
Mohamed Abdel-Raouf
Gastroenterology Surgical Center, Mansoura University, Mansoura, Egypt
LiveDNA: 20.31282
Hisham M. Aldeweik
Department of Mcirobiology, Environmental monitoring Administration, Ministry of Health, Mansoura, Egypt
Ehab Y. Elbendary
Medical Laboratory Technology Department, College of Applied Medical Science, Jazan University, Kingdom of Saudi Arabia
LiveDNA: 20.31196
Journal of Biological Sciences
Year: 2020 | Volume: 20 | Issue: 1 | Page No.: 48-55







ABSTRACT
Background and Objective: L- Asparaginase is one of the amidohydrolases enzymes which catalyses the deamination of asparaginine to aspartic acid. Our study aimed to produce and purify L-asparaginase enzyme from soil bacteria and investigate the physical and chemical factors that effect on the amount of produced L-asparaginase enzymes. Materials and Methods: Fifty soil samples were collected from soil surface, the soil suspension was then inoculated on selective nutrient agar medium, Isolated bacterial colonies were identified by biochemical tests to select the most active bacterial strains for production of L-asparaginase enzyme. The L-asparaginase enzyme was purified by Sephadex G-100 column. The effects of temperature, pH, different nutritional factors were tested on the amount of produced L-asparaginase enzyme. Results: Erwinia carotovora has the highest L-asparaginase enzyme activity with 14 mm of intense pink zone, followed by E. coli (13 mm) while Streptococcus pyogenes has (12 mm). Optimal conditions for L-asparaginase enzyme production from Erwinia carotovora were pH 7.0 at 37°C. The best carbon source was glucose with activity (2.7) and best concentration of glucose was 10 g L–1. The best nitrogen source was L-asparagine by concentration of 5 g L–1 which gave activity (4.7). Finally this study indicated that the maximum L-asparaginase activity was obtained after incubation for 10 min at 37°C, pH 8.0 with adding 1.0 mL of magnesium chloride (MgCl2) as activator. Conclusion: Our study concluded that the soil can be used for obtaining L-asparaginase produced bacteria and the Erwinia carotovora is consider as the most active bacteria for L-asparaginase production.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mohamed Abdel-Raouf, Hisham M. Aldeweik and Ehab Y. Elbendary, 2020. Impact of Physical and Chemical Factors on L-asparaginase Enzyme Purified from Soil Bacteria. Journal of Biological Sciences, 20: 48-55.
DOI: 10.3923/jbs.2020.48.55
URL: https://scialert.net/abstract/?doi=jbs.2020.48.55
DOI: 10.3923/jbs.2020.48.55
URL: https://scialert.net/abstract/?doi=jbs.2020.48.55
INTRODUCTION
L-asparaginase belongs to a group of homologous amidohydrolases family, which catalyses the hydrolysis of amino acid L-asparagine (amide of 2-aminosuccinic acid), to aspartic acid and ammonia1. They are naturally occurring enzymes expressed and produced via animal tissues, bacteria, plants and in the serum of certain rodents, however it is absent in human2. Large quantity of microorganisms that consist of Erwinia carotovora, Pseudomonas stutzeri, Pseudomonas aerugenosa and E. coli has been recognized to produce L-asparaginase3.
The presence of asparagine is indispensable for a regular path of the cell cycle. In most human cells the deficit of the endogenous asparagine may additionally be compensated by way of alternative synthesis from aspartic and glutamic acid involving the asparagine syntheses. L-asparaginase enzyme is absent in sure neoplasmatic cells, particularly in lymphoblasts, for this reason the depletion of asparagine from the plasma leads to DNA, RNA and protein synthesis inhibition and in consequence to blast cell apoptosis1.
Different sorts of asparaginase can be used for pharmaceutical enterprise as an alternative for treatment of exclusive cancers such as acute lymphoblastic leukemia (ALL), malignant ailments of the lymphoid system and Hodgkin’s lymphomas4.
