
Manal Ibrahim El-Barbary
Laboratory of Fish Disease, National Institute of Oceanography and Fisheries, Alexandria, Egypt
LiveDNA: 20.15073
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 5 | Page No.: 222-234







ABSTRACT
Background and Objective: Bacterial pathogens are the causative agents of most serious disease problems in both wild and cultured fish causing mortalities and severe economic losses. In spring 2016, mass mortality occurred in some fish farms with different salinity levels in Damietta governorate, Egypt. This study aimed to isolate and characterize pathogenic Aeromonas hydrophila (A. hydrophila) that naturally infect Oreochromis niloticus (O. niloticus) and Sparus aurata (S. aurata). Materials and Methods: Sixteen presumptive motile Aeromonas were isolated from samples of liver and kidney of O. niloticus and S. aurata on selective medium. Some isolates were identified by API 20NE and 16S rRNA, their phenotypes and multiple antibiotic resistance (MAR index); in addition, blood chemistry and histological changes in the liver, kidney and spleen of O. niloticus were investigated after infection with A. hydrophila. Results: Isolates, aer114 and aer8 were identified by 16S rRNA and submitted in GenBank under the accession numbers LC208788 and LC208789 and they exhibited degrees of virulence (71.4 -76.2%) for O. niloticus post intraperitoneal injected which showed significant increase (p<0.05) in the activity of serum AST and ALT, creatinine and urea levels but significant decreases (p<0.05) in total protein, albumin and globulin concentrations. The major histopathologic findings of infected O. niloticus were lipid vacuoles, pyknotic nuclei and necrosis in hepatocytes, while kidney showed necrosis area and hemorrhage between renal tubules but spleen showed necrosis and infiltration of fibroblasts around damage blood vessels and hemolytic between red pulps. In addition, the tested isolates showed resistant patterns of 66.6% (16/18) for many antibiotics (18), while MAR index was 0.16. Conclusion: It was concluded that the negative effects of aer114 isolated from O. niloticus on organs histology and blood examinations were more than the isolate aer8 that was isolated from Sparus aurata.
PDF Abstract XML References Citation
Received: June 18, 2017;
Accepted: August 24, 2017;
Published: September 12, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Manal Ibrahim El-Barbary, 2017. Serum Biochemical and Histopathological Changes Associated with Aeromonas hydrophila Isolated from Oreochromis niloticus and Sparus aurata with Multiple Antibiotic Resistance Index. Journal of Biological Sciences, 17: 222-234.
DOI: 10.3923/jbs.2017.222.234
URL: https://scialert.net/abstract/?doi=jbs.2017.222.234
DOI: 10.3923/jbs.2017.222.234
URL: https://scialert.net/abstract/?doi=jbs.2017.222.234
INTRODUCTION
Bacterial pathogens are the causative agents for serious disease problems in wild and farmed fish causing mortalities and severe economic losses. Aeromonas species is classified into two main groups; psychrophilic nonmotile bacteria1,2 or mesophilic motile bacteria3 that belong to family Aeromonadaceae4.
Motile Aeromonas sp. are Gram-negative rods, oxidase positive, facultative anaerobic, opportunistic pathogens and causative agents of motile aeromonas septicemia (MAS) which cause various diseases in fish where they are largely distributed in aquatic environments and they can be easily found in water rich in organic matter, so stress and poor water quality play a major role in their occurrence5,6. Since early 1960s A. hydrophila has been known as a pathogen of fish in Europe7 and in USA8, also, A. hydrophila is responsible for hemorrhagic septicemia9,10. The symptoms of A. hydrophila infection include swelling of tissues, ascites, necrosis, ulcers and hemorrhage, exophthalmia, redness of the skin and fluid accumulation in the pockets scale11-13. Since A. hydrophila is commonly found in aquatic habitats, it is a secondary invader of injured tissue where it causes disease in the stressed fish14, A. hydrophila can also become a primary pathogen, causing outbreaks in fish farms with high mortality rates, which leads to severe economic losses to the aquaculture sector worldwide15,16 of either freshwater or marine water fish17. A. hydrophila is often found in association with disease outbreaks in aquaculture production in diseased fish and crabs18. A. hydrophila was isolated from the cultured fish that exposed to numerous stress factors such as handling, crowding, contaminants and physiological changes, which may lead to immune depression and outbreaks of infections in aquaculture19, where water pollution increases susceptibility to infectious diseases, leading to higher mortality rates20 because any change in water parameters leads to conditions that allow bacterial infections to take place21.
In Egypt, A. hydrophila has been isolated from various fish species and water salinity such as Groper, Sea bass, Mullet, Tilapia, Carp and some ornamental fish species22,23. Several studies reveled that A. hydrophila caused histological changes in tissues and organs of fish infected22,24,25, where A. hydrophila indicated liver and kidney damage26. That can be attributed to the presence of many extracellular toxins and enzymes such as cytotoxins, hemolysins, enterotoxins and proteases that are excreted from the virulence of A. hydrophila22,27.
On the other hand, previous studies have reported that A. hydrophila caused significant disorders of blood parameters in infected fish25,28,29 where many parameters such as ALT, AST, creatinine and urea are vital indicators of any dysfunction in the liver and kidney30. So, the purpose of this study was to investigate the chemical phenotypes, multiple antibiotic resistance (MAR index) and clinical results of A. hydrophila isolated from natural infected O. niloticus and Sparus aurata (fresh and marine water fish), in addition to studying blood chemistry and histological changes in the liver, kidney and spleen of O. niloticus after being infected with A. hydrophila.
