
Mahdhi Mosbah
Centre for Environmental Research and Studies, Jazan University, 82817 Jazan, Saudi Arabia
LiveDNA: 216.13300
Tounekti Taieb
Centre for Environmental Research and Studies, Jazan University, 82817 Jazan, Saudi Arabia
LiveDNA: 216.15670
Khemira Habib
Centre for Environmental Research and Studies, Jazan University, 82817 Jazan, Saudi Arabia
Journal of Biological Sciences
Year: 2018 | Volume: 18 | Issue: 3 | Page No.: 115-123







ABSTRACT
Background and Objective: The impact of plant invasions on ecosystems, habitats and native species is severe and often irreversible. The present study attempted to observe the effect of mesquite (Prosopis juliflora) on seed germination and plant growth of two native Acacia species (Acacia ehrenbergiana and Acacia tortilis). The effect of mesquite on mutualistic interactions and soil microbial biomass was also investigated. Materials and Methods: The effect of Prosopis juliflora (P. juliflora) extract on seed germination and plant growth of two native legumes was studied in petri dish and pot experiments, respectively. Root nodule bacteria, mycorrhizal fungi and soil microbial biomass were compared between the invasive legume and two native species. Results: The results showed that different aqueous extracts of Prosopis juliflora significantly affected the germination of native Acacia species. Plant growth of these species was also affected by the litter of the invasive plant. A relationship between mutualisms and the invasion process by Prosopis juliflora has been recorded. The MPN analysis showed that the density of rhizobia organisms which were able to nodulate Prosopis juliflora was greater than for Acacia species. Arbuscular mycorrhizae fungi colonization in the roots of native species was affected by Prosopis juliflora. Conclusion: Results suggested a significant effect of Prosopis juliflora on soil microbial biomass. This is the first study reporting the relationship between invasion process of Prosopis juliflora and soil microbial community composition in Saudi Arabia.
PDF Abstract XML References Citation
Received: October 26, 2017;
Accepted: February 07, 2018;
Published: March 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mahdhi Mosbah, Tounekti Taieb and Khemira Habib, 2018. Invasive Character of Prosopis juliflora Facilitated by its Allelopathy and a Wide Mutualistic Interaction with Soil Microorganisms. Journal of Biological Sciences, 18: 115-123.
DOI: 10.3923/jbs.2018.115.123
URL: https://scialert.net/abstract/?doi=jbs.2018.115.123
DOI: 10.3923/jbs.2018.115.123
URL: https://scialert.net/abstract/?doi=jbs.2018.115.123
INTRODUCTION
Invasive plant species are now considered one of the greatest threats to plant biodiversity. In their new habitat, invasive plant species become predators, competitors, parasites, breeders and disease transmitters for native plants1. Invasive plants can destabilize the environment and promote the establishment of other foreign plants2,3. Allelopathy is a biological phenomenon by which a plant produces one or more biochemical substance to influence growth, survival and reproduction of other plants. These allelochemicals can positively influence (positive allelopathy) or negative (negative allelopathy) on neighboring plants. Successful establishment of many plants in new environments can be explained by their capacity to produce allelochemical compounds which are toxic to native species4. Such compounds are particularly disruptive when they cannot be detoxified by soil microflora. Furthermore, it has also been shown that invasive plants affect soil rhizosphere microflora and alter plants-microbes mutualism5,6. Given the apparent role of mutualistic soil microbes in maintaining/determining plant community diversity and structure7, it is surprising that few studies have explored changes in their interactions with plants within communities in response to invasion8. Plant invasions can greatly impact soil microbial community composition and function and consequently, the above-ground structure and composition of native plant communities9. Furthermore, plant-microbe mutualistic interactions often facilitate invasions. The symbiotic association with both rhizobia and arbuscular mycorrhizae (AM) can be a major advantage for invading legumes.
Symbiotic relationships between legumes and rhizobia represent an ideal system to study the effects of invasions on below-ground plant-mutualism interaction networks. Rhizobia are capable of establishing symbiosis with legumes by entering their roots and inducing the formation of nitrogen fixing root nodules10. Native plants can be the source of symbiotic rhizobia for the invasive species11. The capacity of many introduced legumes to nodulate abundantly in new environments could be explained by the presence of a cosmopolitan rhizobial symbionts with a vast host range12. An increasing number of studies indicate that nodulation plays an important role in the invasion process13,14. Invasive plants also form symbiotic relationships with AM fungi (AMF) which facilitate phosphorus (P) uptake and plant growth. Although AMF can associate with a wide variety of plants from various geographical regions, the success of invasive legumes may depend on their ability to form effective symbiotic relationships with resident AMF populations of native ecosystems. Mycorrhizal colonization improves growth and nutrition of an invasive legume15. Actually the distinct advantage of invasive legumes may be attributed to tripartite rhizobia-AMF-plant symbiosis.
