
A. David Adebote
Department of Biological Sciences, Alnnadu Bello University, Zaria,
800001 Kaduua State, Nigeria
J. Sonnie Oniye
Department of Biological Sciences, Alnnadu Bello University, Zaria,
800001 Kaduua State, Nigeria
S. Iliya Ndams
Department of Biological Sciences, Alnnadu Bello University, Zaria,
800001 Kaduua State, Nigeria
K. Mary Nache
Department of Biological Sciences, Alnnadu Bello University, Zaria,
800001 Kaduua State, Nigeria
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 2 | Page No.: 180-188







ABSTRACT
Larval mosquitoes species (Diptera: Culicidae) breeding in specific containers in the peridomestic environments and larval Aedes aegypti L. indices predictive of potential yellow fever epidemics were investigated in Bomo, Hanwa, Jama’a and Samaru settlements around Zaria, Nigeria. Systematic sampling, using small soup laddle dipper was adopted in the examination of every container with water, including wells and tree-holes, for larval mosquitoes in the four villages. Aedes aegypti, Culex horridus Edwards, Culex nebulosus Theobald, Culex quinquefasciatus Say and Culex tigripes Grandpre, larvae were encountered in 44(14.97%) and 14 varieties of the 294 containers examined, whose surface areas ranged from 0.005 to 1.28 m2. The mean larval population of the five container-breeding mosquitoes species differ significantly (p<0.05). Aedes aegypti and C. nebulosus showed highest preferences for breeding in automobile tyres; C. horridus and C. quinquefasciatus larvae occurred most in wells. Aedes aegypti as sole species occurred in 20(45.45%) and 11 varieties of the containers. Culex horridus, C. nebulosus and C. quinquefasciatus as sole species were found in 6(13.64%), 2(4.55%) and 2(4.55%) containers, respectively. A maximum of three larval mosquitoes combinations also occurred in same container habitats, highlighting both conspecific and heterospecific breeding patterns for the mosquitoes. Of the one hundred and twenty one (121) houses inspected, 20(16.53%) had larvae of A. aegypti in their peridomestic containers; with larval indices (total house index = 16.53%, total container index = 9.52%, total Breteau index = 23.14%) for the four villages within the limit predictive of potential epidemic yellow fever outbreak.
PDF Abstract XML References
How to cite this article
A. David Adebote, J. Sonnie Oniye, S. Iliya Ndams and K. Mary Nache, 2006. The Breeding of Mosquitoes (Diptera: Culicidae) in Peridomestic Containers and Implication in Yellow Fever Transmission in Villages Around Zaria, Northern Nigeria. Journal of Entomology, 3: 180-188.
DOI: 10.3923/je.2006.180.188
URL: https://scialert.net/abstract/?doi=je.2006.180.188
DOI: 10.3923/je.2006.180.188
URL: https://scialert.net/abstract/?doi=je.2006.180.188
INTRODUCTION
Vector-borne infections constitute a substantial number amongst the globally emerging and resurging diseases of public health importance (Gratz, 1999). These diseases require constant surveillance and the development of effective control methods. The emergence of new vector-borne disease entities and the resurgence of old ones, depend on ecological changes that increase vector densities, climate, immunity status, human and potential vector population densities and the presence of suitable reservoir (Gratz, 1999). However, the prime factor in the establishment of infection in a new focus is the presence of a suitable vector in densities capable of transmitting the infection (Gratz, 1999). In some areas, efficient vectors are present in the absence of the disease(s) they transmit; but these areas become threatened on the introduction of the infectious agents (Romi et al., 1997).
