
Kasimu Ghandi Ibrahim
Department of Physiology, Faculty of Basic Medical Sciences, College of Health Sciences, Usmanu Danfodiyo University, P.M.B. 2254, Sokoto, Nigeria
LiveDNA: 234.16858
Eliton Chivandi
School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, Parktown 2193, Johannesburg, South Africa
Kennedy Honey Erlwanger
School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, Parktown 2193, Johannesburg, South Africa
Richard Leslie Brooksbank
Cardiovascular Pathophysiology and Genomics Research Unit, School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, Parktown 2193, Johannesburg, South Africa
Journal of Biological Sciences
Year: 2019 | Volume: 19 | Issue: 3 | Page No.: 224-230







ABSTRACT
Background and Objective: Telomere lengths are affected epigenetically during stages of developmental plasticity. Whether curcumin and/or fructose administered to pups affects their leucocyte telomere lengths in adolescence following post-weaning a high fructose diet was investigated. Materials and Methods: One hundred and twenty eight suckling pups were gavaged with either a 0.5% DMSO, 500 mg kg–1 curcumin, fructose (20%, w/v) or a combination of curcumin and fructose for 15 days. On post-natal day 21, each of the initial groups was split into two subgroups: One had plain drinking water while the other was provided fructose (20%, w/v) as their drinking fluid for 6 weeks. The body masses of the rats were measured before termination with sodium pentobarbitone (150 mg kg–1, i.p.). Genomic DNA was extracted from whole blood. Relative leucocyte telomere length was determined using real time polymerase chain reaction. Results: The administration of curcumin and or fructose during suckling in Sprague Dawley rat pups, did not have any significant effects (p>0.05, ANOVA) on the body masses and relative leucocyte telomere lengths in both male and female rats across the treatment groups. Conclusion: Curcumin and fructose, though previously shown to have epigenetic effects did not alter the relative leucocyte telomere lengths in the growing rats and may therefore not lead to increased telomere attrition.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Kasimu Ghandi Ibrahim, Eliton Chivandi, Kennedy Honey Erlwanger and Richard Leslie Brooksbank, 2019. Neonatal Oral Curcumin: Effect on Telomere Lengths of Adolescent Rats Fed a High Fructose Diet Post-weaning. Journal of Biological Sciences, 19: 224-230.
DOI: 10.3923/jbs.2019.224.230
URL: https://scialert.net/abstract/?doi=jbs.2019.224.230
DOI: 10.3923/jbs.2019.224.230
URL: https://scialert.net/abstract/?doi=jbs.2019.224.230
INTRODUCTION
Telomeres are specialized noncoding repetitive nucleotide sequences (TTAGGG) found at the end of eukaryotic deoxyribonucleic acid (DNA)1. Their main function is to ensure chromosome stability during mitosis2 and protect the DNA from damage by the cellular repair processes, fusion of adjacent genes and erosion of genetic material 1. Leucocyte telomere length indicates chronological ageing in cells3 and is negatively associated with chronic diseases including obesity4. In addition to genetics, telomere lengths are determined by environmental factors5 that generate oxidative stress and inflammation6. Telomere length has been shown to be susceptible to programmed development during the early ontogenic periods7. Although telomere lengths exhibit high heritability8, embryonic telomere lengths are subject to the influences of in utero and post-natal life conditions9. These conditions exert stable and long-term effects on the telomeres through epigenetic mechanisms such as inhibition of the deoxyribonucleic acid methyltransferase-1 (DNMT-1)10. For example, a high maternal folate concentration in early pregnancy was found to correlate with longer foetal telomere lengths compared to those with low folate concentration11. This finding indicates that foetal telomeres show a developmental plasticity that is affected by maternal nutrition and which may accelerate ageing12.
The period extending from gestation to the end of lactation is a window of developmental plasticity in rats when epigenetic changes can modify the expression or repression of genes13. Nutritional interventions in the neonatal period interfere with the establishment of the normal gut micro-biome14 which in addition to its essential role in nutrient absorption and energy homeostasis also serves as a source of short chain fatty acids (SCFA) like butyrate, that acts epigenetically as a histone deacetylase inhibitor14.