Prosopis juliflora is a vigorous evergreen tree belonging to the Leguminosae family which was introduced to Saudi Arabia from the Americas. Since it became invasive taking over large swaths of the wooded areas in Tihama plains and elsewhere at the expense of endogenous, well-adapted Acacia species16. Prosopis juliflora is a significant invasive plant in Asia, Australia and also Eastern and Southern Africa, where it impacted plant communities, soil microflora and ecosystem function17,18. Several countries attempted to check through the spread of this species eradication19. In Ethiopia and Sudan, mechanical methods were used, the trees were uprooted mechanically with machines or by hand. In other countries such as Yemen and Australia, chemical methods (kerosene or diesel oil application followed by burning) were used to kill the plants. In Australia, insects have been used as biological control agents against P. juliflora. Unfortunately, these methods don’t give the expected result, expensive and mostly ineffective20. In order to explain why P. juliflora is a better competitor than native tree species, the present study aimed to (i) Evaluate the effect of P. juliflora on seed germination and plant growth of two native woody legumes Acacia tortilis (A. tortilis), Acacia ehrenbergiana (A. ehrenbergiana) and (ii) Study the effect of P. juliflora on mutualistic interactions and soil microbial biomass.
MATERIALS AND METHODS
Plant materials and preparation of aqueous extracts: Litter, leaves and roots of P. juliflora were collected from different regions of Jazan region of Saudi Arabia (16.8894°N, 42.5706°E) in January, 2017. Approximately 500 g of each plant material was collected by randomly taking fragments from eight adult P. juliflora trees. A subsample of 10 g of plant materiel was soaked in 100 mL of distilled water for 24 h, homogenized and filtered with filter paper (Whatman No. 1). The extract was diluted with distilled water to 1:2 (50%) and 1:4 (25%) in order to obtain 3 applied concentrations: 100, 50 and 25%. These aqueous extracts were stored at -10°C. Part of the leaf litter sample was stored until used in pot experiments.
Germination bioassay: The goal of this experiment was to examine the effect of aqueous extract of P. juliflora on percent germination and germination speed of two native legumes (A. tortilis and A. ehrenbergiana). Germination trials were carried out in sterile petri dishes using 1% agar. Four milliliter of each concentration of aqueous extracts were added to each petri dish. The control was treated with 4 mL of sterile distilled water. Three replicates were used for each of the four treatments. Following seed sowing, germinated seeds were enumerated. During 5 days, germinated seeds were daily counted in order to get two variables: Percent germination (final number of germinated seeds×100/total number of seeds) and germination speed (S) calculated using the index provided by Einhelling et al.21:
where, N1, N2, N3, Nn represent the proportion of germinated seeds in day 1,2,3…n after the start of the experiment.
Pot experiments: Seedling were grown in pots to assess the effects of P. juliflora litter on plant growth of two target legumes. Seeds of each target legume were aseptically transferred into pots filled with autoclaved vermiculite. In each pot, 200 g of freeze-dried litter were crushed and mixed with autoclaved vermiculite. Eight replicates were considered. Control pots (without litter) were included. The pots were placed in a growth chamber at 25°C. Plants were watered once a week with a sterilized nutrient solution. Two months latter the plants were harvested, their shoots and roots were separated, rinsed free of vermiculite, dried at 70°C then weighed.
Estimation of legume-nodulating rhizobia populations: The enumeration of legume nodulating rhizobia associated with P. juliflora and the target legumes was carried out using the most probable number (MPN) method according to Vincent22. Soil was collected from three locations in Jazan (Sabia: 17.20°N, 42.62°E, Wadi Jazan: 16.98°N, 42.63°E and Abu Areesh: 17.02°N, 42.92°E). Soil was passed through a 2 mm sieve and stored at room temperature prior to the experiment. Seeds of the P. juliflora and the two target legumes were surface-sterilized and germinated in petri dishes. One seedling was aseptically transplanted into plastic pot filled with autoclaved vermiculite. Inoculation was performed 48 h after transfer with 2 mL of diluted soil suspensions. Four replicates were considered for each tree species. The pots were placed in a growth chamber at 23°C with a 14 h photoperiod and watered daily with sterilized distilled water. Thirty days latter, the presence or absence of nodules was recorded and the MPN was calculated according to Bennett et al.23.
AMF spore isolation: Soil samples were collected from three sites (described above). For each site, bulk soil and soil from understory of P. juliflora was collected. AMF spores were extracted from 100 g soil. AM spores were isolated by wet-sieving and sucrose centrifugation24. Quantification was carried out in petri dishes under a stereoscopic microscope. The spore density was expressed as the total number of spores per 100 g of soil25.
