
T. Ramasubramanian
Department of Entomology, Tamil Nadu Agricultural University, Coimbatore-641 003, Tamil Nadu, India
K. Ramaraju
Department of Entomology, Tamil Nadu Agricultural University, Coimbatore-641 003, Tamil Nadu, India
A. Regupathy
Department of Entomology, Tamil Nadu Agricultural University, Coimbatore-641 003, Tamil Nadu, India
Journal of Entomology
Year: 2005 | Volume: 2 | Issue: 1 | Page No.: 33-39







ABSTRACT
Tetranychus urticae is known to have a high tendency to develop resistance to acaricides among the mite species. It has been heavily exposed to acaricides among the acari and had developed resistance to dicofol, amitraz, organotins, propargite, pyrethroids, fenbutatin oxide, hexythiazox, clofentezine, abamectin and METI (Mitochondrial Electron Transport Inhibitors) acaricides fenazaquin, fenpyroximate, pyridaben and tebufenpyrad around the globe. The compilation and subsequent comparison of resistance data reported for different strains world wide is a complicated task because of the difference in bioassay methodology adopted, variation in the susceptibility of reference strains used to calculate the resistance ratio and stage of the mite (adult female/larvae) assayed for by the toxicologists. The establishment of baseline LC50 to new acaricides before widespread use may allow better monitoring of changes in susceptibility over time and can provide opportunity to detect resistance before the occurrence of field failure. Discriminating concentrations need to be determined for quick and reliable monitoring of resistance in the future. Genetically established resistance mechanisms in spider mites were similar to those found in insects (reduced penetration, target site insensitivity and enhanced metabolism). An ARM (Acaricide Resistance Management) package for T. urticae need to be developed that may guide for the mitigation of resistance. The magnitude of resistance and the mechanisms responsible for the acaricide resistance in T. urticae around the globe have been reviewed.
PDF Abstract XML References
How to cite this article
T. Ramasubramanian, K. Ramaraju and A. Regupathy, 2005. Acaricide Resistance in Tetranychus urticae Koch (Acari:Tetranychidae)-Global
Scenario. Journal of Entomology, 2: 33-39.
DOI: 10.3923/je.2005.33.39
URL: https://scialert.net/abstract/?doi=je.2005.33.39
DOI: 10.3923/je.2005.33.39
URL: https://scialert.net/abstract/?doi=je.2005.33.39
INTRODUCTION
The two-spotted spider mite, Tetranychus urticae Koch enjoys worldwide distribution[1-5] with more than 200 host plants[6]. The validity of T. urticae (the green form) and Tetranychus cinnabarinus (Boisduval) (the carmine form) as two separate species has been questioned by many authors[3]. Dupont[7] synonymized T. cinnabarinus with T. urticae due to the lack of reproductive isolation and production of fertile hybrids. T. urticae is one of the most important pest species responsible for significant yield losses in many horticultural, ornamental and agronomic crops around the globe[8]. Though, many non chemical control strategies are advocated under the IPM umbrella, still the farmers in developing countries like India mainly rely on chemical pesticides for the management of this pest due to unsatisfactory control from cultural and biocontrol methods[9]. Moreover, T. urticae can easily adapt to plant varieties that have been especially selected for resistance to it[10,11]. As mite control at present relies overwhelmingly on chemicals, attention must be given to prevent the loss of such potential, which is a key factor in IMM (Integrated Mite Management) programmes. Thus, every effort has to be made to prolong the effective livespan of currently available selective acaricides, especially those that fit into IMM programme. Such acaricides should be used only when it is absolutely necessary[12,13]. However, the ability of this mite to develop high levels of resistance to acaricides rapidly even after only a few applications[5,14,15] cannot be ruled out. Resistance to almost all the available groups of acaricides in T. urticae is known from different parts of the world[16-20]. Magnitude of resistance and the mechanisms responsible for the acaricide resistance in T. urticae around the globe have been reviewed here.