Cancer cells, often lymphatic cells, require high quantity of asparagine for fast and malignant growth. In this way, cancer cells require the amino acid from diet (blood serum) as well as amino acids produced by way of themselves. However, leukemic lymphoblasts and some others tumor cells do no longer have or existing low volume of L-asparagine synthetase used for L-asparagine synthesis. Thus, these malignant cells are dependent of asparagine from blood serum for their proliferation and survival5. L-asparaginase hydrolyzes asparagine from blood serum, leading tumor cells to dying by means of lacking of critical thing for protein synthetases (p53-dependent apoptosis). However, healthful cells are now not affected, because they are capable to produce asparagine, the use of L-asparagine synthetase existing in enough quantities6. The soil metagenome derived L-asparaginase with enhanced activities could be a potential candidate to develop as a drug in ALL therapy7. Also, El-naggar et al.8, concluded that the L-asparaginase enzyme can be used for development of chemotherapeutic drug. L-Asparaginase (L-asparagine aminohydrolase, E.C. 3.5.1.1) has been proven to be competent in treating ALL, which is widely observed in pediatric and adult groups9.
Also, this enzyme is used in food industry to prevent the acrylamide formation when ingredients are processed in excessive temperatures10. Several researches show that L-asparagine is the primary amino acid responsible for acrylamide production in fried and baked meals when reducing sugars are condensed with a carbonyl source. This phenomenon does now not manifest in boiled food11. Many research confirmed that the use of L-asparaginase enzyme earlier than frying or baking food manner should decrease greater than 99% acrylamide degree in the processed final product. This is due to the fact the enzyme reduces extra than 88% of the L-asparagine concentration from the initial feedstock12. This use is essential because acrylamide is a neurotoxin categorized as potentially carcinogenic to humans13.
L-asparaginases are enzymes derived mainly from bacteria and the three enzymes that have been used are derived from E. coli (E. coli-asparaginase), a pegylated form of native E. coli-asparaginase (PEG-asparaginase) and an Erwinia chrysanthemi-derived asparaginase (Erwinia-asparaginase)14.
The purpose it is preferred for the reason is it is biodegradable, non-toxic and can be administered at the nearby site quite easily. Other sellers are observed pretty painful when administered to the patient and additionally these are pretty costly. Current clinical studies point out that this enzyme is additionally a promising agent in treating some varieties of neoplastic cell diseases in human15.
Industrial L-asparaginase production offers some challenges, such as the search for new microorganisms in a position to produce it with much less unfavorable effects. Nowadays, industrial manufacturing is carried out using bacteria such as E. coli and Erwinia chrysanthemi, fungi and yeast. Currently, new studies have been carried out aiming to increase manufacturing process and set up new approaches for enzyme synthesis6. This study aimed to detect and purify L-asparaginase enzymes from soil bacteria and study the effect of different physical and chemical factors on the enzyme activity.
MATERIALS AND METHODS
Study area: This study was carried out in Gastroenterology Surgical Center, Mansoura University, Egypt during the period February, 2017 till August, 2019.
Collection and preparation of soil samples: Fifty soil samples were collected from soil surface and depth of approximately 20 cm (around the plant roots) of cultivated potato field. The samples were taken in sterilized screw capped cup using sterilized spatula different soil samples were passed through a sieve (1.7 mm mesh) to remove large pieces of debris and vegetation. One gram of the soil samples were added to 10 mL of water to make soil suspension16.
Inoculation: The soil suspension was then inoculated on selective nutrient agar medium supplemented with antifungal agents (Fluconazole 75, Ketoconazole 75 μg mL–1) to control the fungal contamination. The inoculated agar plates were incubated at 37°C for 24 h.
Bacterial Identification and selection: Isolated bacterial colonies were identified by biochemical tests(coagulase, starch hydrolysis, oxidase, urease, catalase andindole). To select the most active bacterial strains for production of L-asparaginase enzyme the identified and purified isolated bacteria were point inoculated on agar-based modified medium (composition (g L–1): KH2PO4 2.0, L-Asparaginase 6.0, MgSO4.7H2O 1.0, CaCl2.2H2O 1.0 and Glucose 3.0) supplemented with 0.004% phenol red dye (prepared in ethanol) and pH was adjust at 6.1. The inoculated plates were incubated at 37°C for overnight. Diameter zone and appearance the pink color indicated that L-asparaginase production16.
L-asparaginase purification: The ammonium sulphate precipitate was applied to a Sephadex G-100 column. Seven grams of Sephadex G-100 were equilibrated with 0.05 M borate buffer (pH 7.5) in water bath at 60°C for 3 h. The gel was then cooled to room temperature. A glass column 1.0×38 cm was packed with swollen gel and equilibrated with the same buffer. Fractions after ammonium sulfate precipitation were subjected to gel filtration column. The protein elution was done with the same buffer at a flow rate of 2 mL every 10 min. The fractions were collected using a fraction collector (LKB Ultorace) at 40°C. It was assayed for protein content at 595 nm as well as for enzyme activity. Preparation of crude enzyme solution by inoculated a loop full of each isolated bacteria in 100 mL of broth-based modified medium. All flasks were incubated under shaking condition (150 rpm) for 2 days at 30°C. After incubation period, the cultures were filtered, then the filtrates were centrifuged at 5000 rpm for 10 min the clarified filtrate represented crude enzyme of L-asparaginase17.