Assessment of root colonization by AM fungi: The symbiosis of AMF of two Acacia species (A. tortilis and A. ehrenbergiana) were investigated after seedlings were raised on soils collected under P. juliflora trees. Bulk soil was used as a control. Three sites (described above) were considered. The soil was passed through a 2 mm sieve and a mixture of soils from the three sites was used in this experiment. Three repetitions were considered for each soil type (i.e., bulk soil and soil collected from under P. juliflora) and for each tree species. Two months later, plant roots were collected, washed with sterile water, cleared by heating in 10% KOH at 90°C for 1 h, bleached by immersion in 10% H2O2 for 5 min, acidified in dilute HCl and stained with 0.05% trypan blue in lactophenol26. Stained roots were checked for AMF infection by examination under a compound microscope27. A minimum of 90 root segments per plant were counted. The intensity of mycorrhization (M) was assessed following the method of Trouvelot et al.28.
Soil microbial biomass: Soil microbial biomass carbon (Cmic) of soil sub-samples collected from under P. juliflora, A. tortilis and A. ehrenbergiana trees growing in three sites (described above) was determined by the fumigation extraction method29 using ninhydrin-N reactive compounds extracted from the soils with KCl after a 10 days fumigation period.
Statistical analysis: Statistical analyses were performed with a SAS statistical package. The data were subjected to ANOVA test (t-test ANOVA and one-way ANOVA). Comparisons among means were made using the least significant difference at the 5% level of significance (p<0.05).
RESULTS
Effect of P. juliflora aqueous extracts on germination and plant growth of A. tortilis and A. ehrenbergiana: Prosopis juliflora shoots aqueous extracts affected significantly the germination of the two Acacia species (Table 1). For the control, germination was 69 and 74% for A. ehrenbergiana,
| Table 1: | Effect of different concentrations of aqueous extracts of Prosopis juliflora on germination of Acacia ehrenbergiana (A. eh) and Acacia tortilis (A. to) seeds |
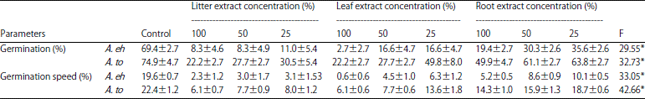 | |
Data are means±standard error of three replicates, *F: Statistic significant at p<0.05 | |
| Table 2: | Effect of litter of Prosopis juliflora on seedling growth of Acacia ehrenbergiana and Acacia tortilis |
 | |
*Significant difference between mean of growth parameter of treated and control seedling by t-test within species p<0.05 | |
A. tortilis, respectively. For extract-treated seeds, germination was reduced with increasing concentrations of the aqueous extracts. Litter and leaves seem to have greater inhibitory effects than roots. In comparison to control, germination of A. ehrenbergiana decreased about 61% when treated with 50% litter extract and about 53% when treated with 50% leaf extract. The 50% root extraction reduced the germination of A. ehrenbergiana and A. tortilis by about 39 and 13%, respectively. Germination speed of Acacia seeds was also significantly affected by P. juliflora aqueous extracts (Table 1). Germination speed of A. ehrenbergiana and A. tortilis decreased from 19 and 22% (For the control) to 4.56 and 7.73% (when treated with 50% leaf extract concentration), respectively.
Analysis of plant growth show that growth of A. ehrenbergiana and A. tortilis seedlings was not significantly affected by the litter of P. juliflora (Table 2). The only significant effect of litter of P. juliflora was a reduction of shoot length of A. tortilis and shoot fresh weight of A. ehrenbergiana. After 60 days, shoot length of A. tortilis and the shoot fresh weight of A. ehrenbergiana were 21 and 33% less than control seedlings, respectively.
Estimation of legume-nodulating rhizobia populations: The most probable number (MPN) of rhizobia able to nodulate the roots of P. juliflora, A. ehrenbergiana and A. tortilis seedlings growing in three soils prospected in this study presented in Fig. 1. The populations of indigenous legume nodulating rhizobia varied between sites.
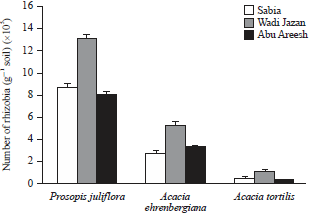 | |
| Fig. 1: | Most probable number (MPN) of rhizobia able to nodulate P. juliflora, A. ehrenbergiana and A. tortilis in soils from three Jazan sites (Sabia, Wadi Jazan and Abu Areesh) |
Error lines correspond to standard deviation (n = 3) | |
For all legumes, MPN was significantly greater in soil 2 (Wadi Jazan) than in soil 1 and soil 3. For all sites, MPN of rhizobia able to nodulate P. juliflora was significantly higher than A. ehrenbergiana and A. tortilis.