Fructose showed a hypomethylating effect in mitochondrial DNA of Sprague Dawley rats when administered as a 20% w/v solution for 14 weeks15. The DNA methylation is involved in myeloid cell differentiation from their stem cell progenitors16 and exhibits remarkable stability that allows it to be transmitted through subsequent cell divisions17. Thus, the effects of interventions administered during the periods of developmental plasticity can have a long-lasting impact on the subsequent generations of circulating blood cells.
Curcumin (diferuloylmethane) a polyphenol found in the dried roots of turmeric (Curcuma longa) has anti-oxidant, anti-inflammatory18, anti-obesity19 and anti-diabetic20 among other metabolic effects. These effects were shown in both in vitro and in vivo adult rodent models and human studies. Additionally, it has been shown that curcumin not only controls gene expression through DNA methylation, histone modification or production of non-coding micro RNAs21,22 but also inhibits telomerase22.
We thus investigated whether neonatal oral administration of curcumin and/or fructose would induce epigenetic changes that would affect the relative leucocyte telomere lengths of rats later in adolescence after being fed a high fructose diet.
MATERIALS AND METHODS
Study location and ethical approval: The study was conducted from May 2016 to April 2018 in the rodent section of the Central Animal Services (C.A.S.) unit of the University of the Witwatersrand, Johannesburg and Laboratories of the School of Physiology in the Faculty of Health Sciences building. All the protocols used in the study complied with international guidelines for the use of laboratory animals and were approved by the Animal Ethics Screening Committee with certificate number: 2016/04/18/B.
Chemicals and reagents: All the chemicals and reagents used in the study were of analytical grade. Curcumin and dimethyl sulfoxide (DMSO) were supplied by Sigma-Aldrich (St. Louis, Missouri, USA). Fructose was from Nature’s Choice (Randvaal, South Africa). The SYBR™ Green Master Mix (Applied Biosystems, Thermo Fisher Scientific, Austin, TX, USA) Forward and reverse primers (telomeres and pyruvate kinase) and nuclease-free water (Inqaba Biotech, Pretoria, South Africa) were also used in the study.
Study design: A total of 128, 6-days old suckling rat pups from 14 Sprague Dawley dams were used in the two-phase study. In phase one, the pups were randomly allocated to four groups in a split litter manner and received treatment as follows: 0.5% dimethyl sulfoxide (DMSO) (n = 32, males = 16, females = 16), fructose solution (20%, w/v) in 0.5% DMSO (n = 32, males = 16, females = 16), curcumin (500 mg kg–1) suspended in 0.5% DMSO (n = 33, males = 17, females = 16) and a combination of curcumin (500 mg kg–1) and fructose (20%, w/v) in 0.5% DMSO (n = 31, males = 16, females = 15). These initial treatments were administered from the post-natal day (PND) six up to weaning at PND 21 at a volume of 10 mL kg–1 body mass via oral gavage.
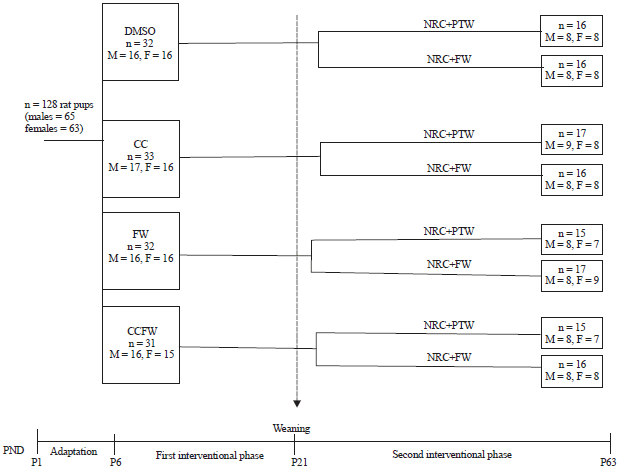 | |
| Fig. 1: | Schematic representation of the experimental design |
| DMSO: Dimethyl sulfoxide, CC: Curcumin 500 mg kg–1 body mass, FW: Fructose water (20%, w/v), NRC: Normal rat chow, PTW: Plain tap water, P: Postpartum day | |
The litters were housed in perspex cages with their respective dams and allowed to suckle freely from their dams for the duration of the first phase of the study. The dams were fed with normal rat chow (Epol®, Centurion, South Africa) and plain tap water ad libitum.
