
Keukeu K. Rosada
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Navisan Najia
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Rahma Widya Ningrum
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Dea Indriani Astuti
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Gede Suantika
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Pingkan Aditiawati
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 1 | Page No.: 11-20







ABSTRACT
Background and Objective: Biofilm plays an important role in causing microbial corrosion. One of prerequisites for microbes as energy source to grow and to form biofilm is carbon source. The purposes of this study were measuring the ability of biofilm community sampled from Saguling hydro power in utilizing carbon sources and assessing culturable heterotrophic of bacterial community from those biofilm. Materials and Methods: Biolog EcoPlateTM and culture-dependent approach were used to assess biofilm community. Results: Heterotrophic bacteria in biofilm have the ability to use 30 different carbon sources consistently. The source of carbon used by this community at the highest rate are N-acetyl-D-glucosamine from carbohydrates group, 2-hydroxybenzoic acid from carboxylic and kenotic acids, glycogen from polymers group and L-serine from the amino acid group at the rate of 0.16, 0.05, 0.14 and 0.09 absorbance U h–1 respectively. Whereas the carbon sources with high consumption are as follow: Pyruvic acid methyl ester, β-methyl-D-glucoside, D-mannitol and N-acetyl-D-glucosamine from carbohydrates group; D-galacturonic acid, 2-hydroxybenzoic acid, D-glucosaminic acid and D-malice acid from the group of carboxylic and kenotic acids; tween 40 and 80 as polymers; L-argentine, L-asparagines, L-serine, L-heroine and glycol-L-glutei from amino acids group. Furthermore, culturable bacterial community of those biofilm were dominated by Gram-negative bacteria, consisted of five common/heterotrophic bacteria, two manganese bacteria, two nitrifying bacteria, three iron bacteria and three sulfate reducing bacteria. Conclusion: Bacterial community of biofilm from Saguling hydro power which were dominated by Gram-negative bacteria have the ability to use various carbon sources and degrade glycogen.
PDF Abstract XML References Citation
Received: October 08, 2016;
Accepted: November 10, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Keukeu K. Rosada, Navisan Najia, Rahma Widya Ningrum, Dea Indriani Astuti, Gede Suantika and Pingkan Aditiawati, 2017. The Ability of Biofilm Community Sampled from Metal Surfaces at Saguling Hydro Power in Utilizing Carbon Sources by Using Biolog EcoPlateTM. Journal of Biological Sciences, 17: 11-20.
DOI: 10.3923/jbs.2017.11.20
URL: https://scialert.net/abstract/?doi=jbs.2017.11.20
DOI: 10.3923/jbs.2017.11.20
URL: https://scialert.net/abstract/?doi=jbs.2017.11.20
INTRODUCTION
Microbiologically Influenced Corrosion (MIC) refers to the influence of microorganisms on the kinetics of corrosion processes of metals, caused by microorganisms adhering to the interfaces (biofilm)1. Those processes involving microorganisms or the products of its metabolic activity including enzymes, exopolimer as well as organic acids and volatile components in the form of inorganic such as ammonia and hydrogen sulfide2. Javaherdashti3 noted that microorganisms are capable of affecting the extent and severity of corrosion. Thus, Coetser and Cloete4 emphasized that the economic implications of those microbial fouling in industrial water systems are much greater than many people realize.
Biofilm is a complex assemblages of microorganisms that are embedded in a matrix of extracellular polymeric substances5. According to Horemans et al.6, environmental Dissolved Organic Matter (DOM) as the main carbon source governs biofilm formation. Most exopolymer-producing microorganisms utilize carbohydrates as their carbon and energy source and amino acids as their source of nitrogen in their growth and extracellular biopolymer’s synthesis7. On the cells of Aureobasidium pullulans, carbohydrates, such as glucose and mannose, determine the exopolymer production8. Santos et al.9 noted that production of bacterial cellulose as an insoluble exopolysaccharide generated by bacterial species lies in its ability to synthesize glucose from the carbon substrate, followed by its polymerization to cellulose. In addition, the differences in biofilm formation may relate to differential expression of fimbriae on the cell surface10. According to Edwards and Schifferli11, regulation of fimbrial production is also affected by carbon source and nitrogen source.
