
Rafia Aziz
Department of Gynecology, Government Medical College Baramulla, Baramulla, Jammu and Kashmir, India
Afak Yusuf Sherwani
Department of General Surgery, Government Medical College Baramulla, Baramulla, Jammu and Kashmir, India
Sameer Mohammad
King Abdullah International Medical Research Center, Department of Experimental Medicine, King Saud bin Abdulaziz University for Health Sciences (Ministry of National Guard Health Affairs), Riyadh 11426, Saudi Arabia
LiveDNA: 91.39026
ORCID: 0000-0002-1129-7685
Journal of Biological Sciences
Year: 2023 | Volume: 23 | Issue: 1 | Page No.: 1-9
DOI: 10.3923/jbs.2023.1.9







ABSTRACT
Background and Objective: Preeclampsia is one of the most dangerous hypertensive disorders that may arise during pregnancy and it is a major cause of maternal and fetal morbidity and mortality. Despite being a prominent topic of scientific inquiry, the fundamental pathways that contribute to the pathophysiology of preeclampsia remain little understood. Recent research suggested that G Protein-Coupled Receptors (GPCRs) may play a role in the etiology of preeclampsia, certain of these receptors have been observed to be elevated during a healthy pregnancy and decreased in preeclamptic people. However, the expression and functional importance of the great majority of GPCRs in the preeclamptic placenta are unclear. The goal of this study was to find highly expressed GPCRs that might play key roles in the physiology and pathology of the placenta. Materials and Methods: Dataset GSE148241 contained RNA sequencing data of 41 placentae in total, including from 9 patients with early-onset severe preeclampsia (EOSPE) and 32 normal controls. The RNA sequencing data were re-analyzed to evaluate the mRNA expression of the whole set of non-visual GPCRs in normal and preeclamptic human placenta. Results: The RNA sequencing identified more than 200 GPCRs in the healthy human placenta. Adhesion receptors (ADRGRG6 and ADGRG1), Atypical Chemokine Receptor 2 (ACKR2), Melanocortin Receptor 1 (MC1R), GPR107 and GPR78 are among the most abundant GPCRs. The expression of several GPCRs is significantly altered in the preeclamptic placenta. The list of GPCRs that are upregulated includes CCR5, HCAR2, GPR32, ADORA2A and GPR17 and those that are downregulated include ACKR1, SSTR1,OPRK1, FPR3 and CX3CR1. Conclusion: The GPCRs with altered expression in the preeclamptic may have substantial physiological importance in the placenta and may bring novel therapeutic options.
PDF Abstract XML References Citation
Copyright: © 2023. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Rafia Aziz, Afak Yusuf Sherwani and Sameer Mohammad, 2023. Analyzing Expression of GPCRs in Healthy and Preeclamptic Human Placenta. Journal of Biological Sciences, 23: 1-9.
DOI: 10.3923/jbs.2023.1.9
URL: https://scialert.net/abstract/?doi=jbs.2023.1.9
DOI: 10.3923/jbs.2023.1.9
URL: https://scialert.net/abstract/?doi=jbs.2023.1.9
INTRODUCTION
Preeclampsia (PE) is a serious illness that develops after 20 weeks of pregnancy and is defined by newly developed hypertension, proteinuria, uteroplacental insufficiency or damage to the mother’s organs1. Women with PE experience a wide range of indications and symptoms related to many organ systems, including headache, thrombocytopenia, chest tightness, pulmonary edema, reduced oxygen saturation, severe hypertension and reduced liver and kidney function. This disease is a leading cause of maternal and fetal morbidity and mortality2,3. Many research investigations have focused on the pathophysiological mechanisms of PE4,5. The placenta is the primary determinant in the development of PE. All maternal difficulties are characterized by a common pathophysiological component that focuses on placental anomalies3. Poor placentation and endothelial cell dysfunction all contribute to the etiology of PE. The molecular mechanism behind PE is still unknown and there has been little progress in its management due to the disease’s variability and the wide range of clinical symptoms associated with it5-7. The G Protein-Coupled Receptors (GPCRs), the largest class of membrane proteins in eukaryotes, regulate virtually all life processes and functions8. The GPCRs are activated by a wide variety of ligands or stimuli, including hormones, neurotransmitters and light, to regulate key physiological processes. The GPCRs are important factors in the onset and progression of various diseases and many marketed drugs exert their efficacy through GPCR signaling9,10. Exploratory studies have highlighted the critical roles of GPCRs in placental development and offered a theoretical foundation for their usage as prospective PE targets11,12. The GPCRs that have been studied for their role in placental development include Calcitonin Receptor (CALCR), Parathyroid Hormone 1 Receptor (PTH1R), Corticotropin-Releasing Hormone Receptor (CRHR1 and CRHR2), Glucagon Receptor (GCGR), Glucagon Like Peptide Receptor 1 (GLP1R) and adhesion receptors (ADGRA2, ADGRF5, ADGRL4, ADGRG6, ADGRE5 and ADGRG2)13-16. However, the expression and functional significance of a vast majority of GPCRs in placental biology are unclear. The RNA sequencing data was re-analyzed to assess the mRNA expression of the entire set of non-visual GPCRs. The RNA sequencing detected more than 200 GPCRs in the healthy human placenta and the expression of multiple GPCRs is significantly altered in the preeclamptic placenta. The GPCRs with altered expression in the diseased placenta may have significant physiological importance in the placenta and may provide novel treatment approaches.