In phase two, the rats were weaned onto normal rat chow and then the initial groups were further split into two sub-groups each: One group had tap water only as their drinking fluid while the rats in the other group were fed a high fructose diet (20%, w/v) as their drinking fluid. These treatments were provided ad libitum for a period of 6 weeks.
In the first phase of the study, the pups were weighed daily to adjust drug concentration and ensure constant dosing, while in the second phase, the rats were weighed twice weekly. A schematic diagram summarizing the experimental design is shown in Fig. 1.
Terminal procedures: On the morning of termination, the terminal body masses of the rats were determined with an electronic weighing scale (Snowrex Electronic Scale, Clover Scales, Johannesburg). The rats were euthanised on PND 63 by anaesthetic overdose using intraperitoneal sodium pentobarbitone (150 mg kg–1). The thorax was opened to expose the heart from where blood was taken and transferred into heparinised blood collecting tubes (Becton Dickinson VACUTAINER Systems Europe, Meylan Cedex, France). Whole blood was used for deoxyribonucleic acid (DNA) extraction either immediately or when not possible the blood was stored in a fridge (DEFY, Durban, South Africa) at 4°C.
Deoxyribonucleic acid (DNA) extraction: Genomic DNA extraction from whole blood was done using the NucleoSpin Blood QuickPure kit (Macherey-Nagel, Düren, Germany), according to the manufacturer’s instructions.
Real-time quantitative PCR: Leucocyte telomere lengths were determined using the real-time quantitative polymerase chain reaction (RT-qPCR). The RT-qPCR was done with the StepOnePlus™ Real-Time PCR system (Applied Biosystems, Singapore). This method has been previously described23,24. In the present study, it determined the factor by which telomere repeat copy number (T) differed from the copy number of pyruvate kinase (S)24.
A master mix was prepared for each of the runs (telomeres and pyruvate kinase (PK)). The master mix consisted of Power Up™ SYBR™ Green Master Mix, Forward and Reverse Primers (telomeres and PK) and nuclease-free water. The PCR runs were conducted in two separate 96-well plates, one each for telomeres and PK. The genomic DNA samples, which were run in duplicates were matched in terms of position in the wells to give the lowest variability23. It pipetted 2 μL of sample and 18 μL of the master mix in each of the wells. A control DNA sample and a blank (nuclease-free water) were included in the plates and served as a reference sample and endogenous control, respectively. Each of the amplifications was then run in the RT-qPCR machine using appropriate calibrations. The relative telomere length was expressed as the ratio of the telomere repeat copy number and the single gene copy number (T/S).
Statistical analysis: Data were analyzed using the statistical software GraphPad Prism version 7.0 (GraphPad Software Inc., San Diego, CA) and expressed as means±standard error of mean (SEM). A one-way analysis of variance (ANOVA) was used to analyze the data followed by a Bonferroni post hoc test to compare the means. The level of significance was set at p<0.05.
RESULTS
Figure 2a shows the terminal body masses of adolescent male Sprague Dawley rats. There were no significant differences (p>0.05, ANOVA) in the terminal body masses across the treatment groups in the male rats. The terminal body masses of adolescent female Sprague Dawley rats are presented in Fig. 2b. The terminal body masses of the adolescent female rats were similar (p>0.05, ANOVA) across the treatment groups.
Figure 3a shows the relative leucocyte telomere lengths of the male adolescent experimental rats that were administered with oral curcumin during suckling and a high fructose diet post-weaning. The relative telomere lengths were similar (p>0.05, ANOVA) across the treatment groups in the rats. The CC+FW group in the male rats had relatively shorter telomere lengths than the FW+FW group, though not statistically significant (p = 0.0517, ANOVA). The relative leucocyte telomere lengths of adolescent female Sprague Dawley rats that were administered with oral curcumin during suckling and a high fructose diet postweaning are shown in Fig. 3b. The relative leucocyte telomere lengths of the female rats were not statistically different (p>0.05, ANOVA) across the treatment groups.