Magnitude of resistance around the globe: T. urticae is known to have a high tendency to develop resistance to acaricides among the mite species[21]. High reproductive potential and extremely short life cycle, combined with the frequent applications of acaricides usually required to contain the population below economic thresholds, facilitates rapid resistance development resulting in control failures[15,19]. The first failure in the chemical control of spider mite occurred in the early fifties when resistance against organophosphates such as ethyl parathion was detected in green houses in the United States and Europe[22]. Such control failures have been reported more recently to the newer compounds such as hexythiazox, clofentezine and abamectin[16,18,23-25]. T. urticae populations world wide had developed resistance to dicofol, amitraz, organotins, propargite, pyrethroids, fenbutatin oxide and METI (Mitochondrial Electron Transport Inhibitors) acaricides fenazaquin, fenpyroximate, pyridaben and tebufenpyrad[17,19-21,26-36]. The compilation and subsequent comparison of resistance data reported for different strains around the globe is a complicated task because of the difference in bioassay methodology adopted, [microimmersion bioassay[37]/leaf residual bioassay[33] /slide dip bioassay[38] / foliar spray bioassay[19], variation in the susceptibility of reference strains used to calculate the resistance ratio and stage of the mite (adult female /larvae) assayed for by the toxicologists.
The widespread acaricide resistance has been a major obstacle in the cost-effective integrated mite management programme in Korea[36]. The Korean population of T. urticae showed high levels of resistance to fenpyroximate (Resistance Ratio (RR), 182), dicofol (RR, 82) and pyridaben (RR, 78) but little or no tolerance to abamectin (RR, 6.5), fenpropathrin (RR, 9.1), propargite (RR, 6.5) and azocyclotin (RR, 5.4)[29].
In California, the susceptibility of T. urticae population varied greatly to abamectin with resistance ratios at LC95 ranging from 1 to 658 in a one day leaf residual bioassay. These populations had up to 77% survivorship at a discriminating concentration of 3 ppm[24]. T. urticae populations collected from green houses in California, Florida, the Canary Islands and Holland varied greatly in their susceptibility to abamectin with resistance ratios at LC95 ranging from 0.5 to 175 in leaf residual bioassay with reference to the abamectin susceptible Rutgers University strain. The Holland strain showed high levels of resistance to abamectin at diagnostic concentration of 3 ppm[25]. Comins[39] suggested that migration above a critical rate will greatly retard the development of resistance. Most of the nurseries sampled in California, Florida and the Canary Islands had extensive agricultural land areas with varied crops adjacent to the green houses. In comparison, the extensive green house production in Holland with little agricultural land in adjacent areas may not provide sufficient immigration of susceptible individuals to cause dilution of resistant genotypes. Reduced immigration might account for the faster development of resistance in the Holland strain[25].
Dicofol has been proved to be a selective acaricide and several IPM programmes have aimed to prolong its use in future through resistance monitoring and management[13,40,41]. A number of cases of resistance to dicofol were however, reported for the populations of T. urticae from various parts of the world between 1965 and 1985[42]. There were differences in resistance levels at LC95 ranging between 2.6 and 23.9 fold in topical bioassays or between 5.0 and 58.9 fold in residual bioassays of T. cinnabarinus collected from green houses in various districts of Antalya Province, Turkey. It was interesting to note that populations collected from typical green house area where almost no land was left for open field cultivation or wild plant species, showed lower resistance levels[33]. This is in contrast with the earlier report of Campos et al.[25], who suggested intensive green house production with little or no arable land in adjacent areas may provide insufficient immigration of susceptible individuals to cause dilution of resistant genotypes and consequently permit much more rapid development of resistance in T. urticae in green houses.
Discriminating concentration bioassay with field strains from France, Italy, California, Florida and Brazil revealed that T. urticae collected from roses in a Brazilian green house was multi-resistant and exhibited not more than 20-40% mortality when treated with discriminating doses of abamectin (0.14 mg a.i. L-1), chlorpyrifos (130 mg a.i. L-1), clofentazine (11 mg a.i. L-1), deltamethrin (16 mg a.i. L-1), hexythiazox (0.63 mg a.i. L-1) and pyridaben (0.48 mg a.i. L-1). All other strains were however, as susceptible as the reference strain[19]. The discriminating concentrations adopted were the LC95 values calculated from LDPM-relationship of T. urticae strain Wiesmoor (OP resistant strain) larvae.
