
Olfat Malak Wahbi
Department of Fish Reproduction and Aquacultural, National Institute of Oceanography and Fisheries, Aquaculture Division, El-Anfousy, Kayetbay, Alexandria, Egypt
LiveDNA: 20.16715
Zeinab Abdel Baki El-Greisy
Department of Fish Reproduction and Aquacultural, National Institute of Oceanography and Fisheries, Aquaculture Division, El-Anfousy, Kayetbay, Alexandria, Egypt
LiveDNA: 20.19539
Heba Saad El-Sayed
Department of Fish Reproduction and Aquacultural, National Institute of Oceanography and Fisheries, Aquaculture Division, El-Anfousy, Kayetbay, Alexandria, Egypt
LiveDNA: 20.16883
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 6 | Page No.: 278-287







ABSTRACT
Background and Objective: The routine technique applied in spawning of sea bream (Sparus aurata), in the hatchery depends on the use of a mixture of synthetic luteinizing hormone releasing hormone (LHRH-a)+human chorionic gonadotropin hormone (HCG). The present study aims to use HCG alone as a method to take the advantages of HCG specifically lower production costs. Materials and Methods: Adult and fully ripe Sparus aurata were injected with either one of two hormone protocols using two hormones, human chorionic gonadotropin hormone (HCG) and a mixture of luteinizing hormone releasing hormone analogue and HCG (LHRH-a+HCG) during the spawning season. The two hormone protocols were applied through a single dose of each hormone. The first protocol was applied by 5000 IU HCG kg–1 and the second protocol was applied by using 5000 IU HCG+200 μg LHRH-a kg–1. A comparison between the two protocols is demonstrated in the present study. Forty four ripe fish (16♀+28♂) were divided into 2 groups each of (8♀+14♂) and were injected intramuscularly with a single dose of human chorionic gonadotropin hormone (HCG 5000 IU kg–1) or mixture of luteinizing hormone releasing hormone [LHRH-a (200 μg kg–1)+HCG (5000 IU kg–1)]. Data were analyzed using overall one-way analysis of variance (ANOVA) and differences were observed at significance of p<0.05. Results: In the females of the two experimental groups, spawning was induced in 54 and 48 h after injection in the mixture and the single hormone protocol, respectively. A comparison between both protocols showed that both treatments were found to be equipotent in inducing fish spawning with superiority of LHRH-a+HCG mixture over HCG. Fertilization and hatchability rates as well as eggs and larval quality recorded increased values in group injected with hormones mixture. Although no histological differences were seen in pituitary section of the 2 groups, the number of gonadotrophic cells and vacuoles diameter/unit area showed differences between the two groups. The correlation value and F-test showed that there are not any significant differences between the values of survival percentages, average length and average weight in cases of HCG alone or LHRH-a+HCG. Conclusion: Even though the superiority of LHRH-a+HCG mixture over HCG but for economical purpose, the use of HCG alone is preferred.
PDF Abstract XML References Citation
Received: June 18, 2017;
Accepted: September 08, 2017;
Published: February 23, 2019
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Olfat Malak Wahbi, Zeinab Abdel Baki El-Greisy and Heba Saad El-Sayed, 2017. Reproductive Performance of Sea Bream (Sparus aurata) Induced with Two Different Hormone Protocols with Respect to the Effect on Gonadotrophic Cells. Journal of Biological Sciences, 17: 278-287.
DOI: 10.3923/jbs.2017.278.287
URL: https://scialert.net/abstract/?doi=jbs.2017.278.287
DOI: 10.3923/jbs.2017.278.287
URL: https://scialert.net/abstract/?doi=jbs.2017.278.287
INTRODUCTION
Eventhough the natural spawning is the preferred method for breeding cultivated fish, the induced spawning using sexual hormones may be necessary to control timing and synchrony of egg production due to environmental and practical reasons. Choosing a successful synthetic hormone involves the selection of effective hormone formulations, proper duration of hormonal treatment and timing of the hormone administration1. Hormonal inductions are important in aquaculture that can improve final maturation and ovulation. Induced ovulation protocols in marine fish most commonly utilize human chorionic gonadotropin (HCG) or luteinizing hormone releasing hormone analogues (LHRH-a). LHRH-a stimulates the release of endogenous gonadotropin (GtH), whereas, HCG mimics endogenous GtH2.