Chikungunya, dengue, dengue haemorrhagic fever and yellow fever are amongst the emerging and resurging diseases vectored by several Aedes (Stegomyia) (Diptera: Culicidae) mosquitoes, particularly Aedes aegypti L., in Africa, Asia and the Americas (Kouri et al., 1986; Savage and Smith, 1995; Gratz and Knudsen, 1996). Aedes aegypti has utilized the opportunities afforded by unplanned urbanization in most tropical countries and the associated poor solid waste collections, which yield waste containers, to breed in profusion (Hii, 1977; Service, 1990). Breeding opportunities for A. aegypti and other container-breeding mosquitoes are often expanded by inadequate piped water supplies in rapidly expanding areas of poor housing; as well as the storage of rain water, water from wells and street standpipes in a variety of containers for domestic use (Barrera et al., 1993).
These inadequacies, which create conditions that favour the breeding of mosquitoes, exist in the peridomestic environment of many settlements in Nigeria. The bionomics of container-breeding mosquitoes, especially in peridomestic environments in Nigeria are poorly known. Service (1974) found both A. aegypti and Aedes vittatus Bigot as potential vectors of yellow fever, breeding as larvae in water pots and abundant as early evening man-biting adults in six areas north-west of Nigeria. Peterson and Lambrecht (1976) observed that traditional water storage in pots in settlements in and around Enugu, south-eastern Nigeria, provided all year round peridomestic breeding sites for 18 species of mosquitoes. In a recent survey, Anyanwu et al. (1999) found 15 species of mosquitoes predominated by Culex pipiens fatigans Wiedemann (Synonym = C. quinquefasciatus Say) in human residences in Zaria. This study, which was not restricted to the peridomestic environment, also revealed A. aegypti larvae in man-made receptacles.
There had been occurrences of sporadic and unpremeditated epidemics of yellow fever, transmitted partly by A. aegypti, Aedes africanus Theobald and Aedes luteocephalus Newstead, at variable spatiotemporal scales in Nigeria (MacNamara, 1954, Monath et al., 1973; WHO, 1986; DeCock et al., 1988; Nasidi et al., 1989 Service, 1993). However, there are no existing large-scale vector control operations and entomological surveillance activities for monitoring the resurgence of the disease in many parts of the country. This study reports the species of mosquitoes breeding in containers in the peridomestic environments of four villages around Zaria, northern Nigeria. Breeding indices of A. aegypti were also utilized in this study, as a surveillance strategy, to assess yellow fever epidemic risk potential of villages that had hitherto not experienced epidemics of the disease.
MATERIALS AND METHODS
Study Area
Between June and September, 2002, surveys of larval mosquitoes breeding in containers in peridomestic environments were conducted in Hanwa (11°08' N, 07°43' E) Jama’a (11°14' N, 07°32' E), Samaru (11°10'N, 07°39' E) and Bomo (11°12'N 07°38' E) villages located north-west at 7, 13, 14 and 18 km, respectively through the major vehicular road away from the ancient wall of Zaria city (11°07' N, 07°44' E), north-central Nigeria. The villages are located in the northern Guinea savanna of Nigeria. Hanney (1960) and Service (1963; 1965) gave the general descriptions and climate of the northern Guinea savanna monitored in Kaduna, Nigeria (about 80 km south of Zaria). To account for climatic changes that might have occurred since these publications and to be specific for Zaria, data of weather variables monitored in loco at the Samaru campus of the Ahmadu Bello university meteorological unit are herein presented. The total and mean annual rainfall are approximately 974 and 81.1 mm, respectively, falling mainly in June-October, when the temperature ranges from 15.2 to 37.4°C (mean = 21-32.5°C), with a mean relative humidity (RH) of 56.6 to 66%. The other months of the year are dry (<10% of annual rainfall) mainly in March - May; mean RH = 35.7-50%, with mean temperature reaching 33.4°C. Mean sunshine hours for the area being 6.5. Between April and September, winds blew mainly in the south-western and north-western directions while in October to March the predominant winds are the north-easterlies bringing the harmattan cold-cum-dusts. The major river draining the villages is River Kubanni, which flows close by Samaru, Jama’a and Hanwa. Bomo is situated at the edge of a large permanent lake (Bomo lake). The human population in the villages is gregarious and mainly agricultural. Maize, sorghum, millet and cowpeas are the main crops, mostly cultivated around the villages and in some cases, close to family compounds. Domestic animals are mainly goats, sheep, cattle, dogs and poultry. Occasionally there are donkeys and horses in the villages; and pigs exclusively in Samaru. The indigenous populations live mostly in rectangular, often windowless mud houses, but inhabitants of Samaru and Jama’a also live in modern houses built with cement blocks. The village size and density of human population is highest in Samaru. The Zaria waterworks irregularly supply all the villages, except Bomo, with pipe-borne water. Bomo was recently, in 2000, provided with two boreholes with manual pumps. All the villages have several wells, 8-20 m deep, sunk by government agencies and through community self-help.