Japanese strain of T. urticae exhibited a strong resistance to mitochondrial electron transport inhibitors. The resistance factors for pyridaben and fenpyroximate were 2000 and > 4000, respectively and thus represent the highest resistance factors yet published for these compounds in larvae of T. urticae[19]. The resistance was very stable and even after four years in the laboratory the strain did not revert, indicating its dominant nature and lack of association with developmental disadvantages or other fitness costs described for other strains[43].
Pattern of cross-resistance in acaricide selected strains: The time required for the development of resistance to a particular acaricide can vary considerably even within a single mite species on a particular crop. Selection for resistance to a certain acaricide may lead to cross-resistance to others of the same class or even multi-resistance covering a range of different chemical classes[19].
| Table 1: | LCPM relationship of acaricides to the standard German susceptible strain (GSS) of T. urtica |
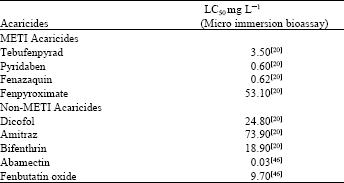 | |
Such cases have been reported for the thiazolidinone compound hexythiazox and the tetrazine acaricide clofentazine in T. urticae[23]. The knowledge of cross resistance pattern and biochemical mechanism conferring resistance to a particular compound have been shown to be necessary for resistance management tactics[44]. In order to provide sustained control of spider mite populations, it is becoming more and more important to design specific resistant management strategies[19], so as to protect the newer active ingredients from rapid resistance development within T. urticae populations[30,45] through cross-resistance.
A strain of T. urticae, collected from hops in England with a short history of tebufenpyrad exposure, exhibited resistance to four METI acaricides; tebufenpyrad, pyridaben, fenazaquin and fenpyroximate. Resistance factors for these compounds in a micro-immersion assay were 46, 346, 168 and 77, respectively with reference to the standard susceptible GSS strain (Table 1)[20,46] and corresponded to those exhibited by Japanese METI acaricide resistant strain (AKITA)[20]. This was the first published incidence of METI-acaricide resistance in Europe. It is clear that this magnitude of resistance seriously compromises the field efficacy of the METI-acaricides in UK.
The resistance patterns seen in this UK strain could have evolved under a number of different scenarios. First, the mechanism(s) imparting resistance to all four METI-acaricides could have arisen as a result of selection by tebufenpyrad in the locality from where the strain was collected[20]. Secondly, cross-resistance between tebufenpyrad and two fenpyroximate and pyridaben resistant strains selected from Korean population had already been reported[32]. In Western Australia, a field strain of T. urticae exhibited 63, 210 and 25 fold resistance to tebufenpyrad, pyridaben and fenpyroximate, respectively[31]. This strain had been exposed to tebufenpyrad applied five times over four seasons. The cross-resistance among these compounds can occur very rapidly, which have similar mode of action.
Korean population of T. urticae selected for resistance to fenpyroximate for 20 generations exhibited extremely strong positive cross-resistance to acrinathrin (RR, 196); high levels of cross-resistance to benzoximate (RR, 55) and propargite (RR, 64); moderate levels of cross-resistance to abamectin, fenbutatin oxide, fenpropathrin, pyridaben and tebufenpyrad but, low levels of cross resistance (RR, 16) to azocyclotin, bromopropylate, chlorfenapyr, dicofol, fenazaquin and milbemectin[36].