Characterization of L-asparaginase enzyme: Effect of temperature on production of L-asparaginase enzyme was determined by incubation the flasks at different temperatures (25, 30, 37, 40 and 50°C) for 2 days18. The influence of pH on the enzyme activity was done by using 0.1 M of sodium acetate and 0.1 M of phosphate buffer at different pH levels (5, 6, 7, 8 and 9). The inoculated flasks were incubated for 2 days at 30°C and shacked at 150 rpm19. The effect of nutritional factors on L-asparaginase production, was studied by adding different carbon sources (Glucose, maltose, fructose, lactose, galactose and raffinose) and different nitrogen source (L-asparagine, urea, ammonium sulphate, ammonium chloride, ammonium nitrate and sodium nitrate) to flasks, then incubated for 2 days at 30°C and shacked at 150 rpm20.
The impact of different concentrations of glucose as carbon source on enzyme production was determined by preparing different concentrations from glucose (2, 4, 6, 8 and 10 g L–1) as well as, preparation of different concentrations of L-asparagine as nitrogen source (2.5, 5, 7.5 and 10 g L–1), then flasks were incubated for 2 days at 30°C and shacked at 150 rpm20.
The effect of different incubation periods on L-asparaginase enzyme production was analyzed by preparing mixture consisting of 0.5 mL of 0.08 M of L-asparagine, 1 mL of 0.05 M of borate buffer and 0.5 mL of purified enzyme, then incubated at 37°C for different period times (10, 20, 30 and 40 min).
The impact of different temperatures and pH on enzyme production was studied by incubated the enzyme mixture at 25, 30, 37, 40 and 50°C for 10 min and different pH (5, 6, 7, 8 and 9) at 37°C19. The effect of the amounts of MgCl2 as activator on enzyme activity was tested by preparing several amounts (0.2, 0.4, 0.6, 0.8 and 1.0 mL) of activator, then incubated at 37°C for 10 min21.
RESULTS
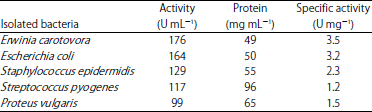
The enzyme production was determined by measuring the diameter zone of L-asparaginase producing bacteria. Erwinia carotovora has the highest (14 mm) intense pink zone, followed by isolate E. coli (13 mm) while isolate Streptococcus pyogenes has (12 mm) pink zone as shown in Table 1.
All the L-asparaginase-positive isolated bacteria were subjected to quantitative screening using CARY 60 spectrophotometer. The results in Table 1 and 2 revealed that Erwinia carotovora showed the highest activity for L-asparaginase production, so we selected for further study.
The results of environmental and nutritional factors affecting on production of L-asparaginase from Erwinia carotovora show in Fig. 1. The first environmental factor is incubation temp. This indicated that the maximum enzyme activity (150 U mL–1) and specific activity (2.7 U mg–1) occur at 37°C.