MATERIALS AND METHODS
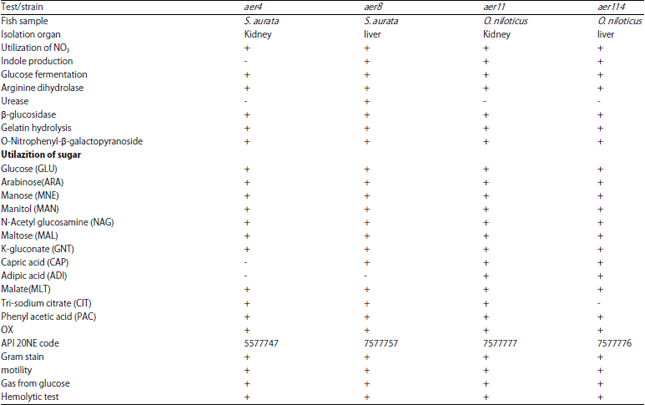
Sampling and bacterial isolation: Aeromonas was isolated from naturally diseased fishes namely O. niloticus, an important fresh fish, farmed in fresh fish farm located on El-Manzala Lake, Egypt and Gilt-head bream (Sparus aurata) cultured in marine fish farm located on the Mediterranean coast in Deeba Triangle in spring season of 2016 during mass mortality. Samples from liver and kidney of infected fish were cultured into trypticase soy broth (TSB; Oxoid) for 24 h at 37±1°C. A loopful of the enrichment culture was streaked on specific medium; Aeromonas Medium Base (AMB, Ryan, Oxoid, Ltd.) that was used for the selective isolation of Aeromonas strains. After 24 h of incubation at 37°C, only the cultures giving pure green with dark centers colonies were randomly selected then subcultured on tryptic soy agar (TSA, Oxoid). Four presumed Aeromonas bacteria, according to the source of isolation, were identified by API 20NE biochemical identification strips (bioMérieux, Marcy l’Etoile, France), (Table 1) after determination the Gram staining for all isolates.
Hemolysin assays: The ability of strains to produce hemolysins was assessed by the plate method. Each isolate was streaked on tryptic soy agar (TSA) medium containing 5% (vol/vol) sheep erythrocytes and incubated at 37°C for 24-48 h. Hemolytic activity was measured as described by Asao et al.31.
DNA isolation and 16S rRNA gene sequencing: Two of the identified isolates were confirmed by PCR. The extraction of bacterial DNA was performed by used QIA gen DNA isolation kit, then amplified with universal bacterial primers DG74 5-AGGAGGTGATCCAACCGCA-3 and RW01 5-AACTGGAGGAAGGTGGGGAT-332. The locations of primers were 1521-1540 and 1170-1189, respectively. After amplification, 10 μL of the PCR sample was electrophoresis. The PCR product was purified with a QIA quick PCR purification kit and directly sequenced with a 3500/3500xL Genetic Analyzer (Applied Biosystems).
| Table 1: | Biochemical patterns of Aeromonas hydrophila isolated from fish |
 | |
| Table 2: | Pathogenicity for O. niloticus of Aeromonas hydrophila isolates by using intraperitoneal injections |
 | |
| +: Isolates positive, -: Isolates negative | |
Multiple antibiotic resistance (MAR index): Multiple antibiotic resistance testing was carried out using the disc diffusion method on Mueller-Hinton Agar (Oxoid, England) and incubated at 37±1°C for 24 h. Eighteen antibiotic disk namely ampicillin (AP) 10 μg, amoxicillin (AX) 25 μg, norfloxacin (NOR) 10 μg, ciprofloxacin (CIP) 10 μg, streptomycin (S) 10 μg, cephazolin (CZ) 30 μg, erythromycin (E) 15 μg, fusidic acid (FA) 10 μg, gentamycin (GN) 120 μg, oxytetracyclin (OTC) 30 μg, nalidixic acid (NA) 30 μg, sulfamethoxazole (SXT) 25 μg, tobramycin (TOB) 10 μg, kanamycin (KA) 30 μg, azithromycin (AZM)15 μg, lomefloxacin (LOM) 10 μg, gatifloxacin (GAT) 5 μg and vancomycin (VA) (Oxoid, England) were used. After incubation, the sizes of the inhibition zones were measured and discussed according to Clinical and Laboratory Standards Institute, CLSI33 (Table 2) and MAR index of the four isolates (aer4, aer8, aer11, aer114) against tested antibiotics was calculated as formula:
Where:
| X | = | Total of antibiotic resistance case |
| Y | = | Total of antibiotic used |
| Z | = | Total of isolates trust and sparrow34 |
Pathogenicity test: Sixty three of apparently healthy O. niloticus (80±2 g, b.wt.) were randomly divided into 3 experimental groups (T1, T2 and T3); each group contained 21 fish kept in three glass aquaria, each aquaria contained 7 fish and were supplied with de-chlorinated water with aeration; the fish were acclimatized for 2 weeks. Fish were fed with commercial diet twice a day at a feeding rate of 3% of B.W. Each fish of T2 and T3 groups was intraperitoneally (IP) injected by 0.5 mL of 1×105 CFU mL–1 of the suspension of either the isolates aer8 or aer114 in groups T2, T3, respectively. Another twenty one fish (T1) were injected with phosphate buffered saline (PBS) using the same procedure. Before injection, fish were anesthetized in a clove oil solution35. The virulence of the strains was classified based on the development of clinical signs and mortality rate within 7 days.
Sample collection and analyses: On day 7 post-infection, blood was collected by cardiac puncture. The serum was separated by centrifugation at 3000 rpm for 15 min and stored at -20°C until further use. The activities of aspartic aminotransferase (AST), alanine aminotransferase (ALT) and the concentrations of serum total protein (TP), albumin (AL) were determined using commercial calorimetric kits (Diamond, Diagnostic, Egypt). The creatinine (Cr) and urea were determined using commercial calorimetric kits (Stanbio Laboratory Diagnostic, USA). All tests were carried out in duplicate.
Histological examination: After collection of the samples from liver, kidney and spleen tissues, samples were fixed in 10% neutral buffered formalin for 24 h and were subjected to histological examination according to Roberts36. Tissue sections were stained with hematoxylin-eosin (HE) and their slides were photographed using an ICC50 HD camera and a Leica LAS EZ microscope.
Analytical methods: The analysis of variance was used to compare the means of different groups using the SAS statistical software package (version 9.3, SAS Institute, Inc., Cary, NC, USA). When one-way ANOVA-test was significant (p<0.05), least significant difference was calculated according to Duncan37 for the comparison among means.
RESULTS
Clinical examination of natural diseased fish: Fish samples were collected from fish farms that suffered from mass mortalities in Damietta governorate Northern of Egypt. Many of O. niloticus samples showed signs of septicemia, while Gilt-head bream fish had no signs. Clinical examination of diseased O. niloticus samples were shown in Fig. 1(a-d), that revealed cloudiness of eyes, detachment and protrusion of the scales, hemorrhage and ulcers on the skin, severe distention of the abdomen and inflammation of the anus.