Sampling
A systematic sampling technique by selecting every other house was adopted for larval mosquito searches of houses in different areas of each village, so as to obtain a representative incidence of breeding in the villages In every selected house, all containers with water, including wells and tree-holes within 10 m radius of each house (peridomestic), were sought and examined, using flashlight where necessary, for mosquito breeding. Larval stages of mosquitoes were collected with a small soup laddle dipper 9 cm in diameter and holding 138 mL of water (Service, 1976). In the bigger containers, 10 dips were made per container and the larvae collected were concentrated with the aid of a large sieve and fixed in 70% alcohol in well-labelled specimen bottles in the field, for subsequent species identification in the laboratory. In the smaller containers and tree-holes, all the larvae were concentrated and fixed in alcohol in the field prior to species identification. Larvae in wells were caught by carefully lowering a bucket into the water and pulling it up against the sides of the well; 10 dips were then made, the larvae concentrated and fixed in 70% alcohol (Service, 1993). The types of positive containers and those negative but contained water were noted and their surface areas estimated with a tape metre rule.
Species Identification
All the larval mosquitoes were identified under the x50 magnification of a stereo-microscope with the guide of the pictorial keys of Hopkins (1952).
Data Analysis
Analysis of variance (ANOVA) statistic was employed to test for significant difference in density of larval mosquitoes in the various containers. Logarithm transformed data of the mean density of larvae in government - sunk and community - sunk wells were subjected to Student’s t-test to test for significant difference.
RESULTS
Five species of larval mosquitoes, including Aedes aegypti, L., Culex horridus Edwards, Culex nebulosus Theobald, Culex quinquefasciatus Say and Culex tigripes Grandpre, were found breeding in various peridomestic containers in four villages around Zaria, northern Nigeria. Aedes aegypti and C. tigripes being the most and least abundant species, respectively. Culex nebulosus was not encountered in containers in Jama’a while C. quinquefasciatus and C. tigripes were only found in containers in Bomo and Samaru, respectively (Table 1).