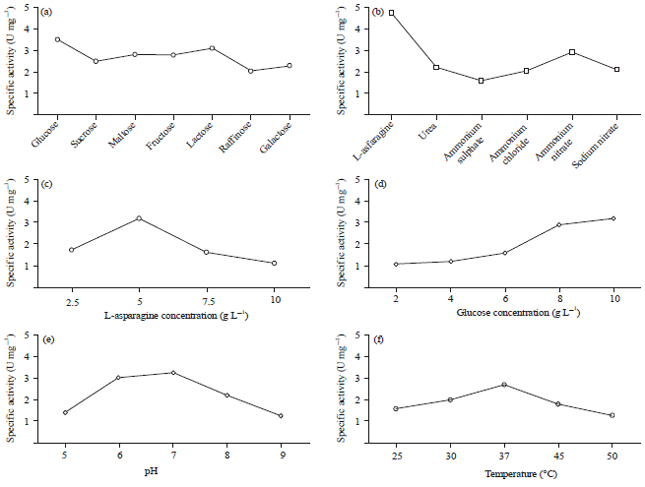 | |
| Fig. 1(a-f): | Specific enzyme activities were measured at different nutrients, (a) Different carbon sources, (b) Different nitrogenous sources, (c) L-asparagine concentrations, (d) Glucose concentrations, (e) pH and (f) Temperatures |
| Table 1: | Diameter zone of L-asparaginase producing bacteria |
 | |
| Table 2: | Quantitative screening for L-asparaginase activity of isolated bacteria in liquid medium |
 | |
Regarding to the second environmental factor it was initial pH value, which showed that the maximum enzyme activity (206 U mL–1) and specific activity (3.25 U mg–1) occur at pH 7. The first nutritional factor is carbon source, a goal of this experiment is study the effect of changing the carbon source on L-asparaginase production. It was found that glucose as the best carbon source for L-asparaginase production with enzyme activity (200 U mL–1) and specific activity (3.5 U mg–1). The second nutritional factor is nitrogen sources, a goal of this experiment is to study the effect of changing the nitrogen source on L-asparaginase production. It was found that L-asparagine is the best nitrogen source for L-asparaginase production with enzyme activity (127 U mL–1) and specific activity (4.7 U mg–1). The optimum glucose conc. was 10 g L–1, which have the highest enzyme activity (169.5 U mL–1) and specific activity (3.2 U mg–1). However, the optimum nitrogen conc. was 5.0 g L–1, which have the highest enzyme activity (170 U mL–1) and specific activity (3.19 U mg–1).
Figure 2 shown that the optimum incubation time, temp. and pH of partially purified activities and the rate of substrate hydrolysis was investigated by incubating the enzyme for with the corresponding substrate at optimum conditions. The maximum enzyme activity and specific activity were obtained after incubation for 10 min at 37°C and pH 8. Enzyme activity at different activator concentrations was investigated, the optimum enzyme activity and specific activity were obtained by addition of 1 mL magnesium chloride (MgCl2).
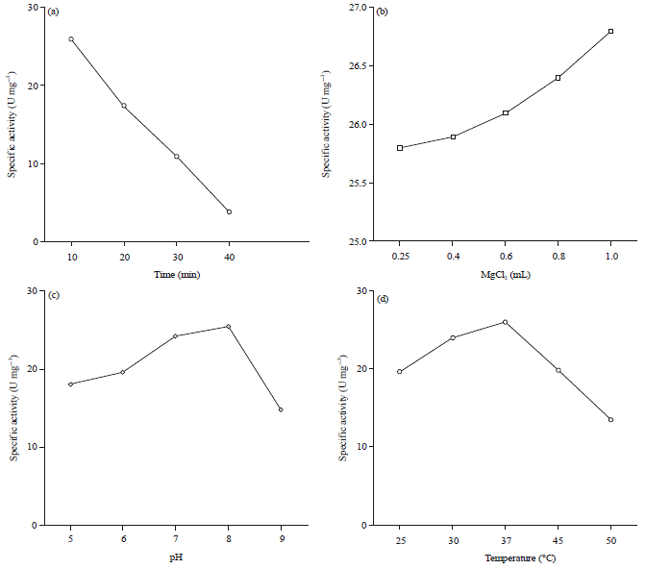 | |
| Fig. 2(a-d): | Specific enzyme activities were measured at (a) Different times, (b) Magnesium chloride concentrations, (c) pH and (d) Temperatures |
DISCUSSION
The present investigation included selection of the most potent bacterial isolate that capable of producing L-asparaginase enzyme. Then, increase productivity of L-asparaginase enzyme by studying the best factors affecting L-asparaginase production. In addition, attempts for purification and characterization of L-asparaginase were also investigated.
Five asparaginase producing bacteria were isolated from fifty soil samples collected from cultivated potato field. This may be attributed to the fact that soil is a rich source for potential enzyme producing organisms.
In this study, the result of screening test revealed that L-asparaginase producing bacteria were able to hydrolyze L-asparagine since they utilized L-asparagine as their substrate and breakdown asparagine to L-aspartate and ammonia which further reacts with water to produce NH4OH, hence the pH of the medium is basic which subsequently changes the medium from yellow to pink, hence the pinkish zone observed around the colony of asparaginase producing bacteria. This is in accordance with the work of Gulati et al.22 who proved that color transformation was due to L-asparaginase production. The bacterial isolates with highest L-asparaginase activity were identified as Erwinia carotovora, E. coli and Streptococcus pyogenes.