MATERIALS AND METHODS
Study area: The study was carried out in Government Medical College Baramulla, Baramulla, Kashmir, India and King Abdullah International Medical Research Riyadh, Saudi Arabia.
Datasets: The RNA sequencing datasets GSE77085 and GSE148241 were used in this study. This dataset GSE77085 contained RNA sequencing data of villous tissue from 16 human placental samples. Third-trimester placenta samples taken from primiparous women with singleton pregnancies were used in this study17. Dataset GSE148241 contained RNA sequencing data of 41 placentae in total, including from 9 patients with early-onset severe preeclampsia (EOSPE) and 32 normal controls18.
RNA sequencing re-analysis: The RNA sequencing datasets GSE77085 and GSE148241were analyzed by the GEO RNA-seq Experiments Interactive Navigator (GREIN) platform19. The CPM-normalized mRNA expression was obtained from the GREIN platform for GSE77085 and the mean expression of each gene was determined. Dataset GSE148241 was analyzed on GREIN to determine differential gene expression between EOSPE and control placentae. Data were downloaded and the expression of the entire set of non-visual GPCRs (as per the International Union of Basic and Clinical Pharmacology Committee on Receptor Nomenclature and Drug Classification guidelines) was extracted TBTools software20,21.
Statistical analysis: The GPCR mRNA expression for normal placentae was expressed as Average±SEM. For comparison of mRNA expression between EOSPE and normal placentae, adjusted p-values (obtained from GREIN) were used and considered significant at p<0.01.
RESULTS
Landscape of the GPCRs in the human placenta: Expression of the entire repertoire of non-visual GPCRs in the human placenta was determined by reanalysis of the GSE77085 RNA sequencing dataset. The dataset contained RNA sequencing data from 16 human placentas and was available on the GEO RNA-seq experiments interactive navigator (GREIN) platform. Normalized gene expression data were downloaded from GREIN and data for all GPCRs were extracted using Tbtools (Fig. 1a).
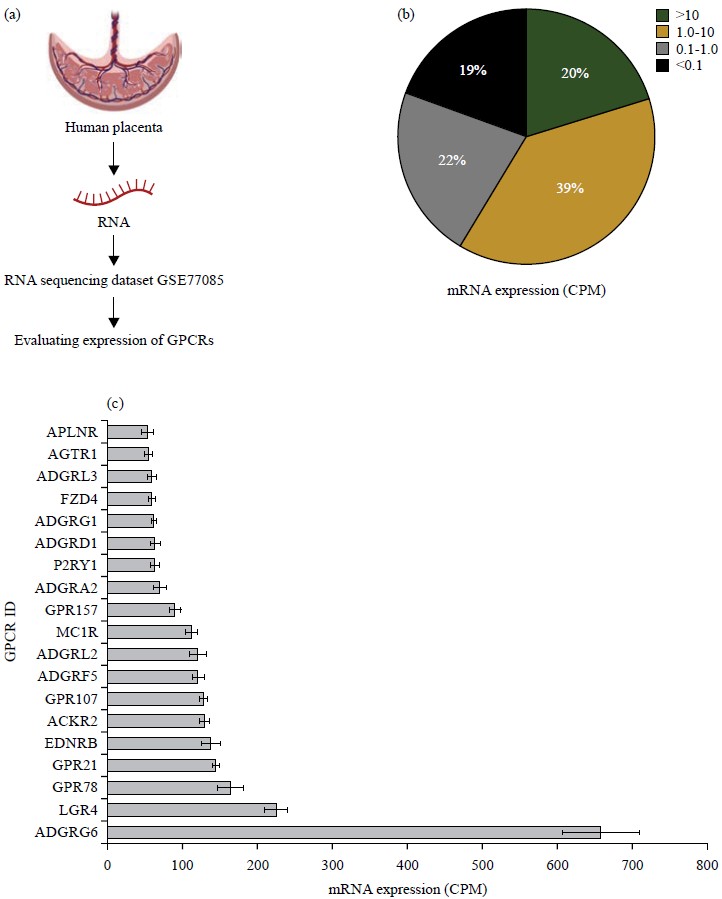 |