The METI-acaricides are potent xenobiotics that act largely by inhibiting complex I (NADH: ubiquinone oxidoreductase) of the mitochondrial respiratory chain[47]. Naphthoquinones, eg. BTG 505, BTG 514 and BTG 522 have recently been shown to have potential for mite control[48] and were reported to inhibit complex III of the mitochondrial respiratory chain[49]. This is distinct from the complex I site upon which the METIs act. METI-resistant strains of two-spotted spider mite from Japan (AKITA) and UK (TUK 4) exhibited no cross-resistance to the naphthoquinones[50].
Cross-resistance in T. urticae is often suggested to be the result of common metabolic detoxification mechanism (eg. dicofol-resistant T. urticae showed strong positive cross-resistance to amitraz). Fergusson-Kolmes et al.[27] evaluated cross-resistance between dicofol and 21 other chemicals. The dicofol resistant strain expressed a strong positive cross-resistance to the chlorinated biphenyl acaricides, chlorobenzilate and bromopropylate and a moderate positive cross-resistance to the formamidine acaricide, amitraz. However, chlorpyrifos was slightly more toxic to dicofol-resistant than to dicofol-susceptible spider mites. Increased oxidative activation of chlorpyrifos to chlorpyrifos oxon (a potent inhibitor of AChE) was responsible for the observed negative cross-resistance[51].
Spirodiclofen belongs to a novel class of acaricides newly discovered and developed by Bayer Crop Science and is currently in the registration process for the world wide use in the management of T. urticae. Spirodiclofen was not cross-resistant to chlorpyrifos, pyridaben, fenpyroximate, tebufenpyrad, hexythiazox, clofentazine and dicofol in the two spotted spider mite T. urticae.
Resistance monitoring with diagnostic concentration of 4 mg L-1 of spirodiclofen[52] revealed that the populations from Colombia were fully susceptible. Synergistic suppression studies with PBO, profenofos and DEM indicated that oxidative metabolism mediated by cytochrome P450 dependent monooxygenases seemed to play a major role in the spirodiclofen-selected strain[5].
Mechanism of acaricide resistance: Genetically established resistance mechanisms in spider mites were similar to those found in insects and consists of enhanced degradation of acaricides through esterases, glutathione S-transferases or cytochrome P450-dependent monooxygenases (metabolic resistance), a molecular change in the target site (altered acetylcholine esterase and nerve insensitivity) and finally, although less important, reduced penetration of acaricides through the mite cuticle[14,21].
Synergistic suppression studies and detoxifying enzyme assays in T. urticae strain selected with fenpyroximate revealed that enhanced activities of both mixed-function oxidases and esterases likely to contribute for the resistance. Pre treatment with PBO exhibited a remarkably higher level of synergism to fenpyroximate (synergistic ratio, 371.3) and it was suggested that PBO can be used as an effective synergist for the control of fenpyroximate resistant field populations of T. urticae[36]. Enhanced oxidative detoxification via MFO was found to be the major mechanism responsible for cross-resistance between acaricides in different strains of T. urticae[20,27,34,36].
T. urticae has been heavily exposed to acaricides among the acari and OP-resistant populations have been reported in more than 40 countries in both green house and open field crops[42]. In 1964 Smissaert[53] was the first who identified insensitivity of the target enzyme acetylcholinesterase as the major mechanism of resistance to OPs in spider mites. This was the first documentation of an altered target site in any arthropod pest species[15]. The most common mechanism of resistance to OPs in T. urticae was reduced sensitivity of the target enzyme acetylcholinesterase (AChE) to the inhibitory action of toxicants; this has been reported in T. urticae strains from several countries including the Nether lands, Germany, USA, New Zealand, Israel and Eygpt[16,53-61]. Another type of OP resistance reported in T. urticae was based on an increase in detoxification capacity resulting from increased carboxyl-esterase and phosphatase activities[62,63].
Mechanism of resistance to abamectin in T. urticae is polyfactorial. Biochemical studies carried out by Stumpf and Nauen[8] indicated the involvement of two major resistance mechanisms viz., increased metabolic detoxification by P450-dependent monooxygenases and glutathione S-transferases. However, actual metabolic studies with abamectin in spider mites need to be done to further verify that this compound is indeed metabolized by the suggested enzymes.