Human chorionic gonadotropin (HCG) has been widely used since the 1930s to induce ovulation and spawning3 and to increase milt volume in various species of aquaculture fish. HCG offers major advantages, it is relatively inexpensive and is of known and consistent potency, more stable and thus has a longer shelf life and it has been available in clinical grade, standardized preparations throughout the world. For the purpose of induced spawning, it is only effective in fish that are already sexually mature. The HCG has been employed in spawning induction trials of many species in culture4-6. In some species, HCG has a low bio-potency and high doses or multiple injections are required, this has led to a more widespread use of LHRH-a2.
Injection of sexually mature females of black bream with LHRH-a has resulted in a better ovulatory response than the treatment with HCG7. However, HCG has been successfully used to induce ovulation in several sparids including gilthead sea bream8 and black bream7.
The use of synthetic luteinizing hormone releasing hormone (LHRH-a) or human chorionic gonadotropin hormone (HCG) has been shown to induce ovulation and spawning in several commercially important fish9-12.
In the present study, two intramuscular injection regimes were applied for the fish. The first was applied by injection of HCG (5000 IU kg–1). The second was applied by injection of two hormones [LHRH-a (200 μg)+HCG (5000 IU kg–1)]. A comparison between both regimes was demonstrated since the development of local economic fish breeds such as Sparus aurata is important for the Egyptian economy. The authors in the present study aim to compare their routine technique applied in spawning of sea bream in the hatchery which depends on use of a mixture of synthetic luteinizing hormone releasing hormone (LHRH-a)+human chorionic gonadotropin hormone (HCG), to the use of HCG alone. This method uses the advantages of the potential of HCG specifically lower production costs and high seed production level.
MATERIALS AND METHODS
Broodstock: About forty four ripe Sparus aurata brood fish (16 females and 28 males) were brought from Damietta in December, 2016 to the marine hatchery of the National Institute of Oceanography and Fisheries (NIOF), Alexandria, Egypt. The average total length of the females was 27±2 and 24±1 cm for males. The average total weight was 415±5 g for females and was 315±3 g for males.
The environmental factors, such as temperature and photoperiod were maintained at the optimum conditions required. The fish were held in tanks filled with sea water (32-35%) over 10 days before hormonal treatment with a temperature ranged from 15-17°C.
The females were characterized by the swelling of the abdomen and the protrusion of the genital papilla. Ripening was checked by applying gentle pressure to the abdomen of female broodstock at intervals.
Experimental design and hormone injection: Adult and fully ripe Sparus aurata were injected with either one of two hormone protocols involving two hormones, human chorionic gonadotropin hormone (HCG) and luteinizing hormone releasing hormone analogue (LHRH-a+HCG) during the spawning season. The two hormone protocols were applied through a single dose of each hormone. The first protocol was applied by using 5000 IU HCG kg–1 and the second protocol was applied by using 5000 IU HCG +200 μg LHRH-a kg–1.
After injection, the females were transferred with spermiating males to the spawning tanks provided with flow-through seawater and aeration with sex ratio nearly 2:1males to females to optimize the fertilization rate. The fish started to spawn after 48 h for the group involved the mixture of hormones and 54 h for the group using HCG only.
Collection of eggs: After spawning, the fertilized eggs were collected from the spawning tanks with a fine mesh size dip net to hatching fiberglass tanks. At day 7, the fry moved to the larval rearing tanks (8 tanks each 2 m3). The stocking density of fry was 100000 fry/tank. Eight round indoor tanks (2 m3) were used for each treatment.