| Fig. 1(a-c): | mRNA expression profile of GPCRs in healthy human placenta, (a) Outline of the analysis, (b) Pi-chart showing the distribution of GPCRs based on their mRNA expression and (c) List of the top 20 highly expressed GPCRs Data was extracted from GSE77085 containing RNA sequencing data of 16 human placentae. The data for each GPCR is expressed as normalized CPM±SEM |
A total of 291 GPCRs were detected in the human placenta of which 73 were highly expressed (CPM>10), 139 were moderately expressed (CPM 1-10) and 79 were expressed at low levels (CPM 0.1-1) (Fig. 1b). Since 70 GPCRs had a CPM of 0.1 or were not detected, they were regarded as absent. The average expression of all the detected GPCRs was 14.2 CPM and 50 GPCRs were expressed at levels higher than the average expression (Table 1). Members of the adhesion receptor family (ADGRG6, ADGRF5, ADGRA2, ADGRD1, ADGRG1, ADGRL3 and ADGRl2), orphan receptors (GPR78, GPR21, GPR107 and GPR157), Endothelin Receptor
Type B (EDNRB), Atypical Chemokine Receptor 2 (ACKR2), Melanocortin Receptor 1 (MC1R), Purinergic Receptor P2Y1 (P2RY1), Frizzled Class Receptor 4 (FZD4), Angiotensin II Receptor Type 1 (AGTR1) and Apelin Receptor (APLNR) were among the highly expressed GPCRs in human placenta (Fig. 1c).
Impact of preeclampsia on GPCRs placental GPCR expression profile: Dataset GSE148241 contains RNA sequencing data from 41 human placentas, 32 from normal controls and 9 from patients with Early-Onset Severe Preeclampsia (EOSPE).
| Table 1: | List of GPCRs with higher levels of expression than the global average | |
| Gene symbol | Gene name | CPM |
| ADGRG6 | Adhesion G protein-coupled receptor G6 | 656.93 |
| LGR4 | Leucine rich repeat containing G-protein-coupled receptor 4 | 224.29 |
| GPR78 | G protein-coupled receptor 78 | 163.55 |
| GPR21 | G protein-coupled receptor 21 | 144.49 |
| EDNRB | Endothelin receptor type B | 136.99 |
| ACKR2 | Atypical chemokine receptor 2 | 129.56 |
| GPR107 | G protein-coupled receptor 119 | 127.96 |
| ADGRF5 | Adhesion G protein-coupled receptor F5 | 121.27 |
| ADGRL2 | Adhesion G protein-coupled receptor L2 | 120.39 |
| MC1R | Melanocortin 1 receptor | 111.90 |
| GPR157 | G protein-coupled receptor 157 | 90.07 |
| ADGRA2 | Adhesion G protein-coupled receptor A2 | 69.66 |