Biochemical characteristics and API 20NE profile of A. hydrophila : Four of the 16 isolates were grown on AMB medium (Ryan), which were isolated from liver and kidney of both S. aurata (aer4 and aer8) and O. niloticus (aer11 and aer114), respectively. Isolates were characterized on the basis of biochemical characteristics and API 20NE as A. hydrophila, where API 20NE revealed four different profile numbers among the four strains (Table 1). All strains were Gram negative, motile rods and they were all able to produce gas from glucose and they were found to produce β-hemolysin on sheep blood agar media. On the other hand, these isolates differed in their responses to the following tests: Indole, urease and assimilation of capric acid, adipic acid and tri-sodium citrate which gave variable results. Isolates from O. niloticus were assigned aer11 and aer114 and API 20NE profile were 7577777 and 7577776, respectively, while isolates from S. aurata were assigned aer4 and aer8 and API 20NE profile were 5577747 and 7577757, respectively.
Identification by 16S rRNA and sequencing: Two isolates were selected for confirmation by 16S rRNA (aer8 and aer114) and sequenced by 3500 Genetic Analyzer (Applied Biosystems) and were compared to those available in the GenBank database. DG74 and RW01 primers were used to amplify 16S rRNA of bacterial isolates, with amplicon size ∼370 bp. The 2 bacterial species belonged to family Aeromonadaceae. The amplified nucleotide sequence expressed as 98.0% homology with the 16S rDNA sequence of Aeromonas hydrophila. The results of phenotypic classification are correlated well with the sequence of 16S rRNA for Aeromonas strains. The 16S rDNA sequence resulting from this study was deposited in the GenBank under the accession numbers. LC208788, LC208789 of aer8 and aer114, respectively.
Virulence for O. niloticus and clinical observation: The results for the virulence test are summarized in Table 2 and revealed that groups challenged with the two A. hydrophila isolates (aer8 and aer114) caused mortality ranging to 71.4-76.2% (T2 and T3, respectively) within 7 days where 31 out of 42 experimental fish infected (73.8%) died rapidly before they have time to develop great clinical signs but a few clinical abnormalities signs of disease such as extensive skin hemorrhages and inflamed vent were seen. Also internally, congestion and enlargement of liver, kidney and spleen with fatty liver were observed. In contrast, the control fish (T1) observed no clinical signs or death confirmed during the experiment period.
 | |
| Fig. 1(a-d): | Gross pathology for naturally infected O. niloticus (a) Eyes cloudiness and detachment of scales, (b) Skin ulceration, (c) Hemorrhage on the skin and (d) scale protrusion |
| Table 3: | Antibacterial susceptibility and multiple antibiotic resistances of Aeromonas hydrophila isolated from different fish |
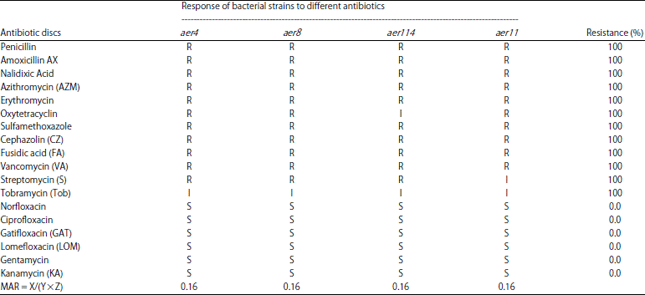 | |
| S: Susceptible, I: Intermediate susceptible, R: Resistant | |
Multiple Antibiotic Resistance (MAR index): All A. hydrophila isolates showed 100% resistance to 12 antibiotic included penicillin, amoxicillin, nalidixic acid, azithromycin, erythromycin, sulfamethoxazole, cephazolin, fusidic acid, vancomycin, streptomycin, oxytetracyclin and tobramycin. On the other hand, all isolates were 100% susceptible to ciprofloxacin, norfloxacin, gatifloxacin, lomefloxacin, gentamycin and kanamycin as shown in Table 3. The four isolates (aer4, aer8, aer11 and aer114) showed multiple resistant patterns to many antibiotics (18) that showed commonly resistant to 12 antibiotics with 66.6% resistance patterns, while MAR index was 0.16.
Assessment of liver and kidney function: Oreochromis niloticus infected by A. hydrophila (T2, T3 groups) showed a significant increase (p<0.05) in activity of serum AST, ALT and in levels of creatinine and urea compared to the uninfected control group (T1).
| Table 4: | Hepatic and kidney parameter functions and total protein, albumin and globulin concentrations in O. niloticus infected by A. hydrophila isolates (Means±SE) |
 | |
| A-C: Means in the same column superscripted with different letters are significantly different at (p<0.05) | |
On the other hand, the results indicates that the total protein, albumin and globulin level significantly decreased (p<0.05) by the infection with A. hydrophila as compared to control (Table 4).
Also, there were significant differences (p<0.05) in ALT between the fish groups infected by isolates aer8 (T2) and aer14 (T3) while the significant differences (p<0.05) were found when comparing their values with the control (T1). In the same trend, group (T3) reflected a significant increase in creatinine if compared with fish infected group (T2). Isolate aer114 was isolated from O. niloticus reflected negative effects on some of blood parameters compared to isolate aer8 isolated from marine water fish.
Histological examination: No histological changes were observed in the liver of control fish (T1) (Plate 1a). The histological changes in the liver of group T2 injected with aer8 showed nuclear and cytoplasmic degeneration, severe dilation in sinusoids with lipid vacuoles (Plate 1b), while aer114 caused pyknotic nuclei, vacuolar degeneration and necrosis (plate 1c) and hemosiderin accumulation, cytoplasmic vacuolation and necrosis in hepatocytes of T3 (Plate 1d).
Kidney of O. niloticus in control group (T1) exhibited normal renal tubules, tubular lumen and Bowman’s space in the glomerular (Plate 1e, f) but kidney of O. niloticus in T2 exhibited tubular degeneration with mononuclear cell infiltration and necrosis area between renal tubules, disconnection of renal tubules and hemosiderin deposits (Plate 2a). Similar lesions were observed in kidney of T3 fish injected with strain aer114, in addition to hemorrhage between renal tubules, glomerular expansion and absence of the Bowman’s space with occlusion of the tubular lumen and accumulation of inflammatory and erythrocytes cells (Plate 2b). On the other hand, the spleen of O. niloticus in control group (T1) showed normal structure red and white pulps (Plates, 2c). The spleen of O. niloticus infected with aer8 (T2) showed severe accumulation of melanomacrophage cells (MMC), necrosis and infiltration of fibroblasts around damage blood vessels (fibrosis) (Plate 2d), while spleen of T3 group injected with aer114 showed accumulations of erythrocytes, hemosiderin and inflammatory cells with necrosis (Plate 2e), in addition to hemolytic between red pulps and vacuolation that were seen (Plate 2f).