Investigation on the physiological mechanism of resistance in methyl parathion selected (23 generations, 44 fold) T.urticae revealed that resistance was not synergized by DEF (S,S,S-tributyl phosphorotrithioate) an inhibitor of esterases and glutathione S-transferases, or by PBO, an inhibitor of cytochrome P450 monooxygenases however, the AChE was less sensitive to inhibition by paraoxon[35].
Suggestions for the mitigation of acaricide resistance: An ARM (Acaricide Resistance Management) package for T. urticae need to be developed based on the earlier studies by keeping the below points in mind. This may guide for the mitigation of resistance.
| • | Regular monitoring should be carried out to detect the extent of resistance to the pesticides used[64]. Discriminating concentrations need to be determined for quick and reliable monitoring of resistance in the future. |
| • | Restricting the use of acaricides to which the magnitude of resistance is higher. |
| • | Potential acaricides having different metabolic routes may be recommended in rotation to reduce the selection pressure. |
| • | Acaricides showing positive cross-resistance can be avoided and those exhibiting negative cross-resistance may be encouraged. METI-resistant strain AKITA and hexythiazox-, clofentezine-and dicofol-resistant strain AU (collected from Australia) did not exhibit any cross-resistance to abamectin[8]. The studies of Beers et al.[18] also showed no cross resistance between abamectin and hexythiazox; so, these two compounds could be considered rotation partners in an overall acaricide resistance management programme[8]. |
| • | Potentiation mixtures should be considered only under certain special situations, provided there should be no cross-resistance between the products in the mixture. The mixtures may be avoided in small scale farming systems with low rate of literacy. If any pesticide mixture is used extensively there will be a danger of selection for resistance in pest insects. If there is any need arises, then mixtures can be considered for one time remedy[65]. |
| • | Possibilities of mitigating the resistance through myco-acaricides such as Beauveria bassiana (Balsamo) Vuillemin and Hirsutella thompsonii Fisher need to be explored. |
| • | Resistance risk analysis of new acaricides, which are under the process of registration, may reduce the chances of field failure. The establishment of baseline LC50 to new acaricides before widespread use may allow better monitoring of changes in susceptibility over time and can provide opportunity to detect resistance before the occurrence of field failure[18]. |
REFERENCES
- Van den Boom, C.E.M., T.A. van Beek and M. Dicke, 2003. Differences among plant species in acceptance by the spider mite Tetranychus urticae Koch. J. Applied Entomol., 127: 177-183.
CrossRefDirect Link - Devine, G.J., M. Barber and I. Denholm, 2001. Incidence and inheritance of resistance to METI‐acaricides in European strains of the two‐spotted spider mite (Tetranychus urticae) (Acari: Tetranychidae). Pest Manage. Sci., 57: 443-448.
CrossRefDirect Link - Tsagkarakou, A., N. Pasteur, A. Cuany, C. Chevillon and M. Navajas, 2002. Mechanisms of resistance to organophosphates in Tetranychus urticae (Acari: Tetranychidae) from Greece. Insect Biochem. Mol. Biol., 32: 417-424.
CrossRefDirect Link - Ramasubramanian, T., 2004. Magnitude, mechanism and management of pyrethroid resistance in Helicoverpa armigera Hubner in India. J. Entomol., 1: 6-11.
CrossRefDirect Link - Kim, Y.J., S.H. Lee, S.W. Lee and Y.J. Ahn, 2004. Fenpyroximate resistance in Tetranychus urticae (Acari: Tetranychidae): Cross-resistance and biochemical resistance mechanisms. Pest Manage. Sci., 60: 1001-1006.
Direct Link - Nauen, R., N. Stumpf and A. Elbert, 2000. Efficacy of BAJ 2740, a new acaricidal tetronic acid derivative against tetranychid spider mite species resistant to conventional acaricides. Proceedings of the Brighton Crop Protection Conference Pests and Diseases, Nov. 13-16, Brighton Hilton Metropole Hotel, UK., pp: 453-458.