| P2RY1 | Purinergic receptor P2Y1 | 63.00 |
| ADGRD1 | Adhesion G protein-coupled receptor D1 | 62.95 |
| ADGRG1 | Adhesion G protein-coupled receptor G1 | 61.35 |
| FZD4 | Frizzled class receptor 4 | 59.23 |
| ADGRL3 | Adhesion G protein-coupled receptor L3 | 58.88 |
| AGTR1 | Angiotensin II receptor type 1 | 54.70 |
| APLNR | Apelin receptor | 53.36 |
| ADGRE5 | Adhesion G protein-coupled receptor E5 | 49.26 |
| OPN3 | Opsin 3 | 47.91 |
| ADGRA3 | Adhesion G protein-coupled receptor A3 | 42.24 |
| P2RY6 | Pyrimidinergic receptor P2Y6 | 39.97 |
| S1PR3 | Sphingosine-1-phosphate receptor 3 | 38.06 |
| S1PR1 | Sphingosine-1-phosphate receptor 1 | 37.08 |
| LPAR6 | Lysophosphatidic acid receptor 6 | 36.90 |
| ADRB1 | Adrenoceptor beta 1 | 33.05 |
| LGR5 | Leucine rich repeat containing G protein-coupled receptor 5 | 31.48 |
| FZD6 | Frizzled class receptor 6 | 26.64 |
| LPAR1 | Lysophosphatidic acid receptor 1 | 26.58 |
| CMKLR1 | Chemerin chemokine-like receptor 1 | 26.24 |
| CYSLTR1 | Cysteinyl leukotriene receptor 1 | 26.15 |
| FZD1 | Frizzled class receptor 1 | 25.48 |
| P2RY14 | Purinergic receptor P2Y14 | 24.76 |
| S1PR2 | Sphingosine-1-phosphate receptor 2 | 24.38 |
| TPRA1 | Transmembrane protein adipocyte associated 1 | 23.03 |
| GPR137 | G protein-coupled receptor 137 | 23.01 |
| GPR34 | G protein-coupled receptor 34 | 20.35 |
| CYSLTR2 | Cysteinyl leukotriene receptor 2 | 18.58 |
| ADGRF1 | Adhesion G protein-coupled receptor F1 | 18.08 |
| FZD7 | Frizzled class receptor 7 | 18.01 |
| ACKR3 | Atypical chemokine receptor 3 | 17.72 |
| SSTR2 | Somatostatin receptor 2 | 16.40 |
| FZD5 | Frizzled class receptor 5 | 16.29 |
| OPRK1 | Opioid receptor kappa 1 | 16.29 |
| C5AR2 | Complement C5a receptor 2 | 16.00 |
| CALCRL | Calcitonin receptor like receptor | 15.84 |
| ADGRL4 | Adhesion G protein-coupled receptor L4 | 15.40 |
| CXCR4 | C-X-C motif chemokine receptor 4 | 15.24 |
| FPR1 | Formyl peptide receptor 1 | 14.36 |
| Information is presented as normalized CPM (mean of 16 healthy human placentae from RNA sequencing dataset GSE77085) | ||
