
Souleymane Nacro
Campus de Beaulieu, Faculte des Sciences,
Universite de Rennes I, Avenue du General Leclerc, F-34042 Rennes Cedex France
Jean-Pierre Nenon
Campus de Beaulieu, Faculte des Sciences,
Universite de Rennes I, Avenue du General Leclerc, F-34042 Rennes Cedex France
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 1 | Page No.: 16-22







ABSTRACT
The anatomy of the reproductive system of the adult female, the newly laid egg and its attaching system of the African Rice Gall Midge, Orseolia oryzivora were explored under photonic and electronic microscopes. The reproductive system of the female adult includes two voluminous and spherical ovaries. Each ovary is formed by a large number of ovarioles, which appear to be independent one from another. The oval laid egg of O. oryzivora is 700 μ long and 150 μ wide. Its external surface, formed by the exochorion is smooth. This exochorion, which is thin and regular, detaches easily from the underlying thicker and lighter endochorion. The egg is covered with a layer of mucus adhering to the plant surface and joining sometimes constricted accessory glands. The female adult of O. oryzivora lays its eggs in most cases singly or in small groups often connected by mucus, on the lower and upper surfaces of the rice leaf, the leaf-sheath and the stem. On the leaf, the eggs are always laid parallel to the axis of the leaf, along the central vein, between two lines of siliceous phytoliths.
PDF Abstract XML References
How to cite this article
Souleymane Nacro and Jean-Pierre Nenon, 2006. Anatomy of the Female Reproductive System and the Ultrastructure of the Egg’s Envelopes of Orseolia oryzivora (Diptera: Cecidomyiidae). Journal of Entomology, 3: 16-22.
DOI: 10.3923/je.2006.16.22
URL: https://scialert.net/abstract/?doi=je.2006.16.22
DOI: 10.3923/je.2006.16.22
URL: https://scialert.net/abstract/?doi=je.2006.16.22
INTRODUCTION
The Diptera are probably the most important of all the groups of insects that closely affect man. The majority of species in this order are harmless to man but some of them are vectors of diseases. Others are important pests of agricultural crops. Some few species such as Tachinidae are beneficial because they destroy large numbers of plant-feeding insects through their parasitic habit (Freeman, 1980). Their eggs are usually small and ovoid, with thin, unsculptured chorion and are typically deposited singly or in small masses on or near the larval food (Howel et al., 1978).
According to Gagne (1994), the Cecidomyiidae are possibly the largest family of flies. Ordinary, they lay their eggs singly or in clusters on or near the food or in the habitat of the larvae. In certain instances, they are deposited beneath the water, in the soil, inside the tissues of plants, on living hosts or other places. Free-living larvae, which feed in the stems, flowers, fruit and other parts of the plants, are gall makers. They produce galls of various shapes and sizes in the flowers and on the leaves, stems and roots of many plants. Most of the gall-inducing species are extremely host-specific, developing only on a particular genus or species of plants and then only certain tissues during a restricted period of their development.
Two species of the genus Orseolia, feeding on the cultivated rice in Asia (Orseolia oryzae) Wood Mason, 1880 and in Africa (O. oryzivora) Harris and Gagne (1982) were confused for a long time. Harris and Gagne (1982) showed that these species were morphologically distinct. O. oryzivora is indigenous to Africa and is particularly widespread in West Africa (Dale, 1994).
This research reports studies on the anatomy of the reproductive system and the egg’s envelopes of O. oryzivora, in relation to oviposition site. The reproductive system was described and features related to the multiplication potential of the pest in the field. In addition, the ultrastructure of the newly-laid egg’s envelopes and its relation to oviposition site were explored by electron microscopy.
