
T. Ramasubramanian
Department of Entomology, Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu-641003, India
A. Regupathy
Department of Entomology, Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu-641003, India
Journal of Entomology
Year: 2004 | Volume: 1 | Issue: 1 | Page No.: 17-20







ABSTRACT
In Helicoverpa armigera, withdrawal of selection pressure for fourteen consecutive generations resulted in 2.58 and 3.01 fold increase in the susceptibility to lambdacyhalothrin and betacyfluthrin, respectively. Similarly, continuous selection enhanced the resistance level to the extent of 6.77 and 7.14 fold to the respective pyrethroids. Populations selected for resistance to lambdacyhalothrin and betacyfluthrin showed positive cross resistance to all other pyrethroids tested and no cross resistance to endosulfan. The increased level of mixed function oxidases with advancement of generation favoured the positive cross resistance among the pyrethroids.
PDF Abstract XML References
How to cite this article
T. Ramasubramanian and A. Regupathy, 2004. Pattern of Cross Resistance in Lambdacyhalothrin and Betacyfluthrin Selected
Populations of Helicoverpa armigera Hub. Journal of Entomology, 1: 17-20.
DOI: 10.3923/je.2004.17.20
URL: https://scialert.net/abstract/?doi=je.2004.17.20
DOI: 10.3923/je.2004.17.20
URL: https://scialert.net/abstract/?doi=je.2004.17.20
INTRODUCTION
The cotton bollworm, Helicoverpa armigera Hubner (Lepidoptera: Noctuidae) which was never traced as a major bollworm of cotton in any part of the country before 1986 has become number one agricultural pest in India. The crop loss due to this pest was estimated at 47–90%[1], the monitory value of which was more than Rs 2000 crores ($ 450 m)[2]. To combat the unprecedented pressure from H.armigera, farmers in southern peninsular region of India had applied over 30 sprays as against the recommended 8–10 sprays[1]. More than 75% of the insecticides used in cotton are being targeted towards H. armigera[3] of which, synthetic pyrethroids constitute 50-70%[4]. This high selection pressure for the past one decade slowly builds up the resistance in H.armigera and now it has climbed to a peak of more than 80% to synthetic pyrethroids in the Tamil Nadu State, India[5]. Low to high level of resistance had been detected in H. armigera to insecticides for which it had never been exposed earlier. This is due to the phenomenon of cross resistance. So, cross resistance in H.armigera is a major threat to present day intensive agriculture and is likely to remain an important aspect in the management strategies for the foreseeable future. The cross resistance pattern observed in lambdacyhalothrin and betacyfluthrin selected populations of H.armigera is discussed in this study.
MATERIALS AND METHODS
Development of resistant strains: H. armigera larvae collected from the Tamil Nadu State, India were reared on standard chickpea based semi synthetic diet[6] for fourteen consecutive generations. One set of laboratory-reared population was subjected to selection pressure with the synthetic pyrethroids to develop the respective pyrethroid resistant strains. The other set was maintained without exposure to any of the insecticide to serve as susceptible strain. The selection pressure was created by two different methods
| I | Discriminating doses (lambdacyhalothrin 0.025 μg and betacyfluthrin 0.2 μg-LD99 calibrated for susceptible strains in Australia[7]) were applied topically to create selection pressure in the first generation and the doses were gradually increased at the rate of 0.1 μg for each generation up to third. |
| II | The LD50 value arrived for F3 (unselected) generation was subsequently used to create selection in respective resistant populations from F4 onwards. |
The resistant strains were developed by retaining the survivors of the respective synthetic pyrethroids.
Insecticide bioassay: The pyrethroid resistant strains were topically treated with other insecticides of similar and different modes of action to assess the level of cross resistance in F9 and F14 generations.
Third instar larvae (30-40 mg) were used for bioassay. The required concentrations/discriminating doses prepared from technical grade insecticides were applied on the thoracic dorsum of each insect @ 1.0 μl using Hamilton repeating dispenser. The larvae were allowed to feed on the artificial diet. Minimum of 48 larvae were used per concentration. The treated larvae were kept at 25±2°C for 48 h when mortality was recorded. Larvae were considered dead if they were unable to move in a coordinated manner when prodded.