The data was processed using the GREIN platform and a list of differentially expressed genes between preeclamptic and normal placentae was generated (Fig. 2a). The data for all GPCRs were extracted using TBtools. Preeclampsia had a substantial effect on the expression of GPCRs and several members of this family show significant changes in their expression in preeclamptic placenta (Fig. 2b). Twenty two GPCRs were upregulated and 9 GPCRs were downregulated in the preeclamptic placenta (padj<0.01).
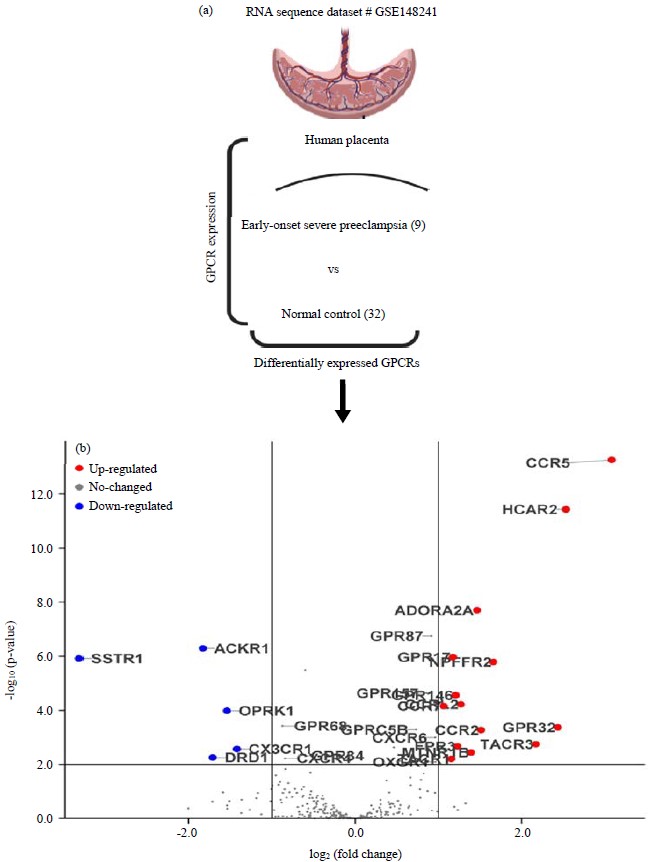 |
| Fig. 2(a-b): | Differential expression of GPCRs in preeclamptic placentae compared to normal healthy controls, (a) Framework of the analysis and (b) Volcano plot showing the differential expression of GPCRs between preeclamptic and normal placentae Differential expression of genes was obtained by re-analyzing RNA sequencing dataset # GSE148241, which contains data from placentae from 9 preeclampsia patients and 32 normal controls. The highlighted genes show a twofold or greater change and statistical significance (adjusted p<0.01) |
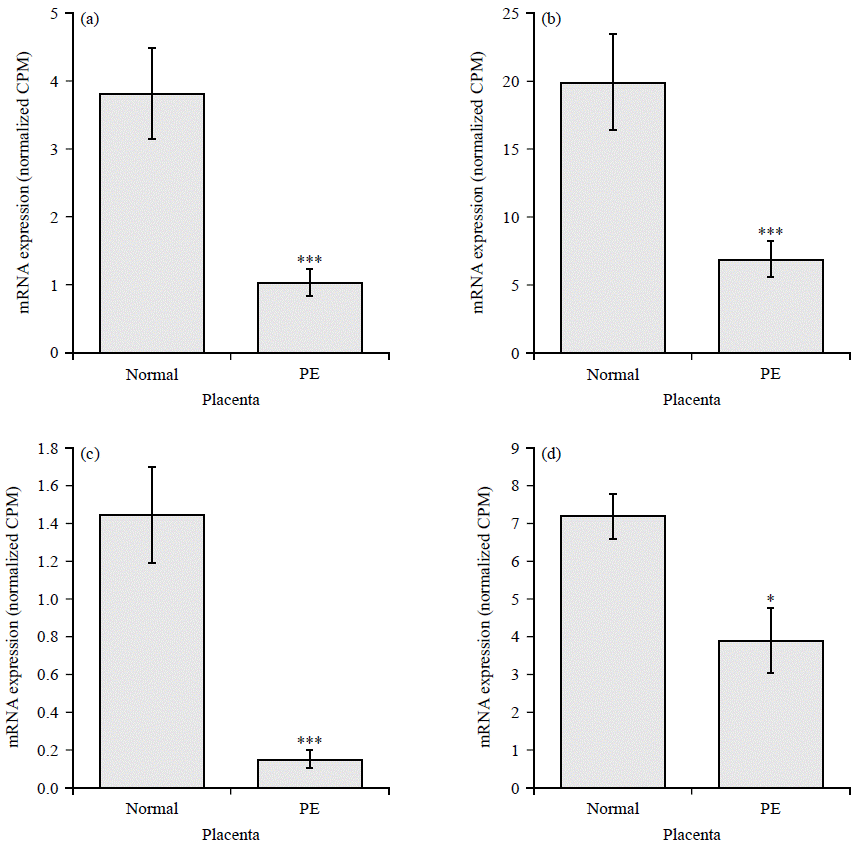
The highly upregulated GPCRs include Chemokine receptor (CCR5), Hydroxycarboxylic Acid Receptor 2 (HCAR2), uracil nucleotide/cysteinyl leukotriene receptor (GPR17), resolving receptor (GPR32), Melatonin Receptor 1B (MTNR1B) and adenosine A2A Receptor (ADORA2A) (Fig. 3a-f). Downregulated GPCRs include Atypical Chemokine Receptor 1 (ACKR1), Kappa Opioid Receptor (OPRK1), Somatostatin Receptor 1 (SSTR1), C-X-C Chemokine Receptor Type 5 (CXCR5), pH-Sensing G Protein-Coupled Receptor (GPR68) and C-X-C chemokine receptor type 4 (CXCR4) (Fig. 4a-f).
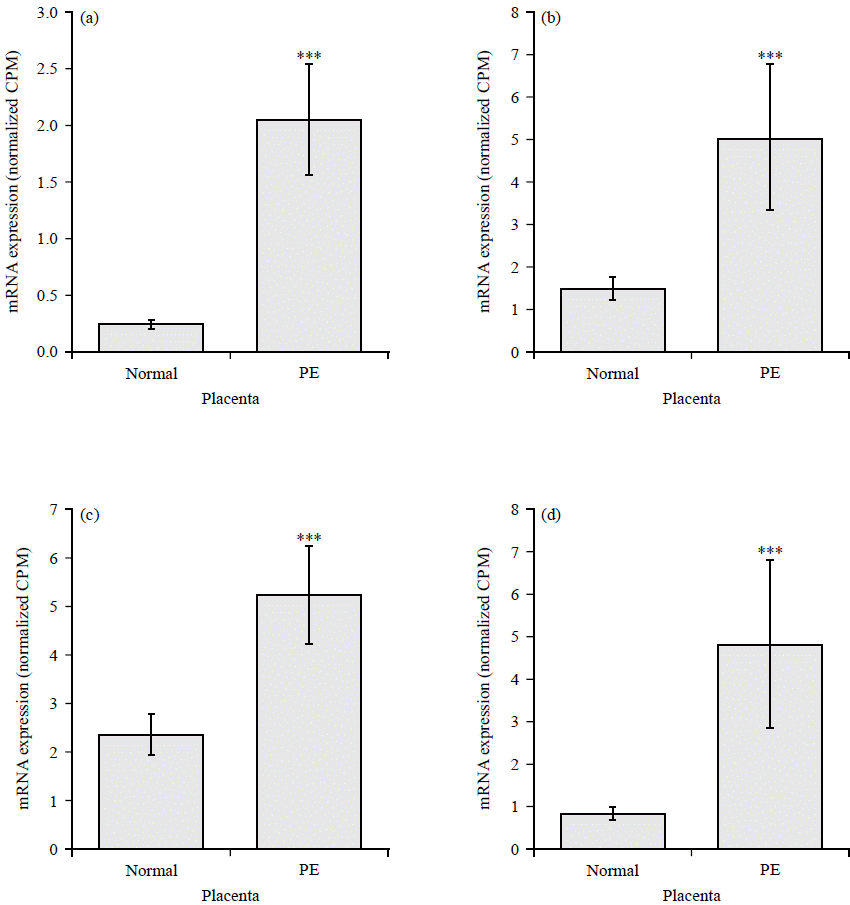 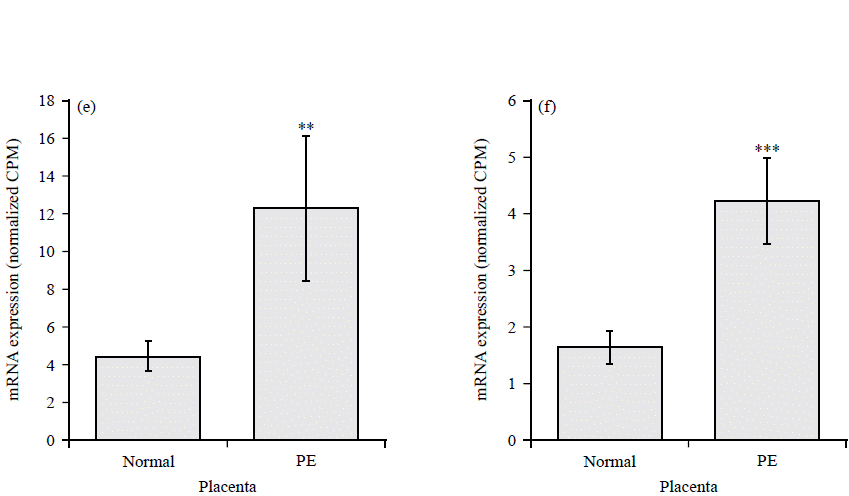 |
| Fig. 3(a-f): | GPCRs upregulated in preeclamptic placentae compared to normal healthy controls, (a) CCR5, (b) HCAR2, (c) GPR17, (d) GPR32, (e) MTNR1B and (f) ADORA2A Normalized CPM of indicated genes in placenta from normal (N = 32) and preeclampsia patients (N = 9), each bar represents the average CPM±SEM, adjusted p-values ***p<0.001 and **p<0.01 |
 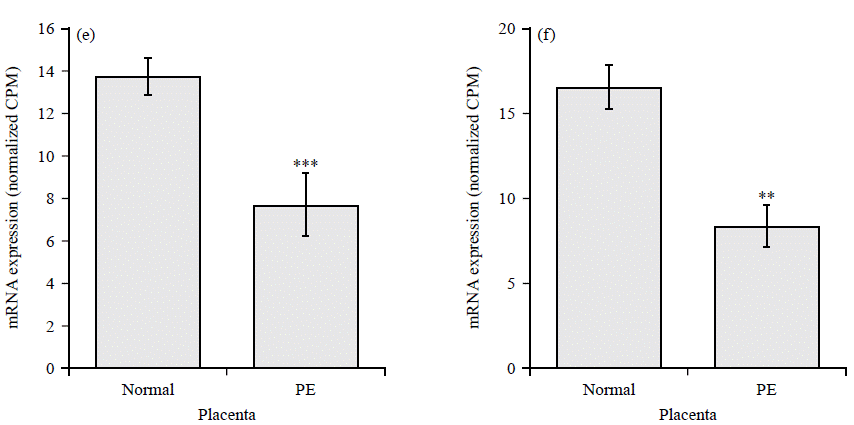 |