In nature, bacterial induced corrosion do not work solely but rather in a consorsium. Differences in bacterial species composition of biofilm consortia and resulting differences in metabolic activities within such biofilm could explain why, in two identical systems under the same environmental condition, corrosion rates can vary significantly12. Therefore, in order to investigate microbiologically-influenced corrosion failures of ferrous metals in Saguling hydro power, besides molecular approach using DGGE technique13, further studies considering physiological activity of biofilm community and culturable bacterial community residing in this biofilm are important. The main objectives of this study were to measure the ability of biofilm community in utilizing carbon sources by using Biolog EcoPlateTM and to assess culturable heterotrophic bacterial community from biofilm using culture-dependent approach.
MATERIALS AND METHODS
Site and sample collection: Saguling hydro power is located in West Bandung Regency, West Java province, Indonesia (South 06°51'49.8", East 107°20'57", the altitude 643 m). Biofilms samples were taken from the power house of saguling hydro power. Those biofilm were gently scrapped from steel surfaces and composited in a sterile tube before stored in an ice-chest.
Measurement of community substrate utilization profiles: Carbon source utilization by heterotrophic aerobic bacterial community of biofilm were assessed by Biolog EcoPlateTM which contains 31 of the most useful carbon source for soil community analysis. The sample was serially diluted with 0.85% sterile saline solution up to a 10–3 w/v dilution (approximately 1×106 CFU mL–1). Each well of the EcoPlate was inoculated by 100 μL of the sample suspension and incubated at 30°C in a dark chamber. The iMarkTM microplate reader (BioRad) was used to read the absorbance in each well at 590 nm (OD590) at even intervals from 0-168 h.
The utilization of any carbon sources by the bacterial community resulted in the respiration-dependent reduction of the tetrazolium dye and purple color formation measured as absorbance, that can be quantified and monitored over time as average well-color development (AWCD) index. The AWCD is bacterial responses in each microplate that express overall activity of bacterial community and calculated as the arithmetic average of the sum of positive carbon responses corrected by subtracting the water control wells values with the equation:
where, ODi is optical density value from each well14. The percentage of functional diversity is determined by the equation:
This value varies from 0-100% with 0 being low diversity and 100% being highly diverse. Furthermore, the inconsistency found within each sample is calculated by percentage of variation of results within sample in the formula (100×i)/31, where, i is the number of carbon sources in which the three replicates were not all positive or all negative.
Beside AWCD, shannon diversity index (H) and evenness index (E) were investigated using formulae: H = -Σpi(lnpi) and E = H/lnS respectively, where, pi is the proportional color development of the well over total color development of all wells of a plate and S is the number of wells with color development (substrate utilization richness)14. The PCA analysis was used to evaluate the substrates which were the most utilized in biofilm using Minitab 17.
Isolation and enumeration of bacteria: In this study, culture-dependent approach was used to isolate, grow and enumerate culturable bacteria associated with microbial corrosion from the biofilm on various agar media. The nutrient agar, Fe bacterial isolaton medium, Mn base agar and winogradsky medium were used to enumerate the heterotrophic bacteria, iron bacteria, manganese-oxidizing bacteria and acid producers bacteria, respectively15. To obtain Sulfate Reducing Bacteria (SRB), the sulfate reducing API agar (Sigma-Aldrich) recommended by American Petroleum Institute was used. Bacterial enumeration was done by total plate count method16.
Biochemical characterization of isolates: All bacteria isolated from all types of media were identified according to Bergey’s manual of determinative bacteriology17. The pure isolated bacterial cultures were identified by their morphological and biochemical characterization using the following analysis: Gram staining, motility test, indole production test, methyl red test, Voges-Proskauer test, citrate utilization test, nitrate reduction test, H2S production test, urease test, carbohydrate fermentation test, starch, lipid, casein and gelatin hydrolysis test16.
RESULTS
The ability of heterotrophic bacteria from the biofilm samples for both culturable and unculturable in metabolizing a variety of carbon sources were analyzed using Biolog EcoPlateTM (Table 1). In general, all of the carbon sources can be used by the bacteria present on biofilm except for phenyl ethylamine which is included in the group of amines/amides. Shannon diversity index (H) on the use of the substrates showed a value of 3.24±0.08 with the evenness index (E) as much as 0.965±0.018.