In this study the diameter zones of L-asparaginase enzyme activity were determined, in which the isolate Erwinia carotovora has the highest intense pink zone (14 mm), followed by E. coli (13 mm), then Streptococcus pyogenes (12 mm). This is also similar to the report of Devi and Ramanjaneyulu23, who reported that strain exhibiting diameter zone above 9 mm are referred as good L-asparaginase producers, those strains with diameter zone 6-9 mm and those having below 6 mm diameter zone may be referred to as moderate and poor L-asparaginase producers, respectively.
Erwinia carotovora was subjected to several physiological studies to improve its enzyme production. Effect of incubation temperature of L-asparaginase enzyme production was determine in shake flasks using basal medium. The maximum specific activity of L-asparaginase (2.7 U mg–1 ) was obtained at 37°C. This indicates that the enzyme production was reduced gradually with further increase in incubation temperature. This results were similar to the study of Abbas and Askar24 and Maladkar et al.25, who reported that optimum temperature is 37°C of L-asparaginase enzyme activity by Erwinia spp., from Staphylococcus spp. by Prakasham et al.26 and from Pseudomonas stutzeri by Manna et al.27.
Our results disagreed with Wakil and Adelegan3, who indicated that the optimum incubation temperature at which Bacillus firmus, Streptococcus spp. and Bacillus circulans showed highest asparaginase yield was 45°C. Borkotaky and Bezbaruah28 that reported optimum incubation temperature for L-asparaginase activity by Erwinia spp. was 35°C. Narayana et al.29, who indicated that the optimum incubation temperature for Streptomyces albidoflavus, Bacillus polymyxa and Paenibacillus validus had their highest production at incubation temperature of 35°C. Ghosh et al.30, who proved that the highest production of L-asparaginase from Serratia narcerens at 30°C.
Among the physical parameters, pH of the growth medium plays an important role by inducing morphological changes in the microorganisms and in enzyme secretion. The result of the effect of pH showed that maximum L-asparaginase yield by Erwinia cacrotovora was at pH 7.0. these results was agreed with Ghosh et al.30, who indicated that the maximum L-asparaginase production from Serratia narcerens was at pH 7.0. However Streptomyces albidoflavus was recorded most active at pH 7.5 by Narayana et al.29. This result disagreed with wakil et al.3, who proved that the highest yields of L-asparaginase enzyme from Streptococcus spp., Streptococcus spp. and Bacillus polymyxa was at pH 8.0. Dhevagi and Poorani31, who reported that the maximum L-asparaginase by Streptomyces spp. was at pH 8.0-8.5. However, Deshpande et al.32 and Akilandeswari et al.33 proved that Streptomyces ginsengisoli produced the highest yields of L-asparaginase enzyme at pH 8.0 and 9.0, respectively. Abdel-Fattah and Olama34 indicated that Streptomyces albidoflavus was recorded most active at pH 8.0.
The induction of Erwinia carotovora was studied using media with L-asparagine and other various carbon sources. The data obtained indicated that the maximum L-asparaginase activity (3.5 U mg–1) was recorded in presence of glucose as carbon source. These findings were comparable to that described by Prakasham et al.26 who reported that glucose was the best carbon source under aerobic conditions for the synthesis of L- asparaginase by Staphylococcus spp. Venil et al.35 has reported decrease in L-asparaginase production when supplemented with fructose. Prakasham et al.26 reported that Staphylococcus spp. which was recorded most active when supplemented with maltose and Akilandeswari et al.33 used proline as the best carbon source for L-asparaginase production. Wakil et al.3, who indicated that mannitol, sucrose and maltose were the best carbon sources for Bacillus firmus, Streptococcus spp. however, Kenari et al.36 reported that lactose was the best carbon source for L-asparaginase production. In contrast, Deokar et al.37 observed maximum L-asparaginase production by Erwinia carotovora at 1.13% maltose concentration.
Different nitrogen sources in culture media were investigated for enzyme production by Erwinia carotovora. L-asparagine was the most suitable nitrogen source for optimum L-asparaginase production by Erwinia carotovora with specific activity (4.7 U mg–1). this result agreed with Nawaz et al.38, who reported that L-asparagine was the best nitrogen source for L-asparaginase enzyme production. Wakil et al.3, Verma et al.39, Narayana et al.29 and Deokar et al.37 showed that yeast extract was optimum for L-asparaginase synthesis by Bacillus polymyxa and Erwinia carotovora. However, Deshpande et al.32 used peptone as the best nitrogen source.