| Fig. 4(a-f): | GPCRs with reduced expression in preeclamptic placentae compared to normal healthy controls, (a) ACKR1, (b) OPRK1, (c) SSTR1, (d) CXCR5, (e) GPR68 and (f) CXCR4 Normalized CPM of indicated genes in placenta from normal (N = 32) and preeclampsia patients (N = 9), each bar represents the average CPM±SEM, adjusted p-values ***p<0.001, **p<0.01 and *p<0.05 |
DISCUSSION
Preeclampsia (PE) is a multisystem pregnancy condition that causes new-onset hypertension after 20 weeks of gestation and impairs several organ systems. The PE affects around 5-8% of pregnant women and is a major cause of maternal mortality4,5. Women with preeclampsia are more likely to experience subsequent health issues in addition to negative pregnancy outcomes. The immunological, genetic and environmental factors behind PE are still not completely known and there hasn’t been a significant advancement in PE therapy as a result of the heterogeneity of PE and the variety of its clinical presentations22,23. The GPCRs are a class of seven-transmembrane receptors that make up the largest group of cell surface receptors in mammalian cells. They control several crucial biological processes and abnormal GPCR signalling has been linked to the pathophysiology of several illnesses. The GPCRs are appealing therapeutic targets due to their unique combination of variation and specificity within the GPCR family and the fact that they are easily targetable by exogenous drugs. Several studies have shown the importance of GPCRs in placental development and provided a theoretical basis for their use as prospective PE targets11,24,25. However, the expression and functional importance of a vast majority of GPCRs in placental biology are unknown. This is in part, due to the scarcity of expression studies. Therefore, this study was initiated to comprehensively profile the entire repertoire of non-visual GPCRs by re-analyzing RNA-sequencing data. A total of 291 GPCRs were detected in the human placenta of which 73 were highly expressed (CPM>10). Many of the highly expressed GPCRs have no known role in the placenta. The expression of GPCRs was significantly impacted by preeclampsia and the mRNA expression of 22 GPCRs was elevated and 9 GPCRs were down regulated in the preeclamptic placenta as compared to the normal control.