DISCUSSION
The clinical signs observed on diseased O. niloticus samples were similar to that described by Aboyadak et al.38, who observed that the diseased fish by A. hydrophila exhibited hemorrhagic patches on the skin and around the anus, scale peeling, skin ulcerations and fin erosions.
Concerning the API 20NE profile numbers of strains aer4, aer8, aer11 and aer114 were 5577747, 7577757, 7577777 and 7577776, respectively, which agree with those given by Paniagua9 where API 20NE profile numbers of A. hydrophila were 7577754, 5177754, 7476754, 7575754, 7576754 and 5577754. In the present study, the different responses of biochemical tests were observed among strains that might be attributed to biological effects, where the microorganisms may be acclimate to very particular environmental conditions by converting expression genes that allows them to survive in specific environments39. El-Barbary22 also, reported that the isolates of A. hydrophila (SSU) isolated from various sources of fish in different places were different in biochemical characteristics according to fish species and water salinity.
The present study used universal primer (RW01 and DG74) that was recorded to be sensitive screening technique for discovery of bacterial groups32,40 and the 423 bp products were successfully amplified in PCR reactions and consequent sequencing confirmed A. hydrophila 16S rRNA fragment.
Studies of the virulence of bacterial pathogens and their sensitivity to antibiotics in fish are important for the development of new chemotherapeutic agents to combat bacterial infections, because the resistance of antibiotic has made bacterial diseases one of the major problems in the fisheries sector41. Pathogenicity experiment of aer8 and aer114 of A. hydrophila cause mortality up to 71.4-76.2% in experimental O. niloticus within 7 days. The pathogenicity of Gram negative bacteria such as A. hydrophila in warm water fishes was reported by Thune14 and the pathogenicity might be contributed to the extracellular toxins and enzymes such as hemolysins, cytotoxins, enterotoxins and proteases have been associated with the virulence of A. hydrophila22,42,43 which causes systemic damage to the hematopoietic system and liver that leads to mortality44. A. hydrophila isolates were found to produce β-hemolysin on sheep blood agar media, these results indicated a very close relationship between hemolysin activity and pathogenicity in A. hydrophila isolates.
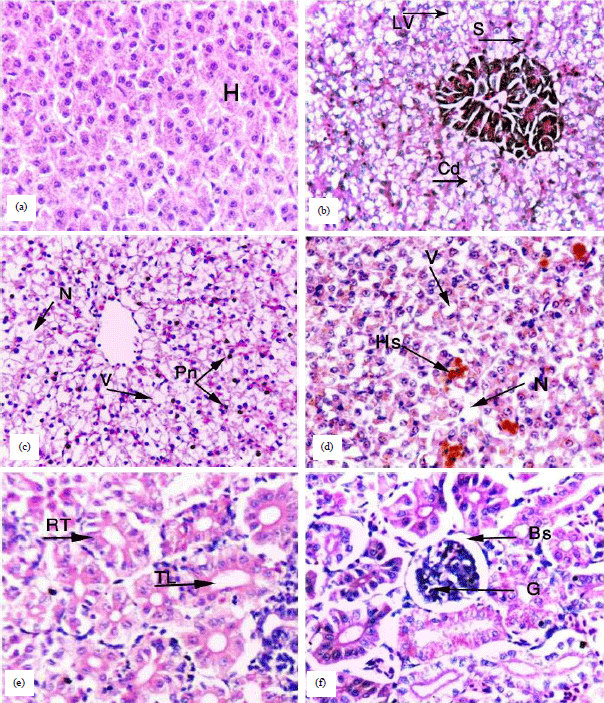 | |
| Plate 1(a-f): | Histopathological changes in the liver (a-d) and kidney (e, f) of O. niloticus groups (T1-T3) stained with H&E; (a,) T1 normal hepatocytes, cytoplasm and central and round nucleus (x300); (b)T2 nuclear and cytoplasmic degeneration, severe dilation in sinusoids with lipid vacuoles (x250); (c,d) T3 pyknotic nuclei, vacuolar degeneration and necrosis (cx200), cytoplasmic vacuolation with necrosis and hemosiderin accumulation (dx300). (e, f) T1 normal renal tubules and tubular lumen (ex 300) and normal glomerular with Bowman’s space (fx 300) |
H: Hepatocyte, Cd: Cytoplasmic degeneration, S: Sinusoids, Lv: Lipid vacuoles, Hs: Hemosiderin, Pn: Pyknotic nucleus, V: Vacuolation, N: Necrosis, G: Glomerular, BS: Bowman’s space, RT: Renal tubular, TL: Tubular lumen | |
Also, Santos et al.45 and El-Barbary22 reported that some biochemical properties as hemolytic activity and production gas from glucose and arabinose can be helpful to differentiate between the pathogenic and non-pathogenic isolates of A. hydrophila. The clinical and postmortem findings post challenged with A. hydrophila were weakness, extensive skin hemorrhages and inflamed vent, internally; congestion and enlargement of liver, kidney and spleen with fatty liver were observed.