Enzyme activity in pyrethroid selected populations: The activity of mixed function oxidases (MFO) and carboxyl esterases (CE) was estimated in F2 and F12 generations. MFO assay was conducted following the method of Hansen and Hodgson[8]. Carboxyl esterase was assayed by following the method of Devonshire[9] and the protein estimation was done by the method of Bradford[10].
RESULTS
The LD50 of the insecticides to F1 generation of H.armigera was 3.63, 3.52, 2.39, 1.53, 1.02 and 6.36 μg larva-1 to fenvalerate, cypermethrin, deltamethrin, lambdacyhalothrin, betacyfluthrin and endosulfan, respectively. Continuous culturing of the population in laboratory without exposure to any insecticide resulted in decline of LD50 values to the extent of 2.58 and 3.01 fold to lambdacyhalothrin and betacyfluthrin, respectively when the population advanced to F14 (Table 1 and 2). Selection by pyrethroids initially with discriminating doses and subsequently with respective median lethal doses of F3 unselected population (1.37 and 0.76 μg to lambdacyhalothrin and betacyfluthrin) enhanced the level of resistance to 6.77 fold to lambdacyhalothrin and 7.14 fold to betacyfluthrin by the end of fourteenth generation (Table 1 and 2).
Population selected for resistance to one pyrethroid showed positive cross resistance to all other pyrethroids tested. The extent of cross resistance was 3.98, 3.14, 2.96 and 4.80 fold to fenvalerate, cypermethrin, deltamethrin and betacyfluthrin, respectively in F14 population selected by lambdacyhalothrin (Table 3). There was no cross resistance was observed to endosulfan (1.11 fold both in F9 and F14 generations). Betacyfluthrin selected population showed 2.47, 2.69, 3.42, 4.34 and 0.95 fold cross resistance to fenvalerate, cypermethrin, deltamethrin, lambdacyhalothrin and endosulfan, respectively (Table 4).
The MFO activity in lambdacyhalothrin selected second generation was 52.5 n mol min-1 mg of protein-1 and it was increased to 71.8 n mol min-1 mg of protein-1 when the generation advanced to F12. Similarly, the MFO activity increased to 68.5 n mol min-1 mg of protein-1 in F12 from 45.3 n mol min-1 mg of protein-1 in F2 in the population selected for resistance to betacyfluthrin. Carboxyl esterase activity in the F2 population selected for resistance to lambdacyhalothrin and betacyfluthrin was 385.3 and 323.5 n mol min-1 mg of protein-1, respectively. Successive selection with respective pyrethroids increased the carboxyl esterase activity with the advancement of generation (492.5 and 473.5 n mol min-1 mg of protein-1 in F12).
| Table 1: | Acute toxicity of lambdacyhalothrin to third instar H. armigera over generations |
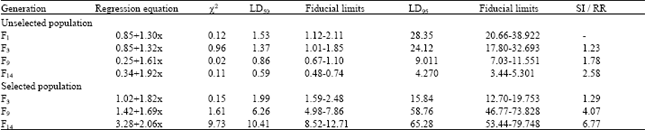 | |
| SI: Susceptibility index; RR: Resistance ratio | |
| Table 2: | Acute toxicity of betacyfluthrin to third instar H. armigera over generations |
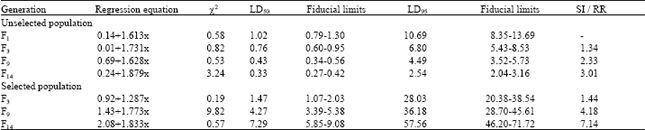 | |
| SI: Susceptibility index; RR: Resistance ratio | |
| Table 3: | Cross resistance pattern in lambdacyhalothrin selected population of H. armigera |
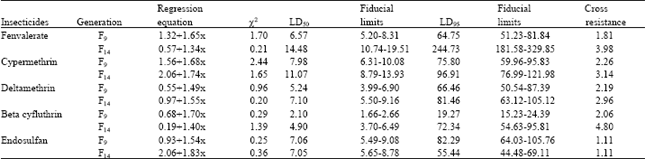 | |
| Table 4: | Cross resistance pattern in betacyfluthrin selected population of H. armigera |
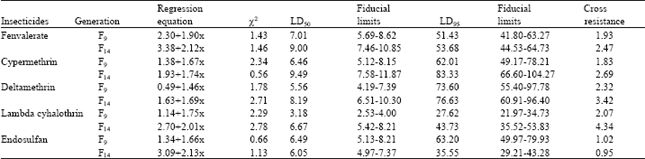 | |
DISCUSSION
Cross resistance is a potential problem that could limit the effectiveness of any insecticide. Information on development of cross resistance is important in formulating resistant management strategies. Furthermore, cross resistance would reveal information on the mechanism of action and metabolic pathways of insecticides. In the present investigation, the pattern of cross resistance studied in F9 and F14 generations revealed that the population selected for resistance to one pyrethroid extended cross resistance to other four pyrethroids tested.