By studying the effect of different concentration of the carbon source which represented in glucose, it was found that 10 gm L–1 (1%) was the best concentration and give the maximum L-asparaginase production. These findings were comparable to that described by Amena et al.19 and Narayana et al.29. However, Prakasham et al.26 and Ghosh et al.30 indicated that the best concentration of glucose for production high yields of L-asparaginase was 20 gm L–1.
By studying the effect of different concentration of the nitrogen source which represented in L-asparagine, it was found that 5 gm L–1 (0.5%) was the best concentration and give the maximum L-asparaginase production. These observation was equivalent to that had been found by Prakasham et al.26. But Amena et al.19 had found 2 gm L–1 (0.2%) was the best concentration.
L-asparaginase enzyme produced by Erwinia carotovora was purified using ammonium sulphate precipitation (70%)40. Followed by gel filtration (fractionation) using Sephadex G100 (Patro and Gupta18). In the present study some biochemical properties of L-asparaginase enzyme produced by Erwinia carotovora have been investigated. The optimum incubation period of high relative enzymes activities and the rate of substrate hydrolysis was investigated and found that L-asparaginase activity was obtained after incubation 10 min. These findings were comparable to that described by Moorthy et al.17. For temperature profile for L-asparaginase enzyme was found that this enzyme showed an optimum temperature at 37°C. These findings were comparable to that described by Abbas and Gupta24, Moorthy et al.17 and Patro and Gupta18. For pH profile using broad pH range from pH 5-9 using different concentrations of borate buffer. L-asparaginase enzyme produced by Erwinia carotovora exhibited an optimum pH 8.0 and this result was similar to that deduced by Dhevagi and Poorani 31 but Moorthy et al.17 had found pH 8 the best for L-asparaginase activity. The effect of activator MgCI2 on L-asparaginase enzyme was determined by adding the activator in the reaction mixture. The enzyme was activated by MgC12 and these by 1.0 mL of activator. These similar to that described before by Moorthy et al.17. Our study recommended that soil can be a good source of L-asparaginase enzyme produced bacteria and evaluate the different conditions that can produce a maximum amount of L- asparaginase enzyme.
CONCLUSION
This study clearly indicates that soil can provide a good source of L-asparaginase producing bacteria and Erwinia carotovora was the most active bacterial isolate for L-asparaginase production from the isolated bacterial species. Glucose was the best carbon source at conc. (10 gm L–1) and L-asparagine the best nitrogen source at conc. (5 gm L–1). A characterization of partially purified L-asparaginase from Erwinia carotovora has been done. It was investigated that this enzyme showed an optimum activity after 10 min at 37°C, pH 8 and highly affected by adding 1.0 mL MgC12 as activator.
SIGNIFICANCE STATEMENT
This study discovered the new methods for isolation and production of L-asparaginase enzyme that can be beneficial for L-asparaginase enzyme production.
REFERENCES
- Piatkowska-Jakubas, B., M. Krawczyk-Kulis, S. Giebel, M. Adamczyk-Cioch and A. Czyz et al., 2008. Use of L-asparaginase in acute lymphoblastic leukemia: Recommendations of the Polish Adult Leukemia Group. Pol. Arch. Med. Wewn, 118: 664-669.
CrossRefDirect Link - Muthusivaramapandian, M., S. Arrivukkarasan, R. Aravindan and T. Viruthagiri, 2008. Perspectives and applications of anticancer enzyme L-asparaginase. Asian J. Microbiol. Biotechnol. Environ. Sci., 10: 851-854.
Direct Link - Wakil, S.S.M. and A.A. Adelegan, 2015. Screening, production and optimization of L-asparaginase from soil bacteria isolated in Ibadan, South-Western Nigeria. J. Basic Applied Sci., 11: 39-51.
Direct Link - Appel, I.M., C. van Kessel-Bakvis, R. Stigter and R. Pieters, 2007. Influence of two different regimens of concomitant treatment with asparaginase and dexamethasone on hemostasis in childhood acute lymphoblastic leukemia. Leukemia, 21: 2377-2380.
CrossRefPubMedDirect Link - Stams, W.A., M.L. den Boer, H.B. Beverloo, J.P. Meijerink and R.L. Stigter et al., 2003. Sensitivity to L-asparaginase is not associated with expression levels of asparagine synthetase in t (12; 21)+ pediatric ALL. Blood, 101: 2743-2747.