MATERIALS AND METHODS
Eggs and female adults of O. oryzivora were obtained from a culture of the insect reared on 25-30-day old seedlings of the released rice variety, FKR 28 under controlled conditions (90±2% RH, 30±1°C) (Nacro, 1994). The insect was reared in Plexiglas® cages (60x60x45 cm). Leaves bearing newly laid eggs (less than 24 h old) were removed from the seedlings of the rice plant. The prepared materials were observed with Photonic Microscope (PM), Scanning Electron Microscope (SEM) and Transmission Electron Microscope (TEM).
Photonic Microscopy: The Reproductive System of the Female of O. oryzivora
The study of the reproductive system of O. oryzivora was based on in toto histological preparations. The living female adults (aged less than 24 h) of the midge were killed by refrigerating them for a few min. They were then transferred to Ringer’s solution and dissected under a binocular microscope. The reproductive system was carefully removed and fixed in alcoholic Bouin on histological slide. The material was coloured according to Feulgen’s method, augmented when necessary by background staining with lacteous G methyl green. The material was then mounted between slide and covership in Canadian balsam.
Scanning Electron Microscope: The Laid Egg’s and its Envelopes (O. oryzivora)
Leaves bearing O. oryzivora’s eggs were cut off in order to keep just the attaching part of the eggs. O. oryzivora’s eggs with leaves attached were fixed and dehydrated in successive alcohol solutions (70, 80, 95 and 100%) and acetone solutions (50, 70, 90 and 100%). The material was then mounted on a lead object-holder. Samples were critical point dried in a Balzers CPD 010 apparatus with liquid CO2 gas then gold palladium coated with a JEOL JFC-100 sputter. These samples were observed under a JSM 6400 electron-scanning microscope.
Transmission Electron Microscope: Annex Glands and Egg’s Envelopes of O. oryzivora
The newly-laid eggs were removed from leaves and washed off into Ringer’s solution before fixed for 1 h in a 2.5% glutaraldehyde solution neutralized by solution cacodylate (0.1 M). After many rinses in the buffer, the material was progressively dehydrated in acetone solutions (50, 70, 90 and 100%) and was embedded in Araldite resin. The polymerisation took 48 to 72 h in a steam room at 58-60°C. Semi-fine sections stained with toluidine blue were prepared and photographed. Ultra-fine sections were made on collodioned grilles, contrasted, respectively by uranyl acetate for 2 h and citrate of lead for 10 min and shaped by carbon. These sections were observed under a JEOL 100 CX transmission electronic microscope.
RESULTS
The Ovaries and their Annex Glands
The reproductive system diameter (Fig. 1A) includes two sub-spherical and pro-ovogeny ovaries with 3.7 mm in length. Each ovary is formed by a large number of ovarioles (probably more than 20) appearing independent one from another. The ovary extends by a lateral oviduct (1.4 mm), which converges medially so that the two lateral oviducts emerge into a common median oviduct that is twice as long as the lateral ones (2.8 mm). The common oviduct extends to the vagina. At this point the ducts of the accessory glands and the spermathecae enter. The spermathecae is a filiform organ of 1.1 mm long. The accessory glands are 3.1 mm long and 1.0 mm large, ovoid, with a slight medial constriction (diameter: 0.86 mm). These glands show a secretory epithelium with variable thickness (9.5 to 14.2 μm) (Fig. 1B). The epithelium separates a reservoir from a draining canal and includes secretory grains (Fig. 1C), which secrete into the reservoir (Fig. 1B). The light of the reservoir is about 12.3 mm large.
The Egg Envelopes
Under scanning electron microscope, the egg layers include a thin regular exochorion (0.2 μm) and an endochorion (Fig. 1D). The internal surface of the exochorion is rough while the external surface is smooth (Fig. 1E). The exochorion detaches easily (Fig. 1F) from the underlying relatively thicker (0.5 μm) endochorion (Fig. 1G). The external surface of the endochorion is spotted with granules (diameter: 0.6 μm). The continuous vitellin membrane under the endochorion measures 0.02 μm in thickness while the discontinuous ovarian membrane observed just underneath measures 0.6 μm on its thickest portions. This group of envelopes re-covers the vitellus, which contains peripheral secretions.