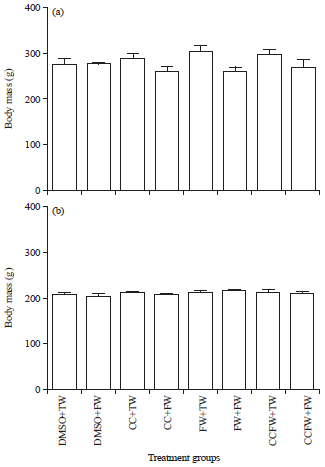 | |
| Fig. 2(a-b): | Terminal body masses of growing (a) Male and (b) Female sprague Dawley rats |
DMSO+TW: 10 mL kg–1 of a 0.5% dimethyl sulfoxide solution as neonates and plain tap water post-weaning, DMSO+FW: 10 mL kg–1 of a 0.5% dimethyl sulfoxide solution as neonates and fructose (20%, w/v) as drinking fluid in the growing period, CC+TW: Curcumin (500 mg kg–1 in 0.5% DMSO) as neonates and plain tap water in the post-weaning period, CC+FW: Curcumin (500 mg kg–1 in 0.5% DMSO) as neonates and fructose (20%, w/v) as drinking fluid in the post-weaning period, FW+TW: Fructose (20%, w/v) as neonates and plain tap water in the post-weaning period, FW+FW: Fructose (20%, w/v) as neonates and fructose (20%, w/v) as drinking fluid in the post-weaning period, CCFW+TW: Curcumin (500 mg kg–1) and fructose (20%, w/v) in 0.5% DMSO as neonates and plain tap water in the post-weaning period, CCFW+FW: Curcumin (500 mg kg–1) and fructose (20%, w/v) in 0.5% DMSO as neonates and fructose (20%, w/v) in the post-weaning period. Data expressed as mean±SEM, n = 7-9 per group | |
DISCUSSION
In this study, there were no observed differences in the relative leucocyte telomere lengths of growing rats of both sexes across the different groups. This means that neither the neonatal oral administration of curcumin and/or fructose nor the post-weaning 6-weeks consumption of a 20% fructose solution (w/v) had any impact on the leucocyte telomere lengths of the rats.
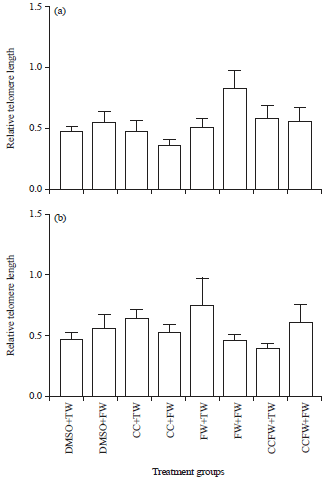 | |
| Fig. 3(a-b): | Effect of an oral administration of curcumin and/fructose during suckling on the relative leucocyte telomere lengths of growing (a) Male and (b) Female sprague Dawley rats that were fed a high fructose diet post-weaning |
DMSO+TW: 10 mL kg–1 o f a 0.5% dimethyl sulfoxide solution as neonates and plain tap water post-weaning, DMSO+FW: 10 mL kg–1 of a 0.5% dimethyl sulfoxide solution as neonates and fructose (20%, w/v) as drinking fluid post-weaning, CC+TW: Curcumin (500 mg kg–1 in 0.5% DMSO) as neonates and plain tap water in the post-weaning, CC+FW: Curcumin (500 mg kg–1 in 0.5% DMSO) as neonates and fructose (20%, w/v) as drinking fluid post-weaning, FW+TW: Fructose (20%, w/v) as neonates and plain tap water post-weaning, FW+FW: Fructose (20%, w/v) as neonates and fructose (20%, w/v) as drinking fluid post-weaning, CCFW+TW: Curcumin (500 mg kg–1) and fructose (20%, w/v) in 0.5% DMSO as neonates and plain tap water post-weaning, CCFW+FW: Curcumin (500 mg kg–1) and fructose (20%, w/v) in 0.5% DMSO as neonates and fructose (20%, w/v) as drinking fluid post-weaning. Data expressed as Mean±SEM, n = 7-9 per group | |
Previous studies as would be presented, however, have shown the telomere system to be affected by epigenetic modifications while curcumin and fructose are known to induce epigenetic changes.
The human telomerase reverse transcriptase (hTERT) gene promoter controls the appropriate expression of the hTERT gene25. When DNA methylation or histone modification of this promoter occurs, it represses the activity of telomerase, thus contributing to telomere attrition25. In addition to telomeres being prone to histone modifications26, the sub-telomeric DNA is also susceptible to methylation27. Several studies have pointed to a functional connection between the epigenetic mark and telomere lengths26,28 which indicates a key relationship between epigenetic changes and the regulation of telomere lengths29.