Feeding regime: Alga (Nannochloropsis salina) and Rotifer (Brachionus plicatilis) were introduced to the larvae of the two groups after 3-4 days after hatch (DAH) at rate of 5 ind mL–1. Rotifer was given daily at rate of 10-15 ind mL–1 from the 7th-9th DAH and 15-20 ind mL–1 from the 10th-20th DAH. The algae serve as a food for rotifer and help in improving water quality. Every day before changing the water, the remaining rotifer in the rearing tanks is counted in order to adjust the amount of rotifer given daily. To ensure that the availability of food is adequate for the fry during the 24 h period, the amount of rotifer remaining in the tank at the following day should not less than 5 ind mL–1. Starting from day 20, rotifer was decreasing gradually with trying the addition of Artemia nauplii (0.1 pcs mL–1).
Enriched Artemia metanauplii with DHA-Selco (INVE product) was given when the larvae reached the 30th-45th day old. The density of Artemia given is 0.2 pcs mL–1. The density was gradually increased to 0.5-1 pcs mL–1 when the fry were 35 days old and 1-1.5 pcs mL–1 for the 35-40 days old fry, then it increased to 1.5-2 pcs mL–1 from day 40th-45th. At day 40, the fry began to be fed with artificial dry food of 60% crude protein. In days 45-50, the diet amount was 2 g m–3 and on days 55-60, the diet amount was 5 g m–3. The actual amount of food required/day should be adjusted by observing the eating and swimming behavior of the fry.
Histological observations: The fish injected with either HCG or LHRH-a+HCG mixture hormones were killed by decapitation, their pituitary with brain were removed and fixed in Bouin’s fixative for 24 h at 4°C, then dehydrated through graded ethanol solution, cleared and embedded in wax (MP:56-58°C). Consecutive median sagittal sections of the gland were made at 4 μm thickness and stained with hematoxylin, eosin and microscopically examined for any histological variations between groups. To compare the activity of gonadotropin cells in pituitary of the two groups, the numbers of GTH cells/unit area (4.2 mm2) were counted. Five different areas in the sagittal section of the central proximal pars distalis were selected at random in both sexes and the number of GTH cells was counted on the photomicrographs and scoring was repeated for 2 fish/sex/group. In the same way vacuoles diameter were measured and compared in both hormones injected groups.
Statistical analysis: Statistical tests were performed using Microsoft Excel 2010. Data were represented as mean±standard deviation. Data were analyzed using overall one-way analysis of variance (ANOVA)13 and when differences observed were significant at p<0.05.
RESULTS
Reproductive parameters: The reproductive parameters of the fish showed that the time lapsed to the first spawning patch were 54 h in HCG group and 48 h in LHRH-a+HCG group. The diameter of the fertilized eggs was higher in case of injection with LHRH-a+HCG (900±50 μm) than in case of HCG injection (850±50 μm). The number of egg spawned per the group in case of injection with LHRH-a+HCG was (5.2×106) higher than in case of HCG injection (4.7×106). The fertilization and hatching rates was higher in case of LHRH-a+HCG than in case of HCG injection (Table 1).
Survival percentage of the larvae: The relation of average survival percentage of sea bream larvae during both of rotifer feeding period and Artemia feeding and weaning period showed that they have the same trend with superiority of LHRH-a+HCG mixture over HCG (Fig. 1 and 2). Also, the correlation and F-test showed that there are not any significant differences between HCG and LHRH-a+HCG mixture during rotifer feeding period (Table 2 and 3) and during Artemia and weaning period (Table 4 and 5).
Length of the larvae: The average length of the sea bream larvae during Rotifer feeding period and Artemia feeding and weaning period showed the same trend with superiority of LHRH-a+HCG mixture over HCG (Fig. 3 and 4).
The values of correlation and F-test of the HCG and LHRH-a+HCG mixture exhibited no significant differences between them during rotifer feeding period (Table 6 and 7) and Artemia feeding and weaning period (Table 8 and 9).