Direct Link - Cachumba, J.J.M., F.A.F. Antunes, G.F.D. Peres, L.P. Brumano, J.C. dos Santos and S.S. da Silva, 2016. Current applications and different approaches for microbial L-asparaginase production. Braz. J. Microbiol., 47: 77-85.
CrossRefDirect Link - Arjun, J.K., B.P. Aneesh, T. Kavitha and K. Harikrishnan, 2018. Characterization of a novel asparaginase from soil metagenomic libraries generated from forest soil. Biotechnol. Lett., 40: 343-348.
CrossRefDirect Link - El-Naggar, N.E.A., S.F. Deraz, S.M. El-Ewasy and G.M. Suddek, 2018. Purification, characterization and immunogenicity assessment of glutaminase free L-asparaginase from Streptomyces brollosae NEAE-115. BMC Pharmacol. Toxicol., Vol. 19, No. 1.
CrossRefDirect Link - Ashok, A., K. Doriya, J.V. Rao, A. Qureshi, A.K. Tiwari and D.S. Kumar, 2019. Microbes producing L-Asparaginase free of glutaminase and urease isolated from extreme locations of antarctic soil and moss. Scient. Rep., Vol. 9, No. 1.
CrossRefDirect Link - Kumar, N.S.M., C.A. Shimray, D. Indrani and H.K. Manonmani, 2014. Reduction of acrylamide formation in sweet bread with L-asparaginase treatment. Food Bioprocess Technol., 7: 741-748.
CrossRefDirect Link - Tareke, E., P. Rydberg, P. Karlsson, S. Eriksson and M. Törnqvist, 2000. Acrylamide: A cooking carcinogen? Chem. Res. Toxicol., 13: 517-522.
CrossRefDirect Link - Zhang, S., Y. Xie, C. Zhang, X. Bie, H. Zhao, F. Lu and Z. Lu, 2015. Biochemical characterization of a novel l-asparaginase from Bacillus megaterium H-1 and its application in French fries. Food Res. Int., 77: 527-533.
CrossRefDirect Link - Vinci, R.M., F. Mestdagh and B. De Meulenaer, 2012. Acrylamide formation in fried potato products-present and future, a critical review on mitigation strategies. Food Chem., 133: 1138-1154.
CrossRefDirect Link - Avramis, V.I. and P.N. Tiwari, 2006. Asparaginase (native ASNase or pegylated ASNase) in the treatment of acute lymphoblastic leukemia. Int. J. Nanomed., 1: 241-254.
PubMedDirect Link - Kamble, K.D., P.R. Bidwe, V.Y. Muley, L.H. Kamble, D.G. Bhadange and M. Musaddiq, 2012. Characterization of L-asparaginase producing bacteria from water, farm and saline soil. Biosci. Discov., 3: 116-119.
Direct Link - Prakasham, R.S., M. Hymavathi, C.S. Rao, S.K. Arepalli and J.V. Rao et al., 2010. Evaluation of antineoplastic activity of extracellular asparaginase produced by isolated Bacillus circulans. Applied Biochem. Biotechnol., 160: 72-80.
CrossRefPubMedDirect Link - Moorthy, V., A. Ramalingam, A. Sumantha and R.T. Shankaranaya, 2010. Production, purification and characterisation of extracellular L-asparaginase from a soil isolate of Bacillus sp. Afr. J. Microbiol. Res., 4: 1862-1867.
Direct Link - Patro, K.R. and N. Gupta, 2012. Extraction, purification and characterization of L-asparaginase from Penicillium sp. by submerged fermentation. Int. J. Biotechnol. Mol. Biol. Res., 3: 30-34.
Direct Link - Amena, S., N. Vishalakshi, M. Prabhakar, A. Dayanand and K. Lingappa, 2010. Production, purification and characterization of L-asparaginase from Streptomyces gulbargensis. Braz. J. Microbiol., 41: 173-178.
CrossRefDirect Link - Kumar, D.S. and K. Sobha, 2012. L-Asparaginase from microbes: A comprehensive review. Adv. Biores., 3: 137-157.
Direct Link - Mohapatra, B.R., R.K. Sani and U.C. Banerjee, 1995. Characterization of L-asparaginase from Bacillus sp. isolated from an intertidal marine alga (Sargassum sp.). Lett. Applied Microbiol., 21: 380-383.
CrossRefDirect Link - Gulati, R., R.K. Saxena and R. Gupta, 1997. A rapid plate assay for screening L-asparaginase producing micro-organisms. Lett. Applied Microbiol., 24: 23-26.