Curcumin has been shown to be a potential epigenetic modulator of telomere lengths in vitro through its ability to inhibit deoxyribonucleic acid methyltransferase-1 (DNMT-1) and histone deacetylase (HDAC)21,30 and to regulate micro-RNAs and their multiple target genes31. However, despite these observed epigenetic activities, its (curcumin) administration to suckling pups in our study did not have any effect on the telomere lengths of the rats later in adolescence.
Additionally, curcumin has direct actions on telomere lengths as reported in several studies. Through its inhibitory action on telomerase, curcumin was found to promote telomere attrition in an in vitro study using brain tumour cells22 and MCF-7 breast cancer cells32. Telomerase is a reverse transcriptase enzyme that maintains telomere lengths33. It is highly expressed in tumour cells but is almost undetectable in somatic cells, thus making it a potential tumour marker22. However, in a mouse model of Alzheimer’s disease, diets enriched with curcumin were found to enhance DNA stability and favour a trend for longer telomere lengths34. In the present study, curcumin had no effect on the telomere lengths of the rats across the treatment groups suggesting that it did not have an impact on the activity of telomerase.
A National Health and Nutrition Examination Survey revealed a strong association between the consumption of sugar-sweetened sodas and shorter telomere lengths35. Perhaps another link between fructose consumption and telomere lengths might be through the formation of advanced glycation end products which randomly bind to proteins or lipids and impair their functioning36. Glycation is usually associated with increased oxidative damage which has been linked to shorter telomere lengths36.
As earlier mentioned in the introduction, fructose, demonstrated a hypomethylating effect in mitochondrial DNA of Sprague Dawley rats when administered as a 20% w/v solution for 14 weeks14, thus it is postulated that the impact of a 20% fructose solution during the period of developmental plasticity and rapid growth might have a programming effect on telomere lengths. However, it has been demonstrated that the administration of a high fructose diet during suckling and extending through to adolescence did not affect the leucocyte telomere length of the rats.
The interventions did not have any adverse effects on the growth of the rats as shown by the similarity in the body masses of the rats taken at termination. Previous studies have shown that with obesogenic dietary interventions from the neonatal stage, significant differences in body mass were only noted after 100 days37. The rats were 63 days old at the termination of the study which was before the onset of the adolescent surge in sex hormones which also have a sex-specific effect on body mass gain.
CONCLUSION
Despite the previously established epigenetic modifying effects of curcumin and fructose and the susceptibility of the telomere system to epigenetic changes. We did not find any difference in the telomere lengths of the rats in this study. This suggests that the telomere attrition associated with curcumin and fructose intake may not manifest in adolescence but later in life during adulthood.
SIGNIFICANCE STATEMENT
The consequences of an early administration of curcumin and fructose in neonates on cellular ageing in adolescence need to be further evaluated. This is more so because both curcumin and fructose are known epigenetic modulators that have previously been shown to accelerate ageing in adult populations. The knowledge of their effects during periods of developmental plasticity may provide new frontiers for the prevention and treatment of ageing related diseases.
ACKNOWLEDGMENT
The assistance of the entire staff of the Central Animal Service Unit, University of the Witwatersrand, Johannesburg in the general care and welfare of the animals is acknowledged. This study was supported by grants from the Faculty of Health Sciences Research Committee of the University of the Witwatersrand, Johannesburg (KGI, Grant number: 001.251.8521101.5121105.000000. 0000000000.4550) and the National Research Foundation of South Africa (KHE, Grant number: IRF 2010041900009).
REFERENCES
- Zierer, J., G. Kastenmuller, K. Suhre, C. Gieger and V. Codd et al., 2016. Metabolomics profiling reveals novel markers for leukocyte telomere length. Aging, 8: 77-86.
CrossRefPubMedDirect Link - Lee, M., H. Martin, M.A. Firpo and E.W. Demerath, 2011. Inverse association between adiposity and telomere length: The fels longitudinal study. Am. J. Hum. Biol., 23: 100-106.
CrossRefDirect Link - Demissie, S., D. Levy, E.J. Benjamin, L.A. Cupples and J.P. Gardner et al., 2006. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the Framingham heart study. Aging Cell, 5: 325-330.