Assessment of root colonization by AM fungi and the number of spores: The microscopic observations have shown that the roots of all legumes considered in this study were colonized by endomycorrhizal fungi typical hyphae, arbuscules and vesicules were observed in the cortex of the roots of the three legumes, although not necessarily in the same root segment.
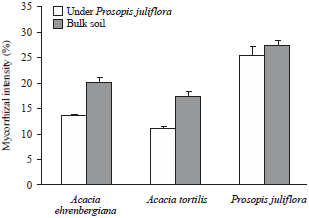 | |
| Fig. 2: | Mycorrhizal intensity in P. juliflora, A. ehrenbergiana and A. tortilis roots in bulk soil and soil collected from under P. juliflora) |
Error lines correspond to standard deviation of the means (n=3) | |
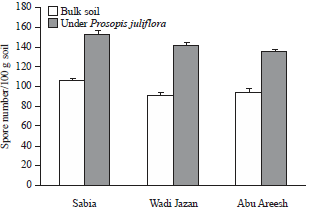 | |
| Fig. 3: | Spores number of the AM fungi in Jazan soils |
Error lines correspond to standard deviation of the means (n=3) | |
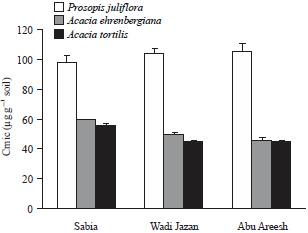 | |
| Fig. 4: | Rhizosphere soil microbial biomass carbon (Cmic) of P. juliflora, A. ehrenbergiana and A. tortilis collected from three sites in Jazan regions. (Sabia, Wadi Jazan and Abu Areesh) |
Error lines correspond to standard deviation (n = 3) | |
The intensity of mycorrhization of P. juliflora was significantly higher than that of native Acacia species (Fig. 2). The intensity of mycorrhization of A. ehrenbergiana and A. tortilis decrease significantly from 20 and 17% (for plant growing on bulk soil) to 13.7 and 11.1%, respectively, for plant growing on soil collected from under P. juliflora tree canopies.
Arbuscular mycorrhizal fungal spores were present in all soil samples and the density of spores did not differ among sites (Fig. 3). The majority of spores we collected were small with diameter less than 70 μm, comparable in shape and size to those of Glomus. The density of AMF spores varied highly among the treatments from 91 spores per 100 g of soil (bulk soil, Wadi Jazan) to 153 spores per 100 g of soil (in soil collected under P. juliflora, Sabia).
Soil microbiological biomass: Microbial biomass differed significantly between soils from under P. juliflora and native Acacia species (Fig. 4). In all sites, Cmic was greater in the rhizosphere of P. juliflora than in the rhizosphere of A. ehrenbergiana and A. tortilis. The highest value of Cmic was recorded in the rhizosphere of P. juliflora from Abu Areesh (105.3±5.7). The lowest Cmic was recorded in the rhizosphere of A. tortilis from Abu Areesh (44.6±2.5).
DISCUSSION
In the present study, the allelopathic effect of P. juliflora on seed germination, plant growth and microbe mutualisms of two native legumes (Acacia tortilis, Acacia ehrenbergiana) was studied.
Seed germination, shoot and root length and shoot and root weight are widely used parameters in allelopathy bioassays30-32. In this study, the effect of P. juliflora extract on seed germination and plant growth of A. ehrenbergiana and A. tortilis was studied in petri dish and pot experiments, respectively. The result showed that P. juliflora aqueous extracts had deleterious effects on germination of native species seeds. Seed germination and germination speed of A. ehrenbergiana and A. tortilis were greatly reduced by aqueous extracts of litter and leaves of P. juliflora. The germination inhibition increased with extract concentration as was reported by Laosinwattana et al.33. Several previous studies have suggested that the allelopathic effect of P. juliflora leaf litter was due to the presence of phenolic compounds34-36.
Growth of the seedlings of the two Acacia species was not significantly affected by the litter of P. juliflora. Similar results were reported by Shaik and Mehar37 for the effect of P. juliflora on germination and plant growth of rice. However, previous study of Alshahrani et al.38 and Mehar39 reported a higher inhibition of germination and plant growth by P. juliflora litter and leaves.
It has been reported that the increased abundance of nitrogen and sulfur caused by the decomposition of litter by soil microbes affects lipid accumulation, acetyl-CoA concentration and acetyl- CoA carboxylase activity40,41. Based on these observations, it was suggested that the negative effect of invasive plants on native species was mediated by changes in the microbial communities.