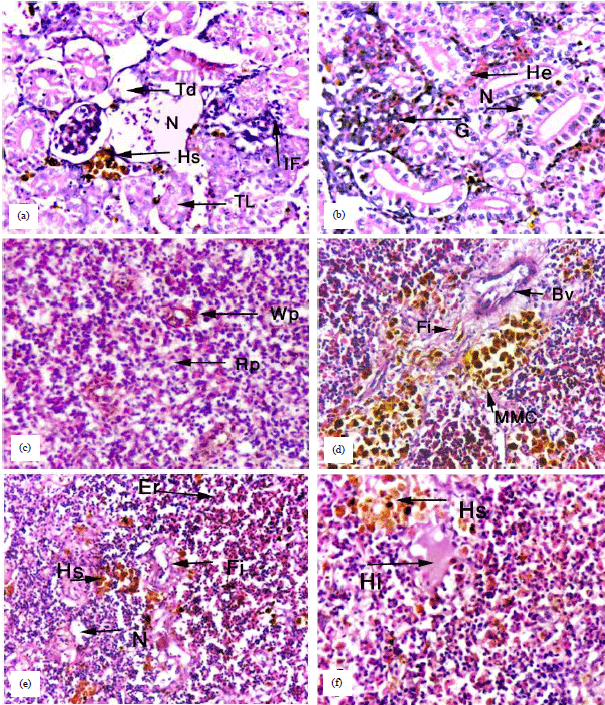 | |
| Plate 2(a-f): | Histopathological changes in the kidney and spleen of O. niloticus groups (T1-T3) stained with H and E.; (a) T2 tubular degeneration, mononuclear cell infiltration, necrosis area between renal tubules, accumulation of hemosiderin and occlusion of the tubular lumen (x300). (b) T3 hemorrhage between renal tubules, glomerular expansion with absence of the Bowman’s space and necrosis (x300). (c) T1 normal structure of red and white pulps in spleen (x200). (d) T2 severe accumulation of MMC and infiltration of fibroblasts around damage blood vessels. (e) T3 erythrocytes and hemosiderin accumulations and necrosis (x200), (f), hemolysis between red pulps and accumulation of hemosiderin (x300) |
| Td: Tubular degeneration, If: Infiltration, N: Necrosis, RT: Renal tubular, Hs: Hemosiderin, TL: Tubular lumen, He: Hemorrhage, G: Glomerular, BS: Bowman’s space, WP: White pulp, RP: Red pulp, MMC: Melanomacrophage cells, Fi: Fibroblasts, Bv: Blood vessel, Er: Erythrocytes, N: Necrosis, Hi: Hemolysis | |
In another study of A. hydrophila, the macroscopic examination showed exophthalmia, skin lesions with hyperaemia of the fins, while necropsy revealed yellow foci on the surface of liver, swollen of gallbladder and fragmentation of kidney and spleen46. In this study aer8 and aer114 isolates were 100% susceptible to six antibiotics and 100% resistance to 12 antibiotics as shown in Table 3. These data may be helpful in controlling and curing bacterial infections caused by A. hydrophila in aquaculture. These results are corresponding to previous data by Emekdas et al.47 and El-Barbary22 who observed that A. hydrophila isolates were found to be resistant to Amoxycillin, Penicillin and Ampicillin. Shak et al.48 also, observed that high antibiotic resistance was seen in bacterial infections caused by Aeromonas sp., thus the emergence of resistance to antibacterial agents in pathogenic bacteria is an international public health problem49.
In addition, the MAR index for the four tested A. hydrophila isolates was 0.16 that is corresponded to the results of Laith and Najiah46 who observed that the MAR index for isolates of A. hydrophila was ranged from 0.10-0.50. The MAR index, that is lower than 0.2 (<0.2) detects strains from the environment, where antibiotics are rarely used or are not used at all 50.However th e MAR index, which is higher than 0.2 (>0.2), detects bacteria isolated from organisms with high infection risk, where antibiotics are often used51.
Results of blood parameters are shown in Table 4 which revealed that A. hydrophila caused a significant increase in activity of serum liver enzymes, AST and ALT, that give indication of liver case, where ALT and AST are considered as biomarkers of hepatic dysfunction and damage30, because the increased AST and ALT values in fish reveal enzymes exporting from liver into bloodstream52,53, which is due to the fact that the fish liver is a major organ involved in metabolic process54. Similar findings were also observed in fish infected by A. hydrophila in studies of Souza et al.24, Dos Santos et al.25 and Ahmed et al.26. On the other hand, A. hydrophila infection caused a significant reduction in serum concentrations of TP, AL and GL compared to the control group (Table 4). This reduction in TP and AL content could be attributed to the damaging effect of A. hydrophila on hepatocytes which were observed in the histological study in T2 and T3 groups. Also, Pal et al.28 found that the total serum protein, albumin and globulin values were significantly (p<0.01) higher in the A. hydrophila infected fishes compared to control fishes. This increase in serum total protein could also be an indication of antibody production in moribund fish with infectious diseases55. In contrast, Dos Santos et al.25 recorded that after A. hydrophila challenge, total protein values were lower in the infected group fish than in the control group, while Li et al.56 reported that total protein, increased at the early infection phase by A. hydrophila and then decreased at other time points in carp. This depression in the total protein of blood is mainly due to excessive loss through nephrosis57. Because the kidney is considered the main excretory organ involved in xenobiotic excretion, its function was investigated in this study, where A. hydrophila led to a significant increase (p<0.05) in serum creatinine and uric acid values (Table 4) in infected groups (T2, T3) as compared with control (T1). Previous studies reported the same findings with creatinine and urea24,25. Also, Wells et al.58 reported that the high creatinine in the blood indicates a low glomerular filtration rate of the fish posterior kidney where creatinine is the product of muscle creatine catabolism and is excreted by trunk kidney59.
Histopathology is widely used to study tissue damage due to various chemicals or biological infectious agent as biological markers worldwide60. In the present study, the histopathological changes of organs in T3 injected with aer114 appeared to be more than those in the fish injected with aer8 (T2).The reason for this result may be due to the similarity in the origin and environmental conditions (O. niloticus and freshwater) between the natural and experimental infection where A. hydrophila was isolated from O. niloticus (aer114) in fresh water and used to infect O. niloticus in aquaria containing fresh water. Liver exhibited severe lesions as nuclear and cytoplasmic degeneration, severe dilation in sinusoids, lipid vacuoles, pyknotic nuclei, hemosiderin accumulation and necrosis in liver hepatocytes. The cause of lipid vacuoles and necrosis in the liver may be due to the presence of some extracellular toxins such as cytotoxins, hemolysin, protease, enterotoxins and elastase produced by A. hydrophila 22,27,61. Similar hepatic lesions were recorded previously in O. niloticus as degenerative changes, cytoplasmic fat vacuoles and lymphocyte infiltration with focal necrosis of hepatocytes and pancreatic cells22,62, also, in other fishes such as catfish (Clarias gariepinus) and carps (Carassius aurata)63,46. While kidney of O. niloticus injected with A. hydrophila (T2, T3) had tubular degeneration, infiltration of mononuclear cell, necrosis area and hemorrhage between renal tubules, accumulation of both melanomacrophage, inflammatory cells and erythrocytes cells, glomerular expansion with absence of the Bowman’s space and occlusion of the tubular lumen which seemed to be likeness to previous studies62 where the kidney of O. niloticus injected with A. hydrophila showed hemorrhage of subcapsular and interstitial, degeneration of parenchyma, formation of cytoplasmic vacuole, necrosis of the tubular epithelium with focal lymphocyte infiltration while interior lumina was filled by yellow mucoid liquid. Also, Laith and Najiah46 recorded histopathological changes in glomerular epithelium in kidney of catfish infected with A. hydrophila.
