the development of O. nubilalis larvae reared on diets containing liophilized maize silk tissues and pCA or FA additions, proved no effect of these compounds on larvae weight, even with higher concentrations than that tested in the present study. These results agree with the lack of an antifeedant effect of free pCA and FA in our laboratory test.
In relation with the higher levels of the four compounds in resistant genotypes, these compounds did not show any effect on S. nonagriodes development as free phenolics. Studying the occurrence of phenolics in plants is useful to distinguish between soluble and bound forms of a compound. Distinction is important since its role in resistance could be structural or biochemical. At a structural level the phenolic constituents of the cell wall of graminaceous plants consist largely of pCA and FA (Hartley and Jones, 1978). Both acids take part in mechanical resistance through cell wall fortification and lignification (Akin et al., 1992; Bergvinson et al., 1994, 1995). In this regard, Santiago et al. (2005) suggested that higher quantities of free pCA in resistant inbreds may perform a pool from which the other various hydroxycinnamic acids and their esters were formed (Bate-Smith, 1962). Thus, although these compounds did not show a direct antibiotic effect in the current study, they could still be related to borer resistance. In fact, recent studies with maize pith have pointed out higher quantities of cell wall phenylpropanoids in resistant genotypes to S. nonagrioides (Santiago et al., 2006).
The major feeding deterrent to greenbug, S. graminum, isolated from sorghum leaves was pHBA (Dreyer et al., 1981); while VA is known as a chemioatractant (Eveland and Haseeb, 1993), even carrying out a sexual pheromone function (Ubik et al., 1975). In the present study these two compounds showed no effects on the growth and development of S. nonagrioides larvae. In the same way, the mixture containing a combination of all phenols did not show any kind of synergistic or antagonistic effect at the tested concentrations.
In summary, our findings demonstrate that the free phenols tested do not have an apparent effect on the larval performance of the lepidopteran S. nonagrioides. However, this does not rule out the possibility that these compounds may be present in high concentration in resistant genotypes to play a structural role in the resistance to this pest. This issue is currently under further evaluation.
ACKNOWLEDGMENTS
The authors thank Emma Muiños for helping in rearing insects. R. Santiago, L. Monetti and B. Ordás acknowledge fellowships from the Autonomous Government of Galicia, Agencia Española de Cooperación Internacional and CSIC respectively. This research was supported by the National Plan for Research and Development of Spain (AGL2001-3736, AGL2003-0961) and the Autonomous Government of Galicia (PGIDIT04PXIC40303PN).
REFERENCES
- Akin, D.E., R.D. Hartley, L.L. Rigsby and W.H. Morrison, 1992. Phenolic acids released from bermudagrass (Cynodon dactylon) by sequencial sodium hydroxide treatment in relation to biodegradation of cell types. J. Sci. Food Agric., 58: 207-214.
Direct Link - Albajes, R., M. Konstantopoulou, O. Etchepare and M. Eizaguirre et al., 2002. Mating disruption of the corn borer Sesamia nonagrioides (Lepidoptera: Noctuidae) using sprayable formulations of pheromone. Crop Prot., 21: 217-225.
CrossRefDirect Link - Appel, H.M., 1993. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol., 19: 1521-1552.
CrossRefDirect Link - Arnason, J.T., J. Gale, B.C. de Beyssac, A. Sen and S.S. Miller et al., 1992. Role of phenolics in resistance of maize grain to the stored grain insects, Prostephanus truncatus (Horn) and Sitophilus zeamais (Motsch). J. Stored Prod. Res., 28: 119-126.
CrossRef - Bergvinson, D.J., R.I. Hamilton and J.T. Arnason, 1995. Leaf profile of maize resistance factors to European corn borer, Ostrinia nubilalis. J. Chem. Ecol., 21: 343-354.
Direct Link - Bily, A.C., L.M. Reid, J.H. Taylor, D. Johnston and C. Malouin et al., 2003. Dehydrodimers of ferulic acid in maize grain pericarp and aleurone: Resistance factors to Fusarium graminearum. Phytopathology, 93: 712-719.
CrossRefDirect Link - Butron, A., R.A. Malvar, M.E. Cartea, A. Ordas and P. Velasco, 1999. Resistance of maize inbreds to pink stem borer. Crop Sci., 39: 102-107.
Direct Link - Cartea, M.E., R.A. Malvar, P. Revilla, A. Ordas and A. Alvarez, 1994. Seasonal occurrence and response of maize inbred lines to pink stem borer in the northwest of Spain. Maydica, 39: 191-196.
Direct Link - Cates, R.G., 1996. The role of mixtures and variation in the production of terpenoids in conifer-insect-pathogen interactions. Recent Adv. Phytochem., 30: 179-216.
Direct Link - Cordero, A., R.A. Malvar, A. Butron, P. Revilla, P. Velasco and A. Ordas, 1998. Population dynamics and life-cycle of corn borers in South Atlantic European coast. Maydica, 43: 5-12.