Despite the widely documented and very strong impact of P. juliflora on natives species35,37,39, few reports described the relationship between mutualisms and invasion process of P. juliflora. In this study, root nodule bacteria, mycorrhizal fungi and soil microbial biomass were compared between the invasive legume (P. juliflora) and two native species.
The most-probable-number (MPN) technique is widely used to enumerate rhizobia based upon the ability of rhizobia to nodulate appropriate host legume plants. This test was based on the assumption that organisms are randomly distributed and that one or more rhizobia are capable of causing nodulation on an appropriate host42. In the present study, MPN analysis of whole soil showed that the population density of rhizobia organisms able to nodulate P. juliflora was greater than that of the two Acacia species. The ability of P. juliflora to find compatible rhizobia in introduced regions may be an important factor in the success of the establishment of this legume in many parts of the world. This result was agreed with previous studies which show that many invasive legumes have successfully established symbiotic associations with rhizobia in introduced regions43-45. Benata et al.46 described a high diversity of bacteria that nodulate the roots of P. juliflora in the Eastern areas of Morocco. This variability in symbiont species and the abundance of nodulation could be considered a major contributing factor to the success of some legume invasive species in colonizing new areas beyond their natural range. Two scenarios are plausible, either P. juliflora formed new interactions with native rhizobia or this invasive species was co-introduced with its symbionts. Molecular identification of rhizobia in root nodules of P. juliflora should used in order to decide between the two scenarios.
The survey of root mycorrhizal status indicated that the presence of invasive plants decreases significantly the AMF colonization of roots of native species. A higher rate of mycorrhizal colonization was recorded in root of P. juliflora compared to roots of native legumes. This suggested that AMF play an important role in the establishment of the invasive legume in new habitats. Given the widespread distribution of AMF and their low-host plant specificity, mycorrhizal associations could favor invasion processes and can be important promoters of plant invasion12,44,47. It appears that AMF increase growth and competitiveness of invasive plants48. Recently De Souza and Freitas49 suggested that invasive plants were associated with specific beneficial AMF species, which give them an advantage over native species. Previous reports50-52 showed that invasive plants negatively affect native plant growth by disrupting their symbiotic associations. Invasive plants produce secondary metabolites that cause changes in soil chemical proprieties that may affect AMF community composition53,54.
In all studied sites, spore abundance decrease significantly in soil collected from under P. juliflora trees as compared to bulk soil. These results were in agreement with previous study of Barto et al.55 and Cantor et al.56 that demonstrated that invasive plants inhibited AMF hyphal growth, spore germination and also altered the AMF community. Plant invasions considerably change the diversity and abundance of soil microbial communities57-60.
The present study showed that P. juliflora affects positively the microbial biomass. The higher microbial biomass was recorded in soil collected from under P. juliflora. This could be explained by the modification of the quantity or quality of litter exerted by invasive plants as suggested by Liao and Boutton61. Previous studies suggested that invasive plants can change above ground (leaf litter) and below ground (root litter) inputs. Recently, Kuglerova et al.62 found that litter from invasive plants generally decomposed faster than from native species. In this study, P. juliflora microbial alterations were mainly driven by leaf litter produced extensively by this legume.
CONCLUSION
In the present investigation, the allelopathic effect of P. juliflora on native woody legumes in Saudi Arabia was documented for the first time. Results showed that P. juliflora possessed strong allelopathic potential on Acacia species as evidenced especially by the inhibition of seed germination. The inhibition of germination increased with extract concentration. Furthermore, P. juliflora affected soil rhizosphere microflora and altered plant-microbe mutualism, especially mycorrhizal associations. This study suggested that relationship between this invasive species and native rhizobia and AMF facilitate its establishment in new habitats and makes it’s a stronger competitor than native plant species.
SIGNIFICANCE STATEMENT
This study discovers the strong allelopathic effect of P. juliflora on native woody legumes. This study will help the researchers to uncover why invasive plants are stronger competitor than native plant species. Thus, find solutions to combat invasive plant species.
ACKNOWLEDGMENT
This study contains the results and findings of a research project that is funded by King Abdulaziz City for Science and Technology (KACST) Grant No. LGP-36-176.
REFERENCES
- Reinhart, K.O. and R.M. Callaway, 2006. Soil biota and invasive plants. New Phytol., 170: 445-457.
CrossRefDirect Link - Weidenhamer, J.D. and R.M. Callaway, 2010. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J. Chem. Ecol., 36: 59-69.
CrossRefDirect Link - Gioria, M. and B.A. Osborne, 2014. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci., Vol. 5.
CrossRefDirect Link - Carroll, S.P., A.P. Hendry, D.N. Reznick and C.W. Fox, 2007. Evolution on ecological time-scales. Funct. Ecol., 21: 387-393.