Weight of the larvae: Data in Fig. 5 and 6 showed superiority of LHRH-a+HCG mixture over HCG in the weight of sea bream larvae during the two periods.
| Table 1: | Reproductive parameters of Sparus aurata in the two different induction protocols |
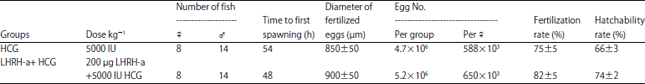 | |
| Mean±Standard Deviation | |
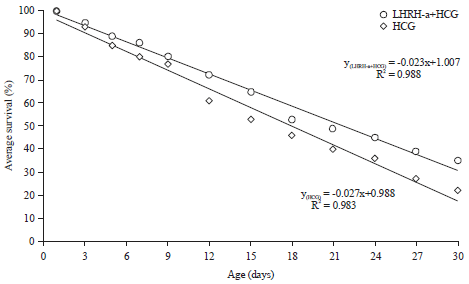 | |
| Fig. 1: | Average survival percentage of sea bream larvae during rotifer feeding period |
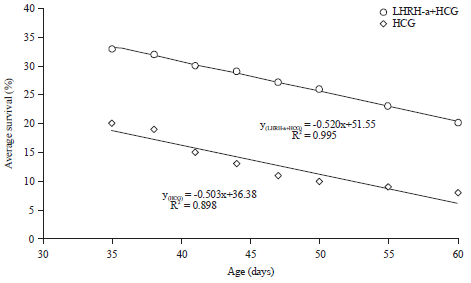 | |
| Fig. 2: | Average survival percentage of sea bream larvae during Artemia and weaning feeding period |
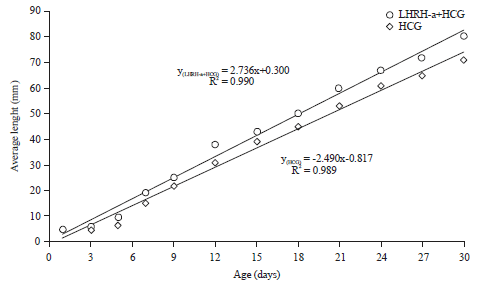 | |
| Fig. 3: | Average length (mm) of sea bream larvae during rotifer feeding period |
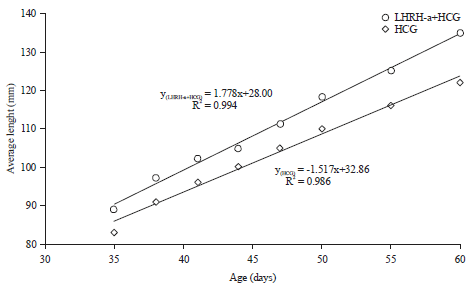 | |
| Fig. 4: | Average length (mm) of sea bream larvae during Artemia and weaning feeding period |
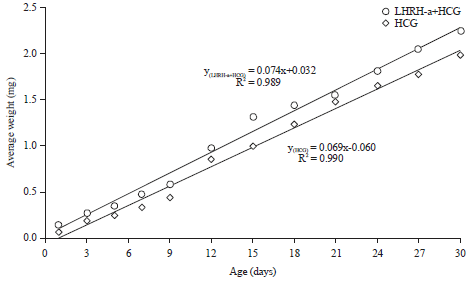 | |
| Fig. 5: | Average weight (mg) of sea bream larvae during rotifer feeding period |
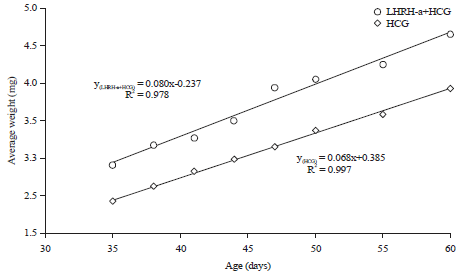 | |
| Fig. 6: | Average weight (mg) of sea bream larvae during Artemia and weaning feeding period |
| Table 2: | Regression statistics between HCG and (LHRH-a+HCG) survival percentages of sea bream larvae during rotifer feeding period |
 | |
| Table 3: | ANOVA between HCG and (LHRH-a+HCG) survival percentages of sea bream larvae during rotifer feeding period |
 | |
| Table 4: | Regression statistics between HCG and (LHRH-a+HCG) survival percentages of sea bream larvae during Artemia and weaning feeding period |
 | |
| Table 5: | ANOVA between HCG and (LHRH-a+HCG) survival percentages of sea bream larvae during Artemia and weaning feeding period |
 | |
| Table 6: | Regression statistics between HCG and (LHRH-a+HCG) length of sea bream larvae during rotifer feeding period |
 | |
| Table 7: | ANOVA between HCG and (LHRH-a+HCG) length of sea bream larvae during rotifer feeding period |
 | |
| Table 8: | Regression statistics between HCG and (LHRH-a+HCG) length of sea bream larvae during Artemia feeding and weaning period |
 | |
Also, correlation and F-test showed that there are not any significant differences between HCG and LHRH-a+HCG mixture during rotifer feeding period (Table 10 and 11) and Artemia and weaning feeding period (Table 12 and 13).