The Laid-egg on the Plant
The laid-egg is oval and measures 700 μm long and 150 μm wide (Fig. 2A). The female adult lays its eggs in most cases singly or in small groups of two (Fig. 2B) to six eggs often connected by mucus (Fig. 2C), on the lower and upper surfaces of the rice leaf. On the leaf, the eggs are always laid parallel to the axis of the leaf, along the central vein, between two lines of siliceous phytoliths (Fig. 2D). These phytoliths are halter-like and measure 10 μm in length and 2 μm in thickness. There is 1.7 papillae per 100 μm2 between two rows of phytoliths. The papillae leave their impressions on the mucus when the eggs are removed (Fig. 2E). The impression of the leaf appears in function of the longitudinal axis of the egg (Fig. 2F). The eggs are attached to the plant surface by the mucus which covers them entirely and which the accessory glands secrete during oviposition. That mucus appears more abundant on the ventral surface of the egg, which is the part in contact with the plant. During the drying process, the mucus can form folds (length: 6 μm) seen in the sections (Fig. 2G).
DISCUSSION
The organization of the reproductive apparatus of the adult female of O. oryzivora is similar to that of other Diptera. There are also some similarities with the reproductive system of the cabbage fly, Delia radicum Linnaeus (Diptera, Anthomyiidae) (Neveu et al., 1997).
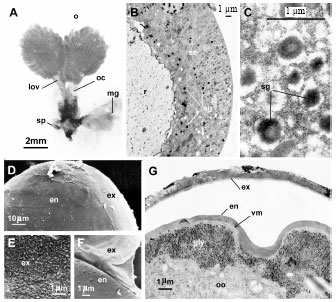 | |
| Fig. 1: | The reproductive system of the female adult of Orseolia oryzivora a: view of two voluminous ovaries and accessory glands; b: transversal section of an accessory gland showing a secretory epithelium which secretes into a voluminous reservoir; c a section of a mucus secretory cell; d: surface of a laid-egg showing the exochorion and the endochorion; e: smouth external surface of the exochorion; f: internal surface of the exochorion that was easily separated from the external surface of the endochorion; g: transversal section showing another view of the two separated envelopes of the egg (oc: oviduct canal; se: secretory epithelium; mg: mucus gland; lov: lateral oviduct; Ov: ovary; S: secretion grains; Sp: spermathecae; R: reservoir; ch: chorion; en: endochorion; ex: exochorion) |
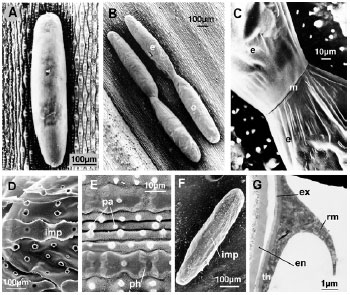 | |
| Fig. 2: | The laid-egg and its attachment to the plant a: an egg laid singly on the leaf of the rice plant; b: two groups of 2 laid-eggs each; c: two eggs joined by the mucus; d: impressions left on the rice leaf by a removed egg; e: impressions left on the mucus-covered egg by the phytoliths; f: two lines of phytoliths where eggs are always laid; g: the thickness of the mucus observed on the surface of the egg in contact with the rice plant (imp : impressions left on the plant by the removed egg m: mucus e: egg ph: phytolith th: thickness of the mucus) |
The dimensions of the laid-egg are closed to those of Bradysia aprica Winnertz (Diptera, Sciaridae), a species studied by Carcupino and Lucchi (1995) but the gall midge differs from Sciaridae in that its egg does not show any micropilar. Carcupino and Lucchi (1995) stressed the interest of the egg’s envelopes in oviparous animal groups, in that they protect the developing embryo from physical insult and dehydration.