| Table 1: | Species composition of container-breeding larval mosquitoes in four villages around Zaria, Nigeria |
 | |
| Table 2: | Container preference of mosquitoes breeding in peridomestic environments around Zaria, Nigeria |
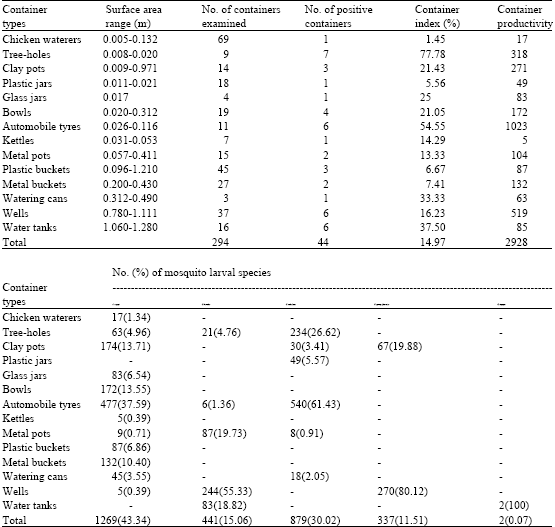 | |
| Table 3: | Breeding pattern of mosquitoes in peridomestic containers around Zaria, Nigeria |
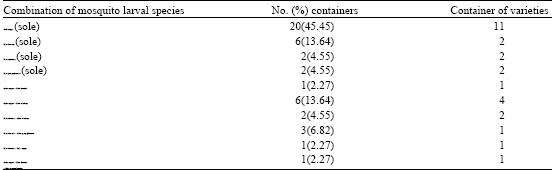 | |
| Table 4: | Aedes aegypti larval breeding indices in four settlements around Zaria, Nigeria |
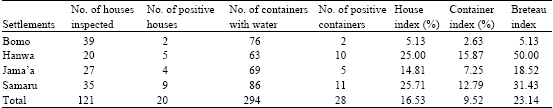 | |
| Table 5: | Comparison of government-sunk and community-sunk wells for mosquito breeding in Bomo village, Zaria, Nigeria |
A total of 294 containers of 14 varieties, with surface areas ranging from 0.005 to 1.28 m2 were examined for larval mosquitoes in the four villages. Forty-four (14.97%) of the containers supported the breeding of variable number of mosquito larvae. The most positive of the containers being tree-holes with 77.78% positivity, while chicken waterer was least, with 1.45% positivity. The most productive of the containers, supporting high populations of three larval mosquito species were automobile tyres (1023 larvae; mean = 170.5), wells (519 larvae; mean = 86.5) and tree-holes (318 larvae; mean = 45.4). Aedes aegypti, C. nebulosus, C. horridus, C. quinquefasciatus and C. tigripes bred in 12, 6, 5, 3 and 1 types of container, respectively (Table 2). The mean density of the five species of larval mosquitoes breeding in the 14 varieties of containers differ significantly (ANOVA, p<0.05).
Aedes aegypti bred as sole species in twenty (45.45%) of eleven varieties of containers. Culex horridus, C. nebulosus and C. quinquefasciatus also bred as sole species in 6(13.64%), 2(4.55%) and 2(4.55%) of the containers, respectively Aedes aegypti and C. nebulosus in combination occurred in 6(13.64%) of four varieties of containers. The duets of A. aegypti and C. horridus; and of C. horridus and C. tigripes occurred in only one (2.27%) container. The triad combination of A. aegypti, C. horridus and C. quinquefasciatus larvae occurred in one container (Table 3).
Out of 121 houses inspected, 20(16.53%) were positive for larval A. aegypti mosquitoes. Only twenty-eight (9.52%) of the 294 containers with water actually supported the breeding of A. aegypti. From the data of A. aegypti breeding, house indices ranging from 5.13% for Bomo to 25.71% for Samaru; container indices ranging from 2.63% for Bomo to 15.87% for Hanwa and Breteau indices ranging from 5.13 in Bomo to 50 in Hanwa were obtained. All the indices are within the critical ranges supportive of epidemic yellow fever outbreaks (Table 4).
Table 5 shows that 80% of the government-sunk wells and only 6.25% of the community-sunk wells supported the breeding of mosquitoes in Bomo village. The log (n + 1) transformed mean density of larvae obtained from both types of wells differed significantly (p<0.05) when subjected to Student’s t-test statistic.
DISCUSSION
This study demonstrates that five species of mosquito larvae breed in various water storage containers in the peridomestic environments of four villages around Zaria, northern Nigeria. Aedes aegypti is the most versatile in its choice of breeding containers and together with C. nebulosus have highest preferences for automobile tyres. Culex horridus and C.quinquefasciatus were amenable to wells as peridomestic breeding sites while C. tigripes only occurred in a water tank. The predominance of A. aegypti larvae in the habitats studied further confirms its preference for water storage containers within the vicinity of human habitations as breeding sites. Four of the five species of mosquitoes encountered in this study were amongst the species of mosquitoes found breeding in tree-holes by Service (1965) in Kaduna, Nigeria. Hanney (1960) and Service (1963) had earlier recorded them in domestic utensils and tree-holes in the northern Guinea savanna close to where this study was conducted. The five species were also amongst the 18 species of larval mosquitoes breeding in water pots and their covers in settlements in and around Enugu, south-eastern Nigeria (Peterson and Lambrecht, 1976).