Spleen is composed of capsule and two general parenchyma components lymphopoietic (white pulp) and the hematopoietic (red pulp), which are important for the defense systems of fishes64-66,where it is the only lymph-node in bony fish67. In the current study, spleen of O. niloticus injected with A. hydrophila exhibited severe accumulation of melanomacrophage cells (MMCs), necrosis and infiltration of fibroblasts around damage blood vessels in addition to hemolytic between red pulps and vacuolation. These lesions may be caused by the presence of different extracellular toxins and enzymes associated with of A. hydrophila and the presence of different toxicants in the environment68,69. The present results are in agreement with the findings of Alsaid et al.70 who observed that red hybrid tilapia injected IP with Streptococcus agalactiae showed red pulp degeneration, spleenic capillary congestion and focal haemorrhages in intercellular spaces together with multiple foci of hemosiderin deposits and melanomacrophage centers in the spleen. Where, bacterial infection increased the number and size of MMCs in the liver and spleen fish71,72. Also, the accumulations of erythrocytes cells in the spleen in turn may result in reduced haemoglobin content which is usually attributed to the destruction of red blood cells and irregular movement of haemoglobin from the spleen in fishes73-75. Deposition of hemosiderin in the liver and spleen of O. niloticus (T2 and T3) infected with A. hydrophila agrees with those of Chang and Plumb76 who showed foci of hemosiderin in Cyprinus carpio spleens exposed to A. hydrophila.
CONCLUSION
It is concluded that A. hydrophila isolates may show slight variation in biochemical characteristics however, the 16S rRNA proved that identified isolates belonged to A. hydrophila. The negative effects of isolate aer114 on organs histology and blood examinations were more than that caused by aer8.
SIGNIFICANCE STATEMENTS
This study confirms that both the API 20NE and sequencing of 16S rRNA can be beneficial for identification of unknown bacterial species of fish pathogen. This study will help the researcher to quickly detect bacterial pathogen in fish farms and treatment of infectious diseases through appropriate antibiotic, leading to reduce the morbidity and mortality rates and contributing to the enhancement of fish production.
REFERENCES
- Beaz-Hidalgo, R. and M.J. Figueras, 2013. Aeromonas spp. whole genomes and virulence factors implicated in fish disease. J. Fish Dis., 36: 371-388.
CrossRefDirect Link - Dallaire-Dufresne, S., K.H. Tanaka, M.V. Trudel, A. Lafaille and S.J. Charette, 2014. Virulence, genomic features and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol., 169: 1-7.
CrossRefDirect Link - Parker, J.L. and J.G. Shaw, 2010. Aeromonas spp. clinical microbiology and disease. J. Infect., 62: 109-118.
CrossRefDirect Link - Monfort, P. and B. Baleux, 1990. Dynamics of Aeromonas hydrophila, Aeromonas sobria and Aeromonas caviae in a sewage treatment pond. Applied Environ. Microbiol., 56: 1999-2006.
Direct Link - Harikrishnan, R., M.N. Rani and C. Balasundaram, 2003. Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture, 222: 41-50.
CrossRefDirect Link - Janda, J.M. and S.L. Abbott, 2010. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev., 23: 35-73.
CrossRefDirect Link - Lewis, W.M. and M. Bender, 1960. Heavy mortality of golden shiners during harvest due to a bacterium of the genus Aeromonas. Progr. Fish-Cult., 22: 11-14.
CrossRefDirect Link - Paniagua, C., O. Rivero, J. Anguita and G. Naharro, 1990. Pathogenicity factors and virulence for rainbow trout (Salmo gairdneri) of motile Aeromonas sp. isolated from a river. J. Clin. Microbiol., 28: 350-355.
Direct Link - Mastan, S.A. and T.A. Qureshi, 2001. Role of bacteria in the Epizootic Ulcerative Syndrome (EUS) of fishes. J. Environ. Biol., 22: 187-192.
PubMedDirect Link - Karunasagar, I., G.M. Rosalind, I. Karunasagar and K.G. Rao, 1989. Aeromonas hydrophila septicaemia of Indian major carps in some commercial fish farms of west Godavari district, Andhra Pradesh. Curr. Sci., 58: 1044-1045.
Direct Link - Thune, R.L., L.A. Stanley and R.K. Cooper, 1993. Pathogenesis of gram-negative bacterial infections in warmwater fish. Annu. Rev. Fish. Dis., 3: 37-68.
CrossRefDirect Link - Nielsen, M.E., L. Hoi, A.S. Schmidt, D. Qian, T. Shimada, J.Y. Shen and J.L. Larsen, 2001. Is Aeromonas hydrophila the dominant motile Aeromonas species that causes disease outbreaks in aquaculture production in the Zhejiang Province of China? Dis. Aquat. Organisms, 46: 23-29.
CrossRefPubMedDirect Link - Fang, H.M., R. Ge and Y.M. Sin, 2004. Cloning, characterisation and expression of Aeromonas hydrophila major adhesin. Fish Shellfish Immunol., 16: 645-658.
CrossRefDirect Link - Larsen, J.L. and N.J. Jensen, 1977. An Aeromonas species implicated in ulcer-disease of the cod (Gadus morhua). Nord. Vet. Med., 29: 199-211.
PubMedDirect Link - Xia, C., Z.H. Ma, M.H. Rahman and Z.G. Wu, 2004. PCR cloning and identification of the β-haemolysin gene of Aeromonas hydrophila from freshwater fishes in China. J. Aquacult., 229: 45-53.
CrossRefDirect Link - Low, K.W. and Y.M. Sin, 1998. Effects of mercuric chloride and sodium selenite on some immune responses of blue gourami, Trichogaster trichopterus (pallus). Sci. Total Environ., 214: 153-164.
CrossRefDirect Link - Sarmento, A., L. Guilhermino and A. Afonso, 2004. Mercury chloride effects on the function and cellular integrity of sea bass (Dicentrarchus labrax) head kidney macrophages. Fish Shellfish Immunol., 17: 489-498.
CrossRefDirect Link - Iregui, C.A., J. Comas, G.M. Vasquez and N. Verjan, 2016. Experimental early pathogenesis of Streptococcus agalactiae infection in red tilapia Oreochromis spp. J. Fish Dis., 39: 205-215.
CrossRefDirect Link - El-Barbary, M.I., 2010. Some clinical, microbiological and molecular characteristics of Aeromonas hydrophila isolated from various naturally infected fishes. Aquacult. Int., 18: 943-954.