CrossRefDirect Link - Iwama, H., K. Ohyashiki, J.H. Ohyashiki, S. Hayashi and N. Yahata et al., 1998. Telomeric length and telomerase activity vary with age in peripheral blood cells obtained from normal individuals. Hum. Genet., 102: 397-402.
CrossRefDirect Link - Muezzinler, A., A.K. Zaineddin and H. Brenner, 2014. Body mass index and leukocyte telomere length in adults: A systematic review and meta-analysis. Obesity Rev., 15: 192-201.
CrossRefDirect Link - Von Zglinicki, T., 2000. Role of oxidative stress in telomere length regulation and replicative senescence. Ann. N. Y. Acad. Sci., 908: 99-110.
CrossRefDirect Link - Tellechea, M., T.F. Gianotti, J. Alvarinas, C.D. Gonzalez, S. Sookoian and C.J. Pirola, 2015. Telomere length in the two extremes of abnormal fetal growth and the programming effect of maternal arterial hypertension. Scient. Rep., Vol. 5.
CrossRefDirect Link - Mangino, M., S.J. Hwang, T.D. Spector, S.C. Hunt and M. Kimura et al., 2012. Genome-wide meta-analysis points to CTC1 and ZNF676 as genes regulating telomere homeostasis in humans. Hum. Mol. Genet., 21: 5385-5394.
CrossRefDirect Link - Entringer, S., C. Buss and P.D. Wadhwa, 2012. Prenatal stress, telomere biology and fetal programming of health and disease risk. Sci. Signal., Vol. 5, No. 248.
CrossRefDirect Link - Shalev, I., S. Entringer, P.D. Wadhwa, O.M. Wolkowitz, E. Puterman, J. Lin and E.S. Epel, 2013. Stress and telomere biology: A lifespan perspective. Psychoneuroendocrinology, 38: 1835-1842.
CrossRefDirect Link - Entringer, S., E.S. Epel, J. Lin, E.H. Blackburn and C. Buss et al., 2015. Maternal folate concentration in early pregnancy and newborn telomere length. Ann. Nutr. Metab., 66: 202-208.
CrossRefDirect Link - Qasem, R.J., E. Yablonski, J. Li, H.M. Tang, L. Pontiggia and A.P. D'Mello, 2012. Elucidation of thrifty features in adult rats exposed to protein restriction during gestation and lactation. Physiol. Behav., 105: 1182-1193.
CrossRefDirect Link - Canani, R.B., M. Di Costanzo, L. Leone, G. Bedogni and P. Brambilla et al., 2011. Epigenetic mechanisms elicited by nutrition in early life. Nutr. Res. Rev., 24: 198-205.
CrossRefDirect Link - Yamazaki, M., E. Munetsuna, H. Yamada, Y. Ando and G. Mizuno et al., 2016. Fructose consumption induces hypomethylation of hepatic mitochondrial DNA in rats. Life Sci., 149: 146-152.
CrossRefDirect Link - Alvarez-Errico, D., R. Vento-Tormo, M. Sieweke and E. Ballestar, 2015. Epigenetic control of myeloid cell differentiation, identity and function. Nat. Rev. Immunol., 15: 7-17.
CrossRefDirect Link - Kim, M. and J. Costello, 2017. DNA methylation: An epigenetic mark of cellular memory. Exp. Mol. Med., Vol. 49, No. 4.
CrossRefDirect Link - Aggarwal, B.B., 2010. Targeting inflammation-induced obesity and metabolic diseases by curcumin and other nutraceuticals. Annu. Rev. Nutr., 30: 173-199.
CrossRefDirect Link - Weisberg, S.P., R. Leibel and D.V. Tortoriello, 2008. Dietary curcumin significantly improves obesity-associated inflammation and diabetes in mouse models of diabesity. Endocrinology, 149: 3549-3558.
CrossRefPubMedDirect Link - Kalani, A., P.K. Kamat, S.C. Tyagi and N. Tyagi, 2013. Synergy of homocysteine, MicroRNA, and epigenetics: A novel therapeutic approach for stroke. Mol. Neurobiol., 48: 157-168.
CrossRefPubMedDirect Link - Qureshi, I.A. and M.F. Mehler, 2010. Emerging themes in epigenetics: Implications for understanding and treating nervous system disorders. Epigenomics, 2: 345-349.