Of the several types of reactions affecting the primary metabolism of pyrethroids, oxidation by MFO is of considerable importance and often plays a dominant role in determining the toxicity[11]. The trans and cis- methyl positions of acid moiety and 4 - phenyl position are the major sites in pyrethroids susceptible to oxidative metabolism[12]. The results of the monitoring studies conducted since 1993 indicated that the predominant mechanism of pyrethroid resistance in H. armigera populations from Tamil Nadu was by the induction of MFO as evidenced by the effective suppression of resistance by MFO inhibitors, piperonylbutoxide, propargyloxypthalimide and pungam oil[5,13-15]. Scott and Georghiou[16] had shown that MFO- mediated resistance is specific to pyrethroids having phenoxy-benzyl group. Since all the five synthetic pyrethroids detected for the level of cross resistance in the current investigation are ester bonded phenoxy-benzyl alcohols, the common MFO-mediated mechanism could be the reason for positive cross resistance observed among the pyrethroids. The enhanced level of MFO activity with the advancement of generation due to pyrethroids selection under laboratory condition also seems to support the above points. The present investigation clearly indicates that the enhanced metabolic degradation of synthetic pyrethroids due to MFO and to certain extent by carboxyl esterases favoured the cross resistance under laboratory selection. The development of cross resistance might be the reason for very high level of resistance to all synthetic pyrethroids observed[5] in H.armigera populations of Tamil Nadu State, India.
No cross resistance was observed to endosulfan in both the pyrethroid selected populations. The synthetic pyrethroids act principally on the voltage sensitive sodium channels[17-19] whereas the cyclodienes including endosulfan specifically attacks the picrotoxinin receptor site[20,21]. Thus, theoretically at least there should be no cross resistance between pyrethroids and endosulfan due to differential site of action. The present study in the laboratory also seems to support this.
ACKNOWLEDGMENT
The financial support from the Common Fund for Commodities (CFC), Europe, International Cotton Advisory Committee (ICAC), U.S.A. and Natural Resources Institute (NRI), U.K. is acknowledged.
REFERENCES
- Regupathy, A., D.S. Rajavel, S. Rajkumar and D. Russell, 1999. Present status of insecticide resistance in Helicoverpa armigera and its management in Tamil Nadu, India. Proceedings of the ICAC-CCRI Regional Consultation Insecticide Resistance Management in Cotton, Jun. 28-Jul. 1, CCRI, Multan, Pakistan, pp: 48-55.
- Ramasubramanian, T. and A. Regupathy, 2004. Magnitude and mechanism of insecticide resistance in Helicoverpa armigera Hub. population of Tamil Nadu, India. Asian J. Plant Sci., 3: 94-100.
CrossRefDirect Link - Forrester, N.W., M. Cahill, L.J. Bird and J.K. Layland, 1993. Management of pyrethroid and endosulfan resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) in Australia. Bull. Entomol. Res., 1: 1-132.
Direct Link - Hansen, L.G. and E. Hodgson, 1971. Biochemical characteristics of insect microsomes: N and O-demethylation. Biochem. Pharmacol., 20: 1569-1578.
CrossRefPubMedDirect Link - Devonshire, A.I., 1977. The properties of a carboxyl esterase from peach-potato aphid, Myzus persicae (Sulz.) and its role in conferring insecticide resistance. Biochem. J., 167: 675-683.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Regupathy, A., T. Manoharan, G. Ashokan, R. Nalini and N.J. Armes, 1995. Detoxification mechanisms involved in fenvalerate resistance in field population of Helicoverpa armigera Hub. in Tamil Nadu. In: Proceedings of the National Symposim IPM: An Entomological Approach to Sustainable Agriculture, pp: 22-24.
- Scott, J.G. and G.P. Georghiou, 1986. Mechanisms responsible for high levels of permethrin resistance in the housefly. Pestic. Sci., 17: 195-206.
Direct Link