CrossRefDirect Link - Abd-El-Rhman, A.M.M., 2009. Antagonism of Aeromonas hydrophila by propolis and its effect on the performance of Nile Tilapia, Oreochromis niloticus. Fish Shellfish Immunol., 27: 454-459.
CrossRefDirect Link - Souza, C.F., M.D. Baldissera, R.A. Vaucher, L.Q.S. Lopes and B.S. Vizzotto et al., 2016. In vivo bactericidal effect of Melaleuca alternifolia essential oil against Aeromonas hydrophila: Silver catfish (Rhamdia quelen) as an experimental model. Microb. Pathog., 98: 82-87.
CrossRefDirect Link - Dos Santos, A.C., F.J. Sutili, B.M. Heinzmann, M.A. Cunha, I.C.M. Brusque, B. Baldisserotto and C.C. Zeppenfeld, 2017. Aloysia triphylla essential oil as additive in silver catfish diet: Blood response and resistance against Aeromonas hydrophila infection. Fish Shellfish Immunol., 62: 213-216.
CrossRefDirect Link - Ahmad, M.H., A.M.D. El Mesallamy, F. Samir and F. Zahran, 2011. Effect of cinnamon (Cinnamomum zeylanicum) on growth performance, feed utilization, whole-body composition and resistance to Aeromonas hydrophila in Nile tilapia. J. Applied Aquacult., 23: 289-298.
CrossRefDirect Link - Allan, B.J. and R.M.W. Stevenson, 1981. Extracellular virulence factors of Aeromonas hydrophila in fish infections. Can. J. Microbiol., 27: 1114-1122.
CrossRefDirect Link - Maqsood, S., M.H. Samoon and P. Singh, 2009. Immunomodulatory and growth promoting effect of dietary levamisole in Cyprinus carpio fingerlings against the challenge of Aeromonas hydrophila. Turk. J. Fish. Aquat. Sci., 9: 111-120.
Direct Link - Recknagel, R.O., E.A. Glende, Jr., J.A. Dolak and R.L. Waller, 1989. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther., 43: 139-154.
CrossRefPubMedDirect Link - Asao, T., S. Kozaki, K. Kato, Y. Kinoshita, K. Otsu, T. Uemura and G. Sakaguchi, 1986. Purification and characterization of an Aeromonas hydrophila hemolysin. J. Clin. Microbiol., 24: 228-232.
Direct Link - Greisen, K., M. Loeffelholz, A. Purohit and D. Leong, 1994. PCR primers and probes for the 16S rRNA gene of most species of pathogenic bacteria, including bacteria found in cerebrospinal fluid. J. Clin. Microbiol., 32: 335-351.
Direct Link - Trust, T.J. and R.A. Sparrow, 1974. The bacterial flora in the alimentary tract of freshwater salmonid fishes. Can. J. Microbiol., 20: 1219-1228.
CrossRefPubMedDirect Link - Hamackova, J., J. Kouril, P. Kozak and Z. Stupka, 2006. Clove oil as an anaesthetic for different freshwater fish species. Bulg. J. Agric. Sci., 12: 185-194.
Direct Link - Aboyadak, I.M., N.G.M. Ali, A.M.A.S. Goda, W.H. Aboelgalagel and A.M.E. Alnokrashy, 2015. Molecular detection of Aeromonas hydrophila as the main cause of outbreak in tilapia farms in Egypt. J. Aquacult. Mar, Biol., Vol. 2, No. 5.
CrossRefDirect Link - Szczuka, E. and A. Kaznowski, 2007. Characterization of Aeromonas caviae and A. veronii by standardized cellular protein electrophoretic patterns. Folia Microbiol., 52: 65-69.
PubMedDirect Link - Teng, L.J., P.R. Hsueh, Y.H. Huang and J.C. Tsai, 2004. Identification of Bacteroides thetaiotaomicron on the basis of an unexpected specific amplicon of universal 16S ribosomal DNA PCR. J. Clin. Microbiol., 42: 1727-1730.
CrossRefDirect Link - Shayo, S.D., C.J. Mwita and K.M. Hosea, 2012. Virulence of Pseudomonas and Aeromonas bacteria recovered from Oreochromis niloticus (Perege) from Mtera hydropower Dam; Tanzania. Ann. Biol. Res., 3: 5157-5161.
Direct Link - Burke, V., J. Robinson, H.M. Atkinson and M. Gracey, 1982. Biochemical characteristics of enterotoxigenic Aeromonas spp. J. Clin. Microbiol., 15: 48-52.
Direct Link - Stelma, Jr.G.N., C.H. Johnson and P. Spaulding, 1986. Evidence for the direct involvement of β-hemolysin in Aeromonas hydrophila enteropathogenicity. Curr. Microbiol., 14: 71-77.
CrossRefDirect Link - Brenden, R.A. and H.W. Huizinga, 1986. Pathophysiology of experimental Aeromonas hydrophila infection in goldfish, Carassius auratus (L.). J. Fish Dis., 9: 163-167.
CrossRefDirect Link - Santos, Y., A.E. Toranzo, J.L. Barja, T.P. Nieto and T.G. Villa, 1988. Virulence properties and enterotoxin production of Aeromonas strains isolated from fish. Infect. Immun., 56: 3285-3293.
Direct Link - Laith, A.R. and M. Najiah, 2013. Aeromonas hydrophila: Antimicrobial susceptibility and histopathology of isolates from diseased catfish, Clarias gariepinus (Burchell). J. Aquacult. Res. Dev., Vol. 5.
CrossRef - Emekdas, G., G. Aslan, S. Tezcan, M.S. Serin, C. Yildiz, H. Ozturhan and R. Durmaz, 2006. Detection of the frequency, antimicrobial susceptibility and genotypic discrimination of Aeromonas strains isolated from municipally treated tap water samples by cultivation and AP-PCR. Int. J. Food Microbiol., 107: 310-314.
CrossRefDirect Link - Shak, J.R., J.A. Whitaker, B.S. Ribner and E.M. Burd, 2011. Aminoglycoside-resistant Aeromonas hydrophila as part of a polymicrobial infection following a traumatic fall into freshwater. J. Clin. Microbiol., 49: 1169-1170.
CrossRefDirect Link - Chugh, T.D., 2008. Emerging and re-emerging bacterial diseases in India. J. Biosci., 33: 549-555.