CrossRefDirect Link - Kisa, M., A. Sanon, J. Thioulouse, K. Assigbetse and S. Sylla et al., 2007. Arbuscular mycorrhizal symbiosis can counterbalance the negative influence of the exotic tree species Eucalyptus camaldulensis on the structure and functioning of soil microbial communities in a sahelian soil. FEMS Microbiol. Ecol., 62: 32-44.
CrossRefDirect Link - Remigi, P., A. Faye, A. Kane, M. Deruaz and J. Thioulouse et al., 2008. The exotic legume tree species Acacia holosericea alters microbial soil functionalities and the structure of the arbuscular mycorrhizal community. Applied Environ. Microbiol., 74: 1485-1493.
CrossRefDirect Link - Hodge, A. and A.H. Fitter, 2013. Microbial mediation of plant competition and community structure. Funct. Ecol., 27: 865-875.
CrossRefDirect Link - Traveset, A. and D.M. Richardson, 2014. Mutualistic interactions and biological invasions. Annu. Rev. Ecol. Evol. Syst., 45: 89-113.
CrossRefDirect Link - Stinson, K.A., S.A. Campbell, J.R. Powell, B.E. Wolfe and R.M. Callaway et al., 2006. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS Biol., Vol. 4.
CrossRefDirect Link - Santi, C., D. Bogusz and C. Franche, 2013. Biological nitrogen fixation in non-legume plants. Ann. Bot., 111: 742-767.
CrossRefDirect Link - Parker, M.A., W. Malek and I.M. Parker, 2006. Growth of an invasive legume is symbiont limited in newly occupied habitats. Divers. Distrib., 12: 563-571.
CrossRefDirect Link - La Pierre, K.J., E.L. Simms, M. Tariq, M. Zafar and S.S. Porter, 2017. Invasive legumes can associate with many mutualists of native legumes, but usually do not. Ecol. Evol., 7: 8599-8611.
CrossRefDirect Link - Mitchell, C.E., A.A. Agrawal, J.D. Bever, G.S. Gilbert and R.A. Hufbauer et al., 2006. Biotic interactions and plant invasions. Ecol. Lett., 9: 726-740.
CrossRefDirect Link - Rodriguez-Echeverria, S., S. Fajardo, B. Ruiz-Diez and M. Fernandez-Pascual, 2012. Differential effectiveness of novel and old legume-rhizobia mutualisms: Implications for invasion by exotic legumes. Oecologia, 170: 253-261.
CrossRefDirect Link - Valentine, A.J., M.R. Le Roux and M.A. Perez-Fernandez, 2008. Mycorrhizal colonization improves growth and nutrition of an invasive legume in a mediterranean ecosystem. Curr. Plant Sci. Biotechnol. Agric., 42: 135-136.
CrossRefDirect Link - Masrahi, Y.S., T.A. Al-Turki and O.H. Sayed, 2011. Wolffiella hyalina (Delile) Monod (Lemnaceae)-a new record for the flora of Saudi Arabia. Feddes Repertorium, 121: 189-193.
CrossRefDirect Link - Shackleton, R.T., D.C. Le Maitre, N.M. Pasiecznik and D.M. Richardson, 2014. Prosopis: A global assessment of the biogeography, benefits, impacts and management of one of the world's worst woody invasive plant taxa. AoB Plants, Vol. 6.
CrossRefDirect Link - Shackleton, R.T., D.C. Le Maitre, B.W. van Wilgen and D.M. Richardson, 2015. The impact of invasive alien Prosopis species (mesquite) on native plants in different environments in South Africa. S. Afr. J. Bot., 97: 25-31.
CrossRefDirect Link - Geesing, D., M. Al-Khawlani and M.L. Abba, 2004. Management of introduced Prosopis species: Can economic exploitation control an invasive species? Unasylva, 55: 36-44.
Direct Link - Einhelling, F.A., M.K. Schon and J.A. Rasmussen, 1982. Synergistic effects of four cinnamic acid compounds on grain sorghum. J. Plant Growth Regul., 1: 251-258.
Direct Link - Gerdemann, J.W. and T.H. Nicolson, 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244.
CrossRefDirect Link - McKenney, M.C. and D.L. Lindsey, 1987. Improved method for quantifying endomycorrhizal fungi spores from soil. Mycologia, 79: 779-782.
CrossRefDirect Link - Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Trouvelot, A., J. Kough and V. Gianinazzi-Pearson, 1986. Evaluation of VA Infection Levels in Root Systems. Research for Estimation Methods Having a Functional Significance. In: Physiological and Genetical Aspects of Mycorrhizae, Gianinazzi-Pearson, V. and S. Gianinazzi (Eds.). INRA Press, France, ISBN-13: 9782853407748, pp: 217-221.