Our in-depth analysis found many GPCRs with remarkable expression profiles, but only a small number of them have been investigated for their function in placental biology12. Many GPCRs with striking expression profiles in our study have no known role in the placenta and merit additional functional investigation. The Kappa Opioid Receptor (OPRK1), proton-sensing receptor (GPR68) and Atypical Chemokine Receptor 1 (ACKR1) are highly expressed in healthy human placenta markedly downregulated in preeclampsia suggesting a possible role in placental biology. Similarly, uracil nucleotide/cysteinyl leukotriene receptor (GPR17), resolvin receptor (GPR32) and Melatonin Receptor 1B (MTNR1B) are significantly upregulated in the preeclamptic placenta and have not been studied before. Insight into the functional significance of these recently discovered GPCRs may help in understanding disease pathophysiology and expedite the drug discovery process.
CONCLUSION
The current study identified the several highly expressed GPCRs with no known role in placental biology, whose expression was altered in preeclamptic human placenta. Understanding the functional significance of these newly identified GPCRs might assist in disease pathogenesis and offer novel therapeutic opportunities. One of the study’s weaknesses is that it used the whole placenta to profile GPCR expression. The placenta is a composite organ and contains many types of cells and as a result, single-cell RNA sequencing will be useful in distinguishing the cell-specific expression of GPCRs in healthy and sick situations. Furthermore, the study is based exclusively on mRNA levels, which may or may not indicate changes in protein levels. As a result, individual GPCR protein levels must be determined using immunoblotting and immunohistochemistry.
SIGNIFICANCE STATEMENT
The expression and function of the vast majority of GPCRs in placental biology are unknown. Re-analysis of RNA sequencing data was performed to examine the mRNA expression of the whole collection of non-visual GPCRs. This study found that the expression of numerous GPCRs was considerably altered in the preeclamptic placenta. The GPCRs with altered expression in diseased placentas may have significant physiological importance in the placenta and could lead to novel treatment approaches.
ACKNOWLEDGMENTS
The authors acknowledge the support for the use of GREIN and TBtools software for data analysis and data extraction. The authors also acknowledge King Abdullah International Medical Research Center (KAIMRC) for supporting this study through research grant RC16/158/R awarded to SM.
REFERENCES
- Brown, M.A., L.A. Magee, L.C. Kenny, S.A. Karumanchi and F.P. McCarthy et al., 2018. Hypertensive disorders of pregnancy: ISSHP classification, diagnosis, and management recommendations for international practice. Hypertension, 72: 24-43.
CrossRefDirect Link - Chappell, L.C., C.A. Cluver, J. Kingdom and S. Tong, 2021. Pre-eclampsia. Lancet, 398: 341-354.
CrossRefDirect Link - Burton, G.J., C.W. Redman, J.M. Roberts and A. Moffett, 2019. Pre-eclampsia: Pathophysiology and clinical implications. BMJ, Vol. 366.
CrossRefDirect Link - Say, L., D. Chou, A. Gemmill, Ö. Tunçalp and A.B. Moller et al., 2014. Global causes of maternal death: A who systematic analysis. Lancet Global Health, 2: e323-e333.
CrossRefDirect Link - Abalos, E., C. Cuesta, A.L. Grosso, D. Chou and L. Say, 2013. Global and regional estimates of preeclampsia and eclampsia: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol., 170: 1-7.
CrossRefDirect Link - Phipps, E.A., R. Thadhani, T. Benzing and S.A. Karumanchi, 2019. Pre-eclampsia: Pathogenesis, novel diagnostics and therapies. Nat. Rev. Nephrol., 15: 275-289.
CrossRefDirect Link - Rana, S., E. Lemoine, J.P. Granger, S.A. Karumanchi, 2019. Preeclampsia: Pathophysiology, challenges, and perspectives. Circ. Res., 124: 1094-1112.