| Table 9: | ANOVA between HCG and (LHRH-a+HCG) length of sea bream larvae during Artemia feeding and weaning period |
 | |
| Table 10: | Regression statistics between HCG and (LHRH-a+HCG) weight of sea bream larvae during rotifer feeding period |
 | |
| Table 11: | ANOVA between HCG and (LHRH-a+HCG) weight of sea bream larvae during rotifer feeding period |
 | |
| Table 12: | Regression statistics between HCG and (LHRH-a+HCG) weight of sea bream larvae Artemia and weaning feeding period |
 | |
| Table 13: | ANOVA between HCG and (LHRH-a+HCG) weight of sea bream larvae during Artemia and weaning feeding period |
 | |
Gonadotrophic cells of broodstock: In the present study, numbers of GTH cells/unit area in the proximal pars distalis were counted to examine the effect of hormonal application on gonadotropic cells activity. The GTH cells/unit area in the LHRH-a+HCG hormone mixture injected group were between 44-46.6 in both sex and in HCG hormone injected group were between (46.3-51.9 in both sex) (Fig. 7). Also vacuoles diameter ranged between 13.0-14.6 in ♂ and ♀, respectively in the LHRH-a+HCG hormone mixture injected group and between 12.5 -13.1in ♂ and ♀, respectively in HCG hormone injected group (Fig. 7). Histological observations of sea bream pituitary showed no structural variations between the two groups. The gonadotropic cells are granulated cells with variable shape and size, vacuolization presents a net or sieve like appearance. The gonadotropic cells in both treatments were hypertrophied and showed difference in staining affinity (Fig. 8 and 9).
 | |
| Fig. 7(a-b): | Gonadotrophic cells numbers and vacuoles diameters of sea bream broodstock/unit area in the proximal pars distalis |
DISCUSSION
Many sea bream fish reared under controlled conditions exhibit signs of reproductive dysfunction, which is manifested in the failure to undergo final oocyte maturation and ovulation. In captivity a disruption of endogenous gonadotropin releasing hormone stimulation occurs and sequentially that of luteinizing hormone, which is indispensible for the final phases of gametogenesis. In addition to methods based on the application of exogenous gonadotropins, the usage of a method functioning on the basis of hypothalamic control of final oocyte maturation and ovulation has become popular recently.
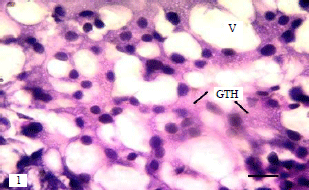 | |
| Fig. 8: | Sagittal section of pituitary of ovulating ♀ from (LHRH-a+HCG) injected group, note hypertrophied gonadotrophs (GTH) with high staining affinity and large vacuoles, bar 20 μm |
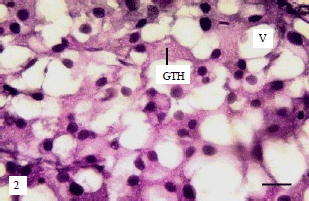 | |
| Fig. 9: | Sagittal section of pituitary of ovulating ♀ from (HCG) injected group, showing hypertrophied gonadotrophs (GTH), enlarged vacuoles (V), bar 20 μm |
The replacement of natural gonadotropin-releasing hormones with chemically synthesized gonadotropin-releasing hormone analogues characterized by amino acid substitutions at positions sensitive to enzymatic degradation has resulted in a centuple increase in the effectiveness of luteinizing hormone secretion induction.