- Amato, M. and J.N. Ladd, 1988. Assay for microbial biomass based on ninhydrin-reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem., 20: 107-114.
CrossRefDirect Link - Mardani, R., A.R. Yousefi, R. Fotovat and M. Oveisi, 2014. New bioassay method to find the allelopathic potential of wheat cultivars on rye (Secale cereale L.) seedlings. Allelopathy J., 33: 53-62.
Direct Link - Getachew, S., S. Demissew and T. Woldemariam, 2012. Allelopathic effects of the invasive Prosopis juliflora (Sw.) DC. on selected native plant species in Middle Awash, Southern Afar Rift of Ethiopia. Manage. Biol. Invasions, 3: 105-114.
Direct Link - Catalan, P., B.R. Vazquez-de-Aldana, P. de las Heras, A. Fernandez-Seral and M.E. Perez-Corona, 2013. Comparing the allelopathic potential of exotic and native plant species on understory plants: Are exotic plants better armed? Anal. Biol., 35: 65-74.
CrossRefDirect Link - Laosinwattana, C., T. Poonpaiboonpipat, M. Teerarak, W. Phuwiwat, T. Mongkolaussavaratana and P. Charoenying, 2009. Allelopathic potential of Chinese rice flower (Aglaia odorata Lour.) as organic herbicide. Allelopathy J., 24: 45-53.
Direct Link - Nakano, H., E. Nakajima, S. Hiradate, Y. Fujii, K. Yamada, H. Shigemori and K. Hasegawa, 2004. Growth inhibitory alkaloids from mesquite (Prosopis juliflora (Sw.) DC.) leaves. Phytochemistry, 65: 587-591.
CrossRefDirect Link - Metlen, K.L., E.T. Aschehoug and R.M. Callaway, 2009. Plant behavioural ecology: Dynamic plasticity in secondary metabolites. Plant Cell Environ., 32: 641-653.
CrossRefDirect Link - Ledger, K.J., R.W. Pal, P. Murphy, D.U. Nagy, R. Filep and R.M. Callaway, 2015. Impact of an invader on species diversity is stronger in the non-native range than in the native range. Plant Ecol., 216: 1285-1295.
CrossRefDirect Link - Shaik, G. and S.K. Mehar, 2015. Evaluating the allelopathic influence of mesquite (Prosopis juliflora DC.) aqueous leaf extract on the germination of rice (Oryza sativa L.) seeds using different germination indices. Int. J. Pharma Bio Sci., 6: 280-287.
Direct Link - Alshahrani, T.S., R.R. Hicks Jr., S. Verlinden and G.E. Siedel, 2009. Effects of leaf extracts of Zizyphus spina-christi and Prosopis juliflora on each others seedlings roots. Allelopathy J., 23: 111-118.
Direct Link - Mehar, S.K., 2011. Assessment of effect of Prosopis juliflora litter extract on seed germination and growth of rice. Food Sci. Qual. Manage., 2: 9-18.
Direct Link - Fazli, I.S., M.Z. Abdin, A. Jamal and S. Ahmad, 2005. Interactive effect of sulphur and nitrogen on lipid accumulation, acetyl-CoA concentration and acetyl-CoA carboxylase activity in developing seeds of oilseed crops (Brassica campestris L. and Eruca sativa Mill.). Plant Sci., 168: 29-36.
CrossRefDirect Link - Fazili, I.S., M. Masoodi, S. Ahmad, A. Jamal, J.S. Khan and M.Z. Abdin, 2010. Interactive effect of sulfur and nitrogen on growth and yield attributes of oilseed crops (Brassica campestris L. and Eruca sativa Mill.) differing in yield potential. J. Plant Nutr., 33: 1216-1228.
CrossRefDirect Link - Boakye, E.Y., I.Y.D. Lawson, S.K.A. Danso and S.K. Offei, 2016. Characterization and diversity of rhizobia nodulating selected tree legumes in Ghana. Symbiosis, 69: 89-99.
CrossRefDirect Link - Callaway, R.M., E.J. Bedmar, K.O. Reinhart, C.G. Silvan and J. Klironomos, 2011. Effects of soil biota from different ranges on Robinia invasion: Acquiring mutualists and escaping pathogens. Ecology, 92: 1027-1035.
CrossRefDirect Link - Rodriguez-Echeverria, S., J.J. Le Roux, J.A. Crisostomo and J. Ndlovu, 2011. Jack-of-all-trades and master of many? How does associated rhizobial diversity influence the colonization success of Australian Acacia species? Divers. Distrib., 17: 946-957.