CrossRefDirect Link - Pierce, K.L., R.T. Premont and R.J. Lefkowitz, 2002. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol., 3: 639-650.
CrossRefDirect Link - Sriram, K. and P.A. Insel, 2018. G protein-coupled receptors as targets for approved drugs: How many targets and how many drugs? Mol. Pharmacol., 93: 251-258.
CrossRefDirect Link - de Francesco, E.M., F. Sotgia, R.B. Clarke, M.P. Lisanti and M. Maggiolini, 2017. G protein-coupled receptors at the crossroad between physiologic and pathologic angiogenesis: Old paradigms and emerging concepts. Int. J. Mol. Sci. Vol. 18.
CrossRefDirect Link - West, C. and A.C. Hanyaloglu, 2015. Minireview: Spatial programming of G protein-coupled receptor activity: Decoding signaling in health and disease. Mol. Endocrinol., 29: 1095-1106.
CrossRefDirect Link - Yin, A., X. Guan, J.V. Zhang and J. Niu, 2022. Focusing on the role of secretin/adhesion (class b) g protein-coupled receptors in placental development and preeclampsia. Front. Cell Dev. Biol. Vol. 10.
CrossRefDirect Link - Makino, Y., K. Shibata, I. Makino, K. Kangawa and T. Kawarabayashi, 2001. Alteration of the adrenomedullin receptor components gene expression associated with the blood pressure in pregnancy-induced hypertension. J. Clin. Endocrinol. Metab., 86: 5079-5082.
CrossRefDirect Link - Karteris, E., A. Goumenou, E. Koumantakis, E.W. Hillhouse and D.K. Grammatopoulos, 2003. Reduced expression of corticotropin-releasing hormone receptor type-1α in human preeclamptic and growth-restricted placentas. J. Clin. Endocrinol. Metab., 88: 363-370.
CrossRefDirect Link - Younes, S.T., K.J. Maeda, J. Sasser and M.J. Ryan, 2020. The glucagon-like peptide 1 receptor agonist liraglutide attenuates placental ischemia-induced hypertension. Am. J. Physiol. Heart Circ. Physiol., 318: H72-H77.
CrossRefDirect Link - Buckberry, S., T. Bianco-Miotto, S.J. Bent, V. Clifton and C. Shoubridge et al., 2017. Placental transcriptome co-expression analysis reveals conserved regulatory programs across gestation. BMC Genomics, Vol. 18.
CrossRefDirect Link - Majewska, M., A. Lipka, L. Paukszto, J.P. Jastrzebski and K. Szeszko et al., 2019. Placenta transcriptome profiling in intrauterine growth restriction (IUGR). Int. J. Mol. Sci. Vol. 20.
CrossRefDirect Link - Yang, X., J. Yang, X. Liang, Q. Chen and S. Jiang et al., 2020. Landscape of dysregulated placental RNA editing associated with preeclampsia. Hypertens., 75: 1532-1541.
CrossRefDirect Link - Al Mahi, N., M.F. Najafabadi, M. Pilarczyk, M. Kouril and M. Medvedovic, 2019. GREIN: An interactive web platform for re-analyzing GEO RNA-seq data. Sci. Rep. Vol. 9.
CrossRefDirect Link - Alexander, S.P.H., A. Christopoulos, A.P. Davenport, E. Kelly and A. Mathie et al., 2021. The concise guide to pharmacology 2021/22: G protein-coupled receptors. Br. J. Pharmacol., 178: S27-S156.
CrossRefDirect Link - Chen, C., H. Chen, Y. Zhang, H.R. Thomas and M.H. Frank et al., 2020. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant, 13: 1194-1202.
CrossRefDirect Link - Hod, T., A.S. Cerdeira and S.A. Karumanchi, 2015. Molecular mechanisms of preeclampsia. Cold Spring Harbor Perspect. Med., Vol. 5.
CrossRefDirect Link - Huppertz, B., 2020. Molecular and cellular mechanisms of preeclampsia. Int. J. Mol. Sci. Vol. 21.
CrossRefDirect Link - Conrad, K.P., 2016. G-protein-coupled receptors as potential drug candidates in preeclampsia: Targeting the relaxin/insulin-like family peptide receptor 1 for treatment and prevention. Hum. Reprod. Update, 22: 647-664.
CrossRefDirect Link - Vidal, J.M.S., C.D.T. Deguit, G.F.B. Yu and M.D. Amosco, 2022. G-protein coupled receptor dysregulation may play roles in severe preeclampsia-A weighted gene correlation network analysis of placental gene expression profile. Cells, Vol. 11.
CrossRefDirect Link