| Table 1: | Biochemical diversity: Community Level Physiological Profiles (CLPP) using Biolog EcoPlateTM |
 | |
| *Gryta et al.14, **Positives from the use of the substrate in three replication | |
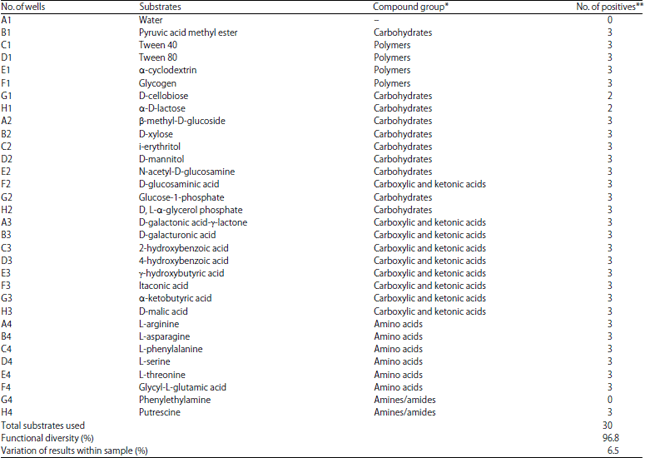
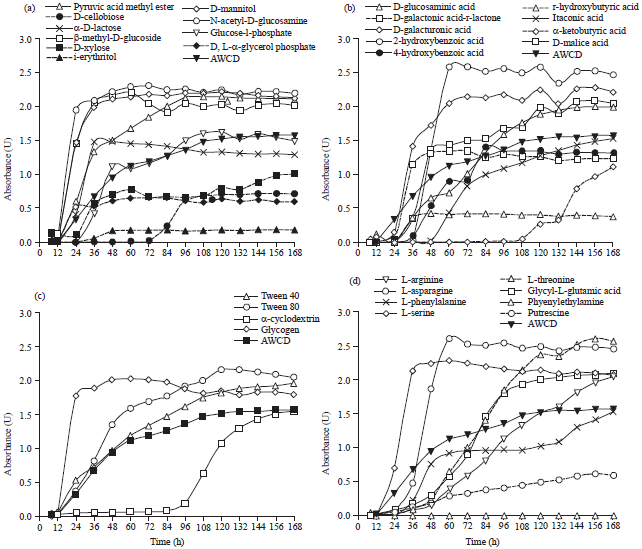
Furthermore, the pattern of reaction of bacteria in biofilm communities over time were analyzed in the form of Average Well Color Development (AWCD) curve until it reached a stationary phase that was in 168 h (7th day) and the use of each carbon source in different groups that were compared with an average of the use of overall carbon sources by the bacterial community (AWCD) during 168 h (Fig. 1). The differences of the consumption of each carbon source up to 168 h analyzed by PCA based on the absorbance values can be seen in Fig. 2.
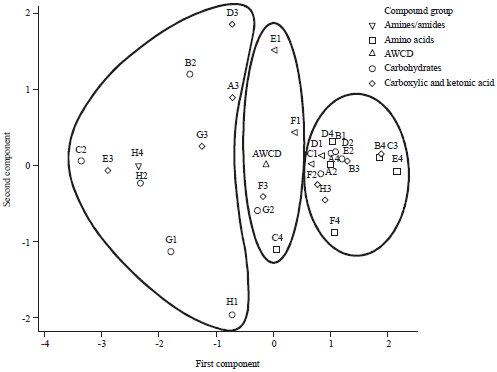
Figure 3 shows the data on the number of bacteria from different groups that were isolated from biofilm. Aerobic bacterial group consist of heterotrophic/common bacteria, manganese oxidizing bacteria, nitrifying bacteria as well as iron bacteria, while anaerobic bacterial group consist of sulfate-reducing bacteria. It can be seen that bacterial plate count for aerobic bacteria obtained from various cultivation media were in the range of 3.4×103 to 3.9×105 CFU g–1 whereas, for anaerobic bacteria were in the number of 1.1×103 CFU g–1.
There were 12 aerobic and 3 anaerobic dominant isolates that found in the culture observed. The aerobic isolates consist of common/heterotrophic bacteria (Brevibacillus laterosporus (NA1), Bacillus lentus (NA2), Burkholderia caryophylli (NA3), Xenorhabdus japonica (NA4) and Bacillus acidiproducens (NA5)), manganese bacteria (Spirosoma sp. (MN1)) and Runella sp. (MN2)), nitrifying bacteria (Nitrosococcus sp. (WN1) and Nitrosomonas europaea (WN2)) and iron bacteria (Burkholderia pseudomallei (FB1), Yersinia pseudo tuberculosis (FB2) and Enterobacter cancerogenus (FB3)), whereas, anaerobic isolates were Desulfobacterium sp. (API1), Desulfobulbus elongatus (API2) and Desulfobacter sp. (API3). The biochemical characteristics of those isolates are shown in Table 2.