CrossRefDirect Link - Wandrag, E.M., A. Sheppard, R.P. Duncan and P.E. Hulme, 2013. Reduced availability of rhizobia limits the performance but not invasiveness of introduced Acacia. J. Ecol., 101: 1103-1113.
CrossRefDirect Link - Benata, H., O. Mohammed, B. Noureddine, B. Abdelbasset and H. Abdelmoumen et al., 2008. Diversity of bacteria that nodulate Prosopis juliflora in the eastern area of Morocco. Syst. Applied Microbiol., 31: 378-386.
CrossRefDirect Link - Pringle, A., J.D. Bever, M. Gardes, J.L. Parrent, M.C. Rillig and J.N. Klironomos, 2009. Mycorrhizal symbioses and plant invasions. Annu. Rev. Ecol. Evol. Syst., 40: 699-715.
CrossRefDirect Link - Lekberg, Y., S.M. Gibbons, S. Rosendahl and P.W. Ramsey, 2013. Severe plant invasions can increase mycorrhizal fungal abundance and diversity. ISME J., 7: 1424-1433.
CrossRefDirect Link - De Souza, T.A.F. and H. Freitas, 2017. Arbuscular mycorrhizal fungal community assembly in the Brazilian tropical seasonal dry forest. Ecol. Process., Vol. 6.
CrossRefDirect Link - Zubek, S., J. Blaszkowski, K. Seidler-Lozykowska, W. Baba and P. Mleczko, 2013. Arbuscular mycorrhizal fungi abundance, species richness and composition under the monocultures of five medicinal plants. Acta Sci. Pol.: Hortorum Cultus, 12: 127-141.
Direct Link - Zubek, S., M.L. Majewska, J. Blaszkowski, A.M. Stefanowicz, M. Nobis and P. Kapusta, 2016. Invasive plants affect arbuscular mycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biol. Fertil. Soils, 52: 879-893.
CrossRefDirect Link - De Souza, T.A.F., S. Rodriguez-Echeverria, L.A. de Andrade and H. Freitas, 2016. Could biological invasion by Cryptostegia madagascariensis alter the composition of the arbuscular mycorrhizal fungal community in semi-arid Brazil? Acta Bot. Bras., 30: 93-101.
CrossRefDirect Link - Shah, M.A., Z.A. Reshi and D.P. Khasa, 2009. Arbuscular mycorrhizas: Drivers or passengers of alien plant invasion. Bot. Rev., 75: 397-417.
CrossRefDirect Link - Da Silva, I.R., C.M.A. de Mello, R.A.F. Neto, D.K.A. da Silva, A.L. de Melo, F. Oehl and L.C. Maia, 2014. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Applied Soil Ecol., 84: 166-175.
CrossRefDirect Link - Barto, E.K., P.M. Antunes, K. Stinson, A.M. Koch, J.N. Klironomos and D. Cipollini, 2011. Differences in arbuscular mycorrhizal fungal communities associated with sugar maple seedlings in and outside of invaded garlic mustard forest patches. Biol. Invasions, 13: 2755-2762.
CrossRefDirect Link - Cantor, A., A. Hale, J. Aaron, M.B. Traw and S. Kalisz, 2011. Low allelochemical concentrations detected in garlic mustard-invaded forest soils inhibit fungal growth and AMF spore germination. Biol. Invasions, 13: 3015-3025.
CrossRefDirect Link - Hejda, M., P. Pysek and V. Jarosik, 2009. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol., 97: 393-403.
CrossRefDirect Link - Moron, D., M. Lenda, P. Skorka, H. Szentgyorgyi, J. Settele and M. Woyciechowski, 2009. Wild pollinator communities are negatively affected by invasion of alien goldenrods in grassland landscapes. Biol. Conserv., 142: 1322-1332.
CrossRefDirect Link - Aguilera, A.G., P. Alpert, J.S. Dukes and R. Harrington, 2010. Impacts of the invasive plant Fallopia japonica (Houtt.) on plant communities and ecosystem processes. Biol. Invasions, 12: 1243-1252.
CrossRefDirect Link - Lenda, M., M. Witek, P. Skorka, D. Moron and M. Woyciechowski, 2013. Invasive alien plants affect grassland ant communities, colony size and foraging behaviour. Biol. Invasions, 15: 2403-2414.
CrossRefDirect Link - Liao, J.D. and T.W. Boutton, 2008. Soil microbial biomass response to woody plant invasion of grassland. Soil Biol. Biochem., 40: 1207-1216.
CrossRefDirect Link - Kuglerova, L., L. Garcia, I. Pardo, Y. Mottiar and J.S. Richardson, 2017. Does leaf litter from invasive plants contribute the same support of a stream ecosystem function as native vegetation? Ecosphere, Vol. 8, No. 4.
CrossRefDirect Link