Direct Link - Dowd, P.F., 1990. Responses of Carpophilus hemipterus larvae and adults to selected secondary metabolites of maize. Entomol. Exp. Applied, 54: 29-36.
Direct Link - Dreyer, D.L., J.C. Reese and C.J. Kenneth, 1981. Aphid feeding deterrents in sorghum bioassay isolation and characterization. J. Chem. Ecol., 7: 273-284.
CrossRefDirect Link - Eveland, L.K. and M.A. Haseeb, 1993. Pellioditis pellio: Substituted phenolic compounds as chemoattractants. J. Chem. Ecol., 19: 2543-2546.
CrossRefDirect Link - Felton, G.W., K. Donato, R.M. Broadway and S.S. Duffey, 1992. Impact of oxidized plant phenolics on the nutritional quality of dietar protein to a noctuid herbivore, Spodoptera exigua. J. Insect Physiol., 38: 277-285.
CrossRefDirect Link - Fry, S.C., S. C.Willis and A.E.J. Paterson, 2000. Intraprotoplasmic and wall-localized formation of arabinoxylan-bound diferulates and larger ferulate coupling-products in maize cell-suspension cultures. Planta, 211: 679-692.
Direct Link - Hsiao, T.H. and G. Fraenkel, 1968. The role of secondary plant substances in the food specificity of the Colorado potato beetle. Ann. Entomol. Soc. Am., 61: 483-493.
Direct Link - Kogan, M. and R.D. Goeden, 1971. Feeding and host-selection behaviour of Lema trilineata daturaphila larvae (Coleoptera: Chrysomelidae). Ann. Entomol. Soc. Am., 64: 1435-1448.
Direct Link - Lege, K.E., J.T. Cothreen and C.W. Smith, 1995. Phenolic acid and condensed tannin concentrations of six cotton genotypes. Environ. Exp. Bot., 35: 241-249.
CrossRefDirect Link - Nelson, A.C. and T.A. Kursar, 1999. Interactions among plant defence compounds: A method for analysis. Chemoecology, 9: 81-92.
CrossRefDirect Link - Ordas, B., A. Butron, P. Soengas, A. Ordas and R.A. Malvar, 2002. Antibiosis of the pith maize to Sesamia nonagrioides (Lepidoptera: Noctuidae). J. Econ. Entomol., 95: 1044-1048.
Direct Link - Ortego, F., M. Ruz and P. Castanera, 1998. Effect of DIMBOA on growth and digestive physiology of Sesamia nonagrioides (Lepidoptera: Noctuidae) larvae. J. Insect Physiol., 44: 95-101.
PubMedDirect Link - Santiago, R., R.A. Malvar, M.D. Baamonde, P. Revilla and X.C. Souto, 2005. Free phenols in maize pith and their relationship with resistance to Sesamia nonagrioides (Lepidoptera: Noctuidae) attack. J. Econ. Entomol., 98: 1349-1356.
Direct Link - Santiago, R., A. Butron, J.T. Arnason, L.M. Reid, X.C. Souto and R.A. Malvar, 2006. Putative role of pith cell wall phenylpropanoids in Sesamia nonagrioides (Lepidoptera: Noctuidae) resistance. J. Agric. Food Chem., 54: 2274-2279.
CrossRefDirect Link - Serratos, A., J.T. Arnason, C. Nozzolillo, J.D.H. Lambert and B.J.R. Philogene et al., 1987. Factors contributing to resistance of exotic maize populations to maize weevil, Sitophilus zeamais. J. Chem. Ecol., 13: 751-762.
CrossRefDirect Link - Summers, C.B. and G.W. Felton, 1994. Prooxidant effects of phenolic acids on the generalist herbivore Helicoverpa zea (Lepidoptera: Noctuidae): Potential mode of action for phenolic compounds in plant anti-herbivore chemistry. Insect Biochem. Mol. Biol., 24: 943-953.
Direct Link - Todd, G.W., A. Getahun and R.C. Cross, 1971. Resistance in barley to the greenbug, S. graminum. Toxicity of phenolic and flavonoid compounds and related substances. Ann. Entomol. Soc. Am., 64: 718-722.
Direct Link - Torto, B., A. Hassanalli, K.N. Saxena and S. Nokoe, 1991. Feeding responses of Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) larvae to sorghum plant phenolics and their analogs. J. Chem. Ecol., 17: 67-78.
CrossRefDirect Link - Ubik, K., J. Vrkoc, J. Zdarek and C. Kontev, 1975. Vanillin: Possible sex pheromone of an insect. Naturwissenschaften, 62: 348-348.
CrossRefDirect Link - Warnock, D.F., W.D. Hutchison, C.B.C. Tong and D.W. Davis, 2001. Evaluating maize for allelochemicals that affect European corn borer (Lepidoptera: Crambidae) larval development. Crop Sci., 41: 1761-1771.
Direct Link