 | |
| Fig. 1(a-d): | Rate of the use of each carbon source in different groups that were compared with an average of the use of overall carbon sources by the bacterial community in biofilm (AWCD) during 168 h (n = 3), (a) Carbohydrates group, (b) Carboxylic and ketonic acids group, (c) Polymers group and (d) Amino acids and amines/amides group |
 | |
| Fig. 2: | PCA plot of the two first principal components for the use of carbon sources during 168 h based on the absorbance values from the EcoPlates (in three replicates). There are three groups of substrates with different level of utilization: Low, average and high |
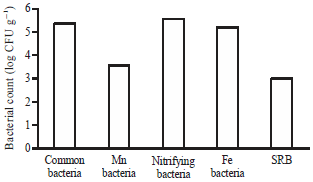 | |
| Fig. 3: | Estimation of bacterial plate count from biofilm at various cultivation media |
DISCUSSION
Functional diversity of bacterial biofilm in substrate utilization reached 96.8% with low variation of results in the sample (6.5%). This indicated that the bacterial community can use almost all of the carbon sources consistently. The calculation of the shannon diversity and evenness index of the use of the substrates showed that the community of biofilm have the ability evenly in the use of diverse carbon sources. Furthermore, in general the biofilm community showed the high rate of the use of the carbon sources, mainly from the carbohydrate group at the 24 h.
Carbon source from the group of carbohydrates that was used with the highest level of respiration was N-acetyl-D-glucosamine (GlcNAc) at the rate of 0.16 absorbance U h–1. GlcNAc is transported into the cell via the phosphotransferase system to form GlcNAc-6-phosphate (GlcNAc-6-P) which is then converted into GlcN-6-P by the enzyme nagA deacetylase. Herein after, GlcN-6-P enter either the glycolytic pathway through fructose-6-P by nagB deaminase or murein and lipopolysaccharide biosynthesis pathway via UDP-GlcNAc by GlmM and GlmU18.
Carbon source from the group of carboxylic and ketonic acids that was used with the highest level of respiration was 2-hydroxybenzoic acid at a rate of 0.05 absorbance U h–1. The 2-hydroxybenzoic acid degraded either through the catechol or salicylate pathway by enzyme gentisate 1-hydroxylase and salicylate 5-hydroxylase19. Furthermore, the carbon sources of the polymer group used with the highest level of respiration was glycogen at the rate of 0.14 absorbance U h–1. Glycogen is a polymer of glucose. Glycogen is catabolized through phosphorylation reaction by glycogen phosphorylase (GlgP) and GlgX into glucose-1-phosphate20 which can later be converted into glucose-6-phosphate as the initial substrate of glycolysis.
| Table 2: | Biochemical characterization of isolates from biofilm |
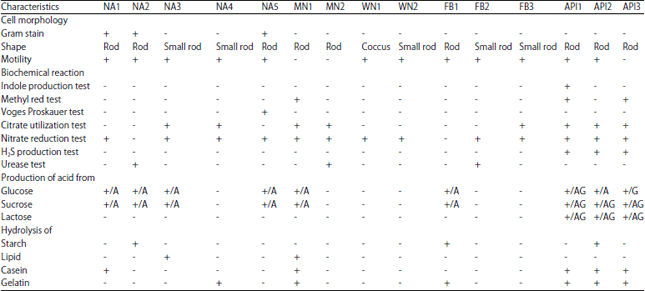 | |
A: Acid, G: Gas, Brevibacillus laterosporus (NA1), Bacillus lentus (NA2), Burkholderia caryophylli (NA3), Xenorhabdus japonica (NA4), Bacillus acidiproducens (NA5), Spirosoma sp. (MN1), Runella sp. (MN2), Nitrosococcus sp. (WN1), Nitrosomonas europea (WN2), Burkholderia pseudomallei (FB1), Yersinia pseudotuberculosis (FB2), Enterobacter cancerogenus (FB3), Desulfobacterium sp. (API1), Desulfobulbus elongatus (API2), Desulfobacter sp. (API3) | |
Meanwhile, the carbon source of the amino acid groups that were used with the highest level of respiration is L-serine with the rate of 0.09 absorbance U h–1. L-serine plays a role in protein synthesis and a compound involved in many metabolic pathways21, one of which is serine-deamination pathway which converts L-serine to pyruvate by the enzyme of L-serine deaminase (sdaA)22. Besides carbon sources with high rate of utilization, there were several sources of carbon with the low rate of usage i.e., D-cellobiose and i-erythritol from the carbohydrate group, α-ketobutyric acid and γ-hydroxybutyric acid of the group of carboxylic and ketonic acids, α-cylodextrin from the polymer group and putrescine of amines/amides groups.