CrossRefDirect Link - Khaw, A.K., M.P. Hande, G. Kalthur and M.P. Hande, 2013. Curcumin inhibits telomerase and induces telomere shortening and apoptosis in brain tumour cells. J. Cell. Biochem., 114: 1257-1270.
CrossRefDirect Link - Cawthon, R.M., 2002. Telomere measurement by quantitative PCR. Nucleic Acids Res., Vol. 30, No. 10.
CrossRefDirect Link - Raymond, A.R., G.R. Norton, P. Sareli, A.J. Woodiwiss and R.L. Brooksbank, 2013. Relationship between average leucocyte telomere length and the presence or severity of idiopathic dilated cardiomyopathy in black Africans. Eur. J. Heart Failure, 15: 54-60.
CrossRefDirect Link - Zinn, R.L., K. Pruitt, S. Eguchi, S.B. Baylin and J.G. Herman, 2007. hTERT is expressed in cancer cell lines despite promoter DNA methylation by preservation of unmethylated DNA and active chromatin around the transcription start site. Cancer Res., 67: 194-201.
CrossRefDirect Link - Gonzalo, S., M. Garcia-Cao, M.F. Fraga, G. Schotta and A.H. Peters et al., 2005. Role of the RB1 family in stabilizing histone methylation at constitutive heterochromatin. Nat. Cell Biol., 7: 420-428.
CrossRefDirect Link - Bellon, M., H.H. Baydoun, Y. Yao and C. Nicot, 2010. HTLV-I Tax-dependent and -independent events associated with immortalization of human primary T lymphocytes. Blood, 115: 2441-2448.
CrossRefPubMedDirect Link - Gonzalo, S., I. Jaco, M.F. Fraga, T. Chen, E. Li, M. Esteller and M.A. Blasco, 2006. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol., 8: 416-424.
CrossRefDirect Link - Blasco, M.A., 2007. The epigenetic regulation of mammalian telomeres. Nat. Rev. Genet., 8: 299-309.
CrossRefDirect Link - Liu, Z., Z. Xie, W. Jones, R.E. Pavlovicz and S. Liu et al., 2009. Curcumin is a potent DNA hypomethylation agent. Bioorg. Med. Chem. Lett., 19: 706-709.
CrossRefPubMedDirect Link - Teiten, M.H., M. Dicato and M. Diederich, 2013. Curcumin as a regulator of epigenetic events. Mol. Nutr. Food Res., 57: 1619-1629.
CrossRefDirect Link - Pirmoradi, S., E. Fathi, R. Farahzadi, Y. Pilehvar-Soltanahmadi and N. Zarghami, 2018. Curcumin affects adipose tissue-derived mesenchymal stem cell aging through TERT gene expression. Drug Res., 68: 213-221.
CrossRefDirect Link - Criscuolo, F., S. Smith, S. Zahn, B.J. Heidinger and M.F. Haussmann, 2018. Experimental manipulation of telomere length: Does it reveal a corner-stone role for telomerase in the natural variability of individual fitness? Philos. Trans. R. Soc. B: Biol. Sci., Vol. 373, No. 1741.
CrossRefDirect Link - Thomas, P., Y.J. Wang, J.H. Zhong, S. Kosaraju, N.J. O'Callaghan, X.F. Zhou and M. Fenech, 2009. Grape seed polyphenols and curcumin reduce genomic instability events in a transgenic mouse model for Alzheimer's disease. Mutat. Res./Fundam. Mol. Mech. Mutagen., 661: 25-34.
CrossRefPubMedDirect Link - Leung, C.W., B.A. Laraia, B.L. Needham, D.H. Rehkopf and N.E. Adler et al., 2014. Soda and cell aging: Associations between sugar-sweetened beverage consumption and leukocyte telomere length in healthy adults from the national health and nutrition examination surveys. Am. J. Public Health, 104: 2425-2431.
CrossRefPubMedDirect Link - Nedic, O., S.I.S. Rattan, T. Grune and I.P. Trougakos, 2013. Molecular effects of advanced glycation end products on cell signalling pathways, ageing and pathophysiology. Free Radic. Res., 47: 28-38.
CrossRefDirect Link - Patel, M.S. and M. Srinivasan, 2010. Metabolic programming due to alterations in nutrition in the immediate postnatal period. J. Nutr., 140: 658-661.
CrossRefDirect Link