Hormonal stimulation of final oocyte maturation and ovulation has, for decades now, been an important aid in the effective reproduction of a majority of economically important species. The development of hormone stimulators first took in gonadotropic hormones found in carp pituitary, choriogonadotropins, through to currently preferred synthetic GnRH analogues.
The sea bream is a protandrous hermaphrodite. It is a functional male in the first 2 years and at over 30 cm in length becomes female. During the male phase, the bisexual gonad has functional testicular, with asynchronous spermatogenesis and non-functional ovarian areas14,15. Ovarian development is also asynchronous and females are batch spawners that can lay 20000-80000 eggs/day for a period of up to 3 months. The eggs are spherical and pelagic, with a diameter slightly lower than1 mm and a single large oil droplet. The planktonic larval stage lasts about 50 days at a temperature range of 17-18°C.
Treatment with LHRH-a+HCG mixture or HCG was approximately equivalent in terms of the number of fish that ovulated, the number of repeat ovulations and egg quality and fertility. Haddy and Pankhurst7 reported that HCG and LHRH-a both successfully induced repeat ovulations in black bream. The proportions of black bream ovulating and the number of repeat ovulations were highest in individuals injected with LHRH-a, that is in agreement with the present study. Zohar et al.16 on inducing spawning of gilthead sea bream found that female treated with LHRH-a spawned for longer periods than those treated with HCG. Similarly in black bream, multiple ovulations can be induced by single intraperitoneal injection of LHRH-a in dose of 50 μg kg–1 7. Poortenaar and Pankhurst17, showed similar results on inducing spawning of greenback flounder treated with HCG, LHRH hormone (50 μg kg–1 ipi). On contrary, Yaron et al.18 and Cleary et al.19 suggested that direct administration of GTH may be more effective than stimulation of a natural GTH flow via administration of LHRH-a and the difference in response to HCG and LHRH-a hormones is likely to result from differences in the mode of action of the two hormones in inducing oocyte maturation and ovulation.
In present study, a dose of 5000 IU kg–1 HCG induced repeated spawning in gilthead sea bream Sparus aurata. Treatment of other sparids with HCG has also yield variable results, a dose of 1000 IU kg–1 induced multiple spawns in yellow fin bream (Acanthopagrus australis20), black bream (Acanthopagrus butcheri) injected with 1000 IU kg–1 HCG induce multiple ovulation over 5 days7. The sheepshead bream (Puntazzo puntazzo) can be induced to spawn repeatedly by intramuscular injection of 1000 IU kg–1 HCG21.
The present study showed an increase in oocyte diameter in response to hormonal treatments. Oocytes diameter increased significantly and oocyte stage advanced significantly in greenback flounder (Rhombosolea tapirina) in response to exogenous hormone treatment (HCG, LHRH-a)17. A single injection dose of 1000-3000 IU kg–1 HCG induced ovulation in trout cod (Maccullochella macquariensis22).
Channa striatus implanted with HCG for a period of 5 months induced a significant increase in ova diameter and fecundity23.
Sahoo et al.3 recorded a fertilization rate of 75-86% in catfish at 3000-5000 IU doses of HCG with 14-17 h post injection. In this study fertilization rate of 70-80% obtained in sea bream at 5000 IU doses of HCG after 48-54 h post injection.
In the present study, the GTH cells in LHRH-a+HCG hormones mixture injected group were hypertrophied, with high staining affinity and large vacuoles. Similarly Matsuyama et al.24,25, showed hypertrophied gonadotropic cells with large vacuoles in induced spawning of red sea bream injected with LHRH-a . The number of GTH cells/unit area in the PPD decrease and vacuoles diameter increases in fish treated with LHRH-a+HCG hormones mixture indicating an increase in cell size than HCG hormonal injection. Van Overbeeke and McBride26 stated that upon maturation of the GTH cells its size and staining affinity increases.