The study of the oviposition sites of the midge reveals that the insect selects surprisingly precise zones on which to oviposit. The eggs are laid parallel to the leaf axis between two lines of siliceous phytoliths near the central vein. These phytoliths, apart from the interest that they represent in taxonomy (Kauffman et al., 1985; Guignard, 1987), botany (Le Cohu, 1973) and ecology (Gueguen et al., 1975; Cherouvier et al., 1975) could be of importance in facilitating the attachment of the eggs on the plant. Biémont et al. (1982) reported that Bruchidius atrolineatus Pic (Coleoptera, Bruchidae) laid its eggs on the stomata of the leguminous plant, Vigna ungiculata Walpers, 1970 after the mucus, which is produced during oviposition, has molted into these stomata. In the case of O. oryzivora, this mucus is secreted by the two accessory glands of the reproductive system of the female adult and is thicker on the ventral surface of the egg, which is in contact with the host plant. In some cases, eggs may be covered by excrement (Damman and Cappuccino, 1991) or protected by armor oil (Eisner et al., 1996). Female of many animal species have showed to prefer to lay their eggs in places, which increase the survival rates or growth opportunities for their offspring. They often respond to the differences in physical features of the habitat. For example, Ward et al. (1999) reported that the yellow dung fly, Scathophaga sternoraria Linneaus (Diptera, Scathophagidae) is clearly able to make a fine decision on the placement of its eggs. The female showed a strong preference for the hills and laid very few eggs on the points because eggs laid on the hills are likely to increase their chances of survival than those laid on the points. Studying the oviposition site selection by predatory midge, Aphidoletes aphidimiza Rondani (Diptera, Cecidomyiidae). Lucas and Brodeur (1999) concluded that by ovipositing within aphid colonies, the midge female not only provides food for neonate larvae, but also protects them against intraguild predators. The midge female preferably lays eggs in high trichomes density where predation by coccinellids is reduced. Trichomes are also believed to acting as physical barrier to predators by preventing egg detection and capture on plant substrate. Kanno and Harris (2000) came to similar conclusions in studying the influence of physical features of grass leaves on the placement of eggs within the plant by the Hessian fly, Mayetiola destructor Say (Diptera, Cecidomyiidae). The female Hessian fly always decides on the genotype, the physiological status of the plant and where she will lay her eggs on the plant. So, eggs were always laid on the leaf blade rather than the leaf sheath. On the leaf blades that comprise a single plant, the majority of the eggs were oviposited on the youngest leaves. Finally, within each leaf, more eggs were oviposited on the adaxial surface than on the abaxial surface. The authors concluded that M. destructor obtained information about the leaf surface through her tactile and/or kinaesthetic senses and used this information when making egg-laying decisions. Parrela and Thompson (1998) gave another example of oviposition site preference. These authors showed that the thornless honey locust tree’s gall midge, Dasineura gleditchiae Asten Sacken (Diptera, Cecidomyiidae) had a quite similar oviposition behavior to our findings in O. oryzivora. D. gleditchiae preferred to lay its eggs singly or in clusters along the rachis of marginal folds of unexpanded leaflets.
Studies of the sensory system (olfactory) of the gall midge are necessary to conclude on the relationship between the ultrastructure of the egg’s envelopes and the oviposition site. Meanwhile, these findings constitute a contribution to the knowledge of the biology and the ecology of this insect pest.
ACKNOWLEDGMENTS
We thank Mrs. Allo for the preparation of the material for observation, Jo Le Lannic for SEM, Jean-Paul Roland for TEM, Drs. Harris and E. A. Heinrichs for reviewing the first drafts of this manuscript.
REFERENCES
- Biemont, J.C., G. Chauvin and J.F. Germain, 1982. Loeuf de Bruchidius atrolineatus (Pic) et son syteme de fixation. Can. J. Zool., 60: 2610-2615.
CrossRefDirect Link - Cherouvier, A., A. Gueguen and J.C. Lefeuvre, 1975. Essai de determination des phytolithes siliceux des Graminees et des Cyperacees. Description, apres etude en microscopie electronique a balayage des principaux types de phytolithes rencontres. CR Acad. Sci. Paris, 281: 839-842.
Direct Link