Culex quinquefasciatus larvae were only found in clay pots and wells in Bomo village, a finding which is in contrast to its preference for foul and polluted water as breeding habitat. In Bomo village, which was the most deficient in polluted peridomestic surface run off and allied polluted water bodies amongst the villages studied, C. quinquefasciatus had utilized the opportunity of the availability of contaminated and abandoned wells to breed in profusion. In the other villages, the availability of numerous polluted water bodies had shifted the attention of the species from domestic containers to these alternatives and preferred breeding sites. Hanney (1960) found C. quinquefasciatus breeding in latrines in Kaduna, Nigeria. Aigbodion et al. (1995) observed that the species does not breed in domestic water storage containers and wells in Jos, Nigeria.
Culex quinquefasciatus in Nigeria is associated with urbanization and has replaced the once urban dominant C. nebulosus which is now restricted to breeding in villages (Service, 1963). This may explain the overt rarity of C. quinquefasciatus compared with C. nebulosus in village settings in which this study was conducted. Whereas Service (1974) found C. quinquefasciatus, in only a single pot, C. nebulosus was present in pots in six areas in eleven villages north-west of Nigeria. Culex nebulosus is nonhuman biting; its breeding close to human habitation may be associated with the availability of avian blood provided by domestic chickens and other poultry kept by residents of the villages. The chickens in turn are suspected to be predatory on the mosquito larvae while drinking in the waterers, as this category of containers were the least positive with mosquito larvae in the villages. Culex nebulosus has been reported to be preferentially ornithophilic (Service, 1963; Snow and Boreham, 1978). The species also showed bias towards calf-baited trap in Burkina Fasso (Costantini et al., 1998). Culex tigripes has a biological control potential but its occurrence only in Samaru and in very low numbers with the not medically important C. horridus precludes its biocontrol role. Service (1974) also encountered C. tigripes as the only aquatic predator of mosquito larvae, but concurred that it occurred in too few pots to have exercised much impact on larval populations of yellow fever vectors. Members of the Anopheles gambiae Giles complex or other species of Anopheles were not found breeding in containers in this study. Bruce-Chwatt (1957) observed the oviposition of Anopheles gambiae s.l. in pots in villages near Enugu. Service (1974) found larvae of An. gambiae s.l. in narrow necked pots in six villages north-west of Nigeria. Peterson and Lambrecht (1976) also recorded Anopheles larvae in waterpots in and around Enugu.
All the phytotelmata from which mosquito larvae were collected were from the flamboyant tree, Delonix regia (Bojer ex Hooker) (Leguminosae: Caesalpinioideae). Although, the present survey was restricted to trees found in the immediate vicinity of human residences, D. regia in a large-scale survey of the Zaria area, offered up to 94% of mosquito phytotelmata (Adebote et al., 2004). Delonix regia was not amongst the four species of trees from which Service (1965) observed tree-hole breeding mosquitoes in Kaduna, Nigeria.
The major sources of water for domestic use in Bomo village are wells. Every household in the village has at least one well and frequently more in many instances. There are few government-sunk wells in the village, which are often abandoned, contaminated with dirts and filthy materials, thus affording larval habitats for mosquitoes. These wells serve as reservoirs for both A. aegypti and C. quinquefasciatus vector species of mosquitoes. The high frequency of use to which the community-sunk wells are put might have served as oviposition deterrents for adult mosquitoes. This could be the reason for the lower larval mosquito densities in the community-sunk wells.