CrossRefDirect Link - Krumperman, P.H., 1983. Multiple antibiotic resistance indexing of Escherichia coli to indentify high-risk sources of fecal contamination of foods. Applied Environ. Microbiol., 46: 165-170.
PubMedDirect Link - Orozova, P., V. Chikova and H. Najdenski, 2010. Antibiotic resistance of pathogenic for fish isolates of Aeromonas spp. Bulg. J. Agric. Sci., 16: 376-386.
Direct Link - Yang, J.L. and H.C. Chen, 2003. Serum metabolic enzyme activities and hepatocyte ultrastructure of common carp after gallium exposure. Zool. Stud., 42: 455-461.
Direct Link - Perez-Rostro, C.I., I.S. Racotta and A.M. Ibarra, 2004. Decreased genetic variation in metabolic variables of Litopenaeus vannamei shrimp after exposure to acute hypoxia. J. Exp. Mar. Biol. Ecol., 302: 189-200.
CrossRefDirect Link - Pacheco, M. and M.A. Santos, 2001. Tissue distribution and temperature-dependence of Anguilla Anguilla L. EROD activity following exposure to model inducers and relationship with plasma cortisol, lactate and glucose levels. Environ. Int., 26: 149-155.
CrossRefDirect Link - Rehulka, J. and B. Minarik, 2007. Blood parameters in brook trout Salvelinus fontinalis (Mitchill, 1815), affected by columnaris disease. Aquacult. Res., 38: 1182-1197.
CrossRefDirect Link - Li, L., Y. Dang, Y. Shen, X. Xu, W. Huang and J. Li, 2016. Hematological and immunological plasma assays for grass carp (Ctenopharyngodon idella) infected with Aeromonas hydrophila as an immune model in carp aquaculture. Fish Shellfish Immunol., 55: 647-653.
CrossRefDirect Link - Yousef, M.I., F.M. El-Demerdash and F.M.E. Radwan, 2008. Sodium arsenite induced biochemical perturbations in rats: Ameliorating effect of curcumin. Food Chem. Toxicol., 46: 3506-3511.
CrossRefDirect Link - Wells, R.M., R.H. McIntyre, A.K. Morgan and P.S. Davie, 1986. Physiological stress responses in big gamefish after capture: observations on plasma chemistry and blood factors. Comp. Biochem. Physiol. A Comp. Physiol., 84: 565-571.
PubMedDirect Link - Trumble, S.J., M.A. Castellini, T.L. Mau and J.M. Castellini, 2006. Dietary and seasonal influences on blood chemistry and hematology in captive harbor seals. Mar. Mammal Sci., 22: 104-123.
CrossRefDirect Link - Camargo, M.M.P. and C.B.R. Martinez, 2007. Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream. Neotrop. Ichthyol., 5: 327-336.
CrossRefDirect Link - Rodriguez, L.A., A.E. Ellis and T.P. Nieto, 1993. Effects of the acetylcholinesterase toxin of Aeromonas hydrophila on the central nervous system of fish. Microb. Pathog., 14: 411-415.
CrossRefPubMedDirect Link - Yardimci, B. and Y. Aydin, 2011. Pathological findings of experimental Aeromonas hydrophila infection in Nile tilapia (Oreochromis niloticus). Ankara Univ. Vet. Fak. Derg., 58: 47-54.
Direct Link - Miyazaki, T. and N. Kaige, 1985. A histopathological study on motile aeromonad disease of Crucian carp. Fish Pathol., 21: 181-185.
CrossRefDirect Link - Fournier-Betz, V., C. Quentel, F. Lamour and A. Leven, 2000. Immunocytochemical detection of Ig-positive cells in blood, lymphoid organs and the gut associated lymphoid tissue of the turbot (Scophthalmus maximus). Fish Shellfish Immunol., 10: 187-202.
CrossRefDirect Link - Lin, H.T., H.Y. Lin and H.L. Yang, 2005. Histology and histochemical enzyme-staining patterns of major immune organs in Epinephelus malabaricus. J. Fish Biol., 66: 729-740.
CrossRefDirect Link - Kurtovic, B., E. Teskerezi and Z. Teskerezi, 2008. Histological comparison of spleen and kidney tissue from farmed and wild European sea bass (Dicentrarchus labrax L.). Acta Adriat., 49: 147-154.
Direct Link - Gogal, Jr.R.M., B.J. Smith, J.L. Robertson, S.A. Smith and S.D. Holladay, 1999. Tilapia (Oreochromis niloticus) dosed with azathioprine display immune effects similar to those seen in mammals, including apoptosis. Vet. Immunol. Immunopathol., 68: 209-227.
CrossRefDirect Link - Garcia-Abiado, M.A., G. Mbahinzireki, J. Rinchard, K.J. Lee and K. Dabrowski, 2004. Effect of diets containing gossypol on blood parameters and spleen structure in tilapia, Oreochromis sp., reared in a recirculating system. J. Fish Dis., 27: 359-368.
CrossRefPubMedDirect Link - Alsaid, M., H. Mohd, N. Mohamed, S. Khairani and Y. Mohamed et al., 2013. Pathological findings of experimental Streptococcusa galactiae infection in red hybrid tilapia (Oreochromis sp.). Proceedings of the International Conference on Chemical, Agricultural and Medical Sciences, December 29-30, 2013, Kuala Lumpur, Malasia, pp: 70-73.
- Alagappan, K.M., B. Deivasigamani, S. Kumaran and M. Sakthivel, 2009. Histopathological alterations in estuarine catfish (Arius maculatus; Thunberg, 1792) due to Aeronomas hydrophila infection. World J. Fish Mar. Sci., 1: 185-189.
Direct Link - Soto, E., J.P. Hawke, D. Fernandez and J.A. Morales, 2009. Francisella sp., an emerging pathogen of tilapia, Oreochromis niloticus (L.), in Costa Rica. J. Fish Dis., 32: 713-722.
CrossRefPubMedDirect Link - Scott, A.L. and W.A. Rogers, 1981. Haematological effects of prolonged sublethal hypoxia on channel catfish* Ictalurus punctatus (Rafinesque). J. Fish Biol., 18: 591-601.
CrossRefDirect Link - Chang, P.H. and J.A. Plumb, 1996. Histopathology of experimental Streptococcus sp. infection in tilapia, Oroochromis niloticus (L.) and channel catfish, Ictafurus punctatus (Ratinesque). J. Fish Dis., 19: 235-241.
CrossRefDirect Link