Eventhough the superiority of LHRH-a+HCG mixture over HCG hormone, but for economical purpose, the use of HCG hormone alone is preferred.
Further research aimed at the evaluation of more potent gonadotropin along with a detailed search for the reasons of reproductive dysfunction should contribute to future progress in the area of artificial stimulation of final oocyte maturation and ovulation in sea bream.
CONCLUSION
All females in the two experimental groups treated with either one of two hormone injection protocols using two hormones, human chorionic gonadotropin hormone (HCG) and a mixture of luteinizing hormone releasing hormone analogue and human chorionic gonadotropic hormone (LHRH-a+HCG) during the spawning season caused the females to start spawning after 48 and 54 h post injection. However both treatments were effective at inducing sea bream females to spawn in captivity under hatchery conditions. It seems also that the doses of both hormones are enough and equipotent to induce spawning in Sparus aurata with lower production costs in case of HCG injection.
SIGNIFICANCE STATEMENT
This study aims to discover the probability of reducing the production costs of induced spawning of sea bream (Sparus aurata). It could help the workers in the hatchery to replace the routine technique using a mixture of luteinizing hormone releasing hormone (LHRH-a) and human chorionic gonadotropin hormone (HCG) with another technique using only HCG to reduce the production costs and use the potential of HCG while maintaining the high seed production level.
REFERENCES
- Elakkanai, P., T. Francis, B. Ahelan, P. Jawahar, P. Padmavathy, N. Jayakumar and A. Subburaj, 2015. Role of GnRH, HCG and kisspeptin on reproduction of fishes. Indian J. Sci. Technol., 8: 1-10.
CrossRefDirect Link - Donaldson, E.M. and R.H. Devlin, 1996. Uses of Biotechnology to Enhance Production. In: Principles of Salmonid Culture (Developments in Aquaculture and Fisheries Science, Volume 29), Pennell, W. and B.A. Barton (Eds.). Chapter 17, Elsevier, Amsterdam, Netherland, ISBN-13: 9780080539669, pp: 969-1020.
Direct Link - Sahoo, S.K., S.S. Giri, S. Chandra and A.K. Sahu, 2007. Spawning performance and egg quality of Asian catfish Clarias batrachus (Linn.) at various doses of human chorionic gonadotropin (HCG) injection and latency periods during spawning induction. Aquaculture, 266: 289-292.
CrossRefDirect Link - Zohar, Y. and C.C. Mylonas, 2001. Endocrine manipulations of spawning in cultured fish: From hormones to genes. Aquaculture, 197: 99-136.
CrossRefDirect Link - Zakes, Z. and K. Demska-Zakes, 2005. Artificial spawning of pikeperch (Sander lucioperca (L.)) stimulated with Human Chorionic Gonadotropin (HCG) and mammalian GnRH analogue with a dopamine inhibitor. Arch. Polish Fish., 13: 63-75.
Direct Link - Minar, M.H., M. Shamsuddin, M.G.U. Bablu and S.I. Bhuyan, 2012. Induced spawning practices of different fishes in the hatcheries of Barisal district, Bangladesh. Trends. Fish. Res., 1: 14-17.
Direct Link - Haddy, J.A. and N.W. Pankhurst, 2000. The efficacy of exogenous hormones in stimulating changes in plasma steroids and ovulation in wild black bream Acanthopagrus butcheri is improved by treatment at capture. Aquaculture, 191: 351-366.
CrossRefDirect Link - Zohar, Y. and H. Gordin, 1979. Spawning kinetics in the gilthead sea-bream, Sparus aurata L. after low doses of human chronic gonadotropin. J. Fish Biol., 15: 665-670.