Larval indices of A. aegypti obtained for the four villages are within the critical ranges associated with yellow fever outbreaks. WHO (1986) gave a house index range of 4-35 and above; container index range of 3-20 and above; Breteau index range of 5-50 and above, as critical for yellow fever epidemics. It is therefore, suggestive that the four villages are yellow fever epidemic prone. More so, the northern Guinea savanna in which the villages are located constitute the epidemic zone, where man-to-man transmission of yellow fever by A. aegypti occurs as epidemics (WHO, 1986). A few adult A. aegypti were incidentally observed biting humans in the villages, which proved that the adult flies in the area are anthropophilic. Despite these positive propensities, epidemics of yellow fever has not been reported in the village. The absence of epidemics may be due to the absence of the causative virus in humans and or absence of monkey reservoir hosts. Vaccination of humans in the villages might be contributory to herd immunity thus boosting the immune status of the population against the disease. No monkey was sited throughout this survey but populations of patas monkeys, Erythrocebas patas were sited on the Dumbi inselberg, 20 km south of Zaria where a putative Ae. vittatus breed in rock pools. Considering the ease and frequency of human communication between Dumbi and Zaria it is plausible that the potential viral reservoir range could be that extensive. However, the epidemiological role of nonhuman primates in the transmission of yellow fever need to be ascertained in the study area. Apart from its yellow fever transmission potential, Anyanwu et al. (2000) had incriminated A. aegypti as a potential vector of the zoonotic dog filaria,. Dirofilaria repens. The presence of dogs and A. aegypti in close proximity with humans in the peridomestic environment studied, could support the transmission of human dirofilariasis.
In conclusion, it is pertinent to recommend measures that will help to ameliorate the epidemic risk posture of the villages examined in this study. Routine yellow fever vaccination as part of the primary health care should be intensified, as this will protect the inhabitants against possible outbreaks of the disease. Health education programme to improve public awareness of the disease and to encourage mass participation in basic sanitary measures and source reduction to prevent breeding of mosquitoes in peridomestic and carelessly discarded containers should be intensified. Insecticidal control of larval mosquitoes in containers including wells, using temephos (organophosphate) at a safe 1 mg L-1 concentration could be undertaken by skilled health officials in the identified villages.
ACKNOWLEDGMENTS
We thank Mr. Bala Bomo of the Bomo village healthcare centre, Mr. Aliyu Abdulmumin of Hanwa village, Mr. and Mrs. Bako Damina of Samaru, the village head of Jama’a and all the villagers for their assistance and cooperation during field surveys. Thanks are due to the Head of Soil Science Department of the Ahmadu Bello University, Zaria and staff of the meteorological unit for the provision of meteorologic data.w
REFERENCES
- Costantini, C., N. Sagnon, A.D. Torre, M. Diallo, J. Brady, G. Gibson and M. Coluzzi, 1998. Odor-Mediated host preferences of West African mosquitoes with particular reference to Malaria vectors. Am. J. Trop. Med. Hyg., 58: 56-63.
Direct Link - DeCock, K.M., T.P. Monath, A. Nasidi, P.M. Tukei and J. Enriquez et al., 1988. Epidemic yellow fever in Eastern Nigeria, 1986. Lancet, 1: 630-633.
PubMedDirect Link - Hanney, P.W., 1960. The mosquitoes of Zaria Province, Northern Nigeria. Bull. Entomol. Res., 51: 145-171.
CrossRefDirect Link - Service, M.W., 1963. The ecology of the mosquitos of the Northern Guinea Savannah of Nigeria. Bull. Entomol. Res., 54: 601-632.
CrossRef - Snow, W.F. and P.F.L. Boreham, 1978. The host-feeding patterns of some culicine mosquitoes (Diptera: Culicidae) in the Gambia. Bull. Entomol. Res., 68: 695-706.
CrossRefDirect Link