CrossRefPubMedDirect Link - Devi, A.L. and R. Ramanjaneyulu, 2016. Isolation of L-asparaginase producing microbial strains from soil samples of Telangana and Andhra Pradesh states, India. Int. J. Curr. Microbiol. Applied Sci., 5: 1105-1113.
CrossRefDirect Link - Maladkar, N.K., V.K. Singh and S.R. Naik, 1993. Fermentative production and isolation of L-asparaginase from Erwinia carotovora, EC-113. Hindustan Antibiot. Bull., 35: 77-86.
PubMedDirect Link - Prakasham, R.S., C.S. Rao, R.S. Rao, G.S. Lakshmi and P.N. Sarma, 2007. L-asparaginase production by isolated Staphylococcus sp. -6A: Design of experiment considering interaction effect for process parameter optimization. J. Applied Microbiol., 102: 1382-1391.
CrossRefPubMedDirect Link - Manna, S., A. Sinha, R. Sadhukhan and S.L. Chakrabarty, 1995. Purification, characterization and antitumor activity of L-asparaginase isolated from Pseudomonas stutzeri MB-405. Curr. Microbiol., 30: 291-298.
CrossRefDirect Link - Borkotaky, B. and R.L. Bezbaruah, 2002. Production and properties of asparaginase from a new Erwinia sp. Folia Microbiol., 47: 473-476.
CrossRefPubMedDirect Link - Narayana, K.J.P., K.G. Kumar and M. Vijayalakshmi, 2008. L-asparaginase production by Streptomyces albidoflavus. Indian J. Microbiol., 48: 331-336.
CrossRefDirect Link - Ghosh, S., S. Murthy, S. Govindasamy and M. Chandrasekaran, 2013. Optimization of L-asparaginase production by Serratia marcescens (NCIM 2919) under solid state fermentation using coconut oil cake. Sustainable Chem. Processes, Vol. 1, No. 1.
CrossRefDirect Link - Dhevagi, P. and E. Poorani, 2006. Isolation and characterization of L-Asparaginase from marine actinomycetes. Ind. J. Biotechnol., 5: 514-520.
Direct Link - Deshpande, N., P. Choubey and M. Agashe, 2014. Studies on optimization of growth parameters for L-asparaginase production by Streptomyces ginsengisoli. Scient. World J., Vol. 2014.
CrossRefDirect Link - Akilandeswari, K., K. Kavitha and M. Vijayalakshmi, 2012. Production of bioactive enzyme L-asparaginase from fungal isolates of water sample through submerged fermentation. Int. J. Pharm. Pharm. Sci., 4: 363-366.
Direct Link - Abdel-Fattah, Y.R. and Z.A. Olama, 2002. L-asparaginase production by Pseudomonas aeruginosa in solid-state culture: Evaluation and optimization of culture conditions using factorial designs. Process Biochem., 38: 115-122.
CrossRefDirect Link - Venil, C.K., K. Nanthakumar, K. Karthikeyan and P. Lakshmanaperumalsamy, 2009. Production of L-asparaginase by Serratia marcescens SB08: Optimization by response surface methodology. Iran. J. Biotechnol., 7: 10-18.
Direct Link - Kenari, S.L.D., I. Alemzadeh and V. Maghsodi, 2011. Production of L-asparaginase from Escherichia coli ATCC 11303: Optimization by response surface methodology. Food Bioprod. Proces., 89: 315-321.
CrossRefDirect Link - Deokar, V.D., M.D. Vetal and L. Rodrigues, 2010. Production of intracellular L-asparaginase from Erwinia carotovora and its statistical optimization using Response Surface Methodology (RSM). Int. J. Chem. Sci. Applic., 1: 25-36.
Direct Link - Nawaz, M.S., D. Zhang, A.A. Khan and C.E. Cerniglia, 1998. Isolation and characterization of Enterobacter cloacae capable of metabolizing asparagine. Applied Microbiol. Biotechnol., 50: 568-572.
CrossRefDirect Link - Verma, N., K. Kumar, G. Kaur and S. Anand, 2007. E. coli K-12 asparaginase-based asparagine biosensor for leukemia. Artif. Cells Blood Substitutes Biotechnol., 35: 449-456.
CrossRefDirect Link - Warangkar, S.C. and C.N. Khobragade, 2010. Purification, characterization and effect of thiol compounds on activity of the Erwinia carotovora L-asparaginase. Enzyme Res., Vol. 2010.
CrossRefDirect Link