CrossRefDirect Link - Kucharczyk, D., R. Kujawa, A. Mamcarz, K. Targonska-Dietrich and E. Wyszomirska et al., 2005. Induced spawning in bream (Abramis brama L.) using pellets containing GnRH. Czech J. Anim. Sci., 50: 89-95.
Direct Link - Legendre, M., J. Slembrouck, J. Subagja and A.H. Kristanto, 2000. Ovulation rate, latency period and ova viability after GnRH- or hCG-induced breeding in the Asian catfish Pangasius hypophthalmus (Siluriformes, Pangasiidae). Aquat. Living Resour., 13: 145-151.
CrossRefDirect Link - Adebayo, O.T. and O.A. Fagbenro, 2004. Induced ovulation and spawning of pond raised African giant catfish, Heterobranchus bidorsalis by exogenous hormones. Aquaculture, 242: 229-236.
CrossRefDirect Link - Zohar, Y., M. Abraham and H. Gordon, 1978. The gonadal cycle of the captivity-reared hermaphroditic teleost Sparus aurata (L.) during the first two years of life. Ann. Biol. Biochem. Biophys., 18: 877-882.
Direct Link - Poortenaar, C.W. and N.W. Pankhurst, 2000. Effect of luteinising hormone-releasing hormone analogue and human chorionic gonadotropin on ovulation, plasma and ovarian levels of gonadal steroids in greenback flounder Rhombosolea tapirina. J. World Aquacult. Soc., 31: 175-185.
CrossRefDirect Link - Yaron, Z., G. Gur, P. Melamed, H. Rosenfeld, A. Elizur and B. Levavi-Sivan, 2003. Regulation of fish gonadotropins. Int. Rev. Cytol., 225: 131-185.
CrossRefDirect Link - Cleary, J.J., S.C. Battaglene and N.W. Pankhurst, 2002. Capture and handling stress affects the endocrine and ovulatory response to exogenous hormone treatment in snapper, Pagrus auratus (Bloch & Schneider). Aquacult. Res., 33: 829-838.
CrossRefDirect Link - Black, B.J. and M. Black, 2013. Efficacy of two exogenous hormones (GnRHa and hCG) for induction of spontaneous spawning in captive yellowfin bream, Acanthopagrus australis (Sparidae) and influence of sex ratio on spawning success. Aquaculture, 416: 105-110.
CrossRefDirect Link - Faranda, F., A. Cavaliere, G.L. Paro, A. Manganaro and A. Mazzola, 1985. Preliminary studies on reproduction of Puntazzo puntazzo (Gmelin 1789) (Pisces, Sparidae) under controlled conditions. Aquaculture, 49: 111-123.
CrossRefDirect Link - Ingram, B.A. and M.A. Rimmer, 1993. Induced breeding and larval rearing of the endangered Australian freshwater fish trout cod, Maccullochella macquariensis (Cuvier) (Percichthyidae). Aquacult. Res., 24: 7-17.
CrossRefDirect Link - Selvaraj, S. and T. Francis, 2007. Influence of human chorionic gonadotropin on maturation in striped murrel, Channa striatus. Asian Fish. Sci., 20: 23-39.
Direct Link - Matsuyama, M., H. Kagawa, T. Ashitani, M. Kashiwagi, T. Iwai, K. Okuzawa and H. Tanaka, 1993. Development of LHRH-a copolymer pellet polymerized by ultraviolet and its application for maturation in red sea bream Pagrus major during the non-spawning season. Nippon Suisan Gakkaishi, 59: 1361-1369.
CrossRefDirect Link - Matsuyama, M., H. Takeuchi, M. Kashiwagi, K. Hirose and H. Kagawa, 1995. Induced gonadal development and spawning of immature red sea bream Pagrus major with LHRH-a administration in different ways during winter season. Fish. Sci., 61: 472-477.
Direct Link - Van Overbeeke, A.P. and J.R. McBride, 1967. The pituitary gland of the sockeye (Oncorhynchus nerka) during sexual maturation and spawning. J. Fish. Board Can., 24: 1791-1810.
CrossRefDirect Link