According to PCA plot, there were three groups of the use of the substrates, one of which was the group with high level of consumption of the carbon sources, namely pyruvic acid methyl ester, β-methyl-D-glucoside, D-mannitol and N-acetyl-D-glucosamine from carbohydrates group; D-galacturonic acid, 2-hydroxybenzoic acid, D-glucosaminic acid and D-malic acid from the group of carboxylic and ketonic acids; tween 40 and 80 as polymers; L-arginine, L-asparagine, L-serine, L-threonine and glycyl-L-glutamic from amino acids group. Ultimately, the information regarding the type and the rate of use of these carbon sources can be used as a marker to identify metabolic pathways traversed by bacteria and enzymes that are involved in it. Horemans et al.6 stated that biofilm biomass, species composition, architecture and colocalization of member strains depended on carbon source and its biodegradability.
Conventional bacteriological methods were used to isolate bacteria with corrosive potential, from a wide diversity of environments including biofilm. Since biofilm consist of various microorganisms, therefore in the present study various cultivation media were used. Among the bacteria isolated, Gram-negative bacteria were more dominant than Gram-positive bacteria by 80%. It indicated that Gram-negative bacteria were more active in the formation of biofilm in freshwater. This result is consistent with the observations by Rickard et al.23 that a Gram-negative bacterium was isolated from a freshwater biofilm developed on a stainless steel surface and that Gram-negative organisms dominated in all of the biofilms observed. Biopolymers and lipopolysaccharides constituting Gram negative bacterial cell walls play an important role in cell adhesion to the hydrophilic material24.
Furthermore, almost all of the isolates were rod shaped whereas one isolates was coccus and among 15 isolates, 12 isolates have the ability to move from one place to another. The ability to move was one of the properties of the cell due to the presence of fimbriae and flagella that influence the rate and extent of attachment of microbial cell to a surface25. Coetser and Cloete4 noted that when bacteria attach to a surface, a whole different suite of genes is activated, making sessile bacteria significantly different to planktonic bacteria suspended in the water.
Generally, according to the biochemical analysis, all of the isolates obtained showed different physiological characteristics with various responses to the tests including intracellular enzymatic activities. These biochemical fingerprints indicate the properties of microorganisms that are controlled by the cell’s enzymatic activities and responsible for bioenergetics, biosynthesis and biodegradation16. This heterogenous type of bacteria and growth activity in biofilm will led to the formation of colonization. The colonization of bacteria that is not uniform resulting in the formation of aeration cell differentiation26. Thereby, resources utilization becomes optimized and the processes that require sintrophic relationships or special micro-environment become facilitated27.
In this study, bacterial isolates with corrosive potential have been found. Heterotrophic bacteria Brevibacillus laterosporus has been proven to cause corrosion28, however its mecanism in causing microbial corrosion has not been known yet. Due to aerobic, this bacteria thought to play a role as a constituent of aerobic biofilm that consume oxygen as to allow anaerobic bacteria such as sulfate-reducing bacteria to grow in the inner lining of biofilm29. In addition, the similar interaction also has been seen on the role of Bacillus acidiproducens in causing microbial corrosion indirectly. Beside create an anaerobic environment, this isolate can produce lactic acid30 which is utilized by sulfate-reducing bacteria as organic carbon source for its growth31. On the other hand, lactic acid itself could potentially cause biocorrosion32.
Nitrifying bacteria Nitrosococcus halophilus and Nitrosomonas europea have been isolated with the highest number among other groups of bacteria. As a chemolitothroph autotroph obligate, the abundance of those bacteria were supported by the content of inorganic carbon compound in the form of CO2 in the water33. The abundance of nitrifying bacteria has been known to contribute on the microbial corrosion by increasing the concentration of nitric acid which is corrosive34, produce nitrate that can increase corrosion rate35 and can be associated with sulfate-reducing bacteria36. The previous study has been revealed that amonium, nitrate, nitrite and chloride ion produced in the biofilm could cause localized corrosion37.
Spirosoma sp., has not been known yet as bacterial that influence corrosion. However, these genus often classified as iron bacteria38. Probably, these bacteria play a role in dissolving the iron oxide or passive layer, thereafter increase the corrosion rate39. Generally, iron/manganese oxidizing bacteria acquire the energy from oxidizing Fe2+ to Fe3+ such as Gallionella sp.40. In the other hand, Enterobacter cancerogenus included in the Enterobacteriaceae, bacterial group that play a role in causing microbiologically induced corrosion41,42.
Desulfobacterium sp., Desulfobulbus elongatus and Desulfobacter sp., found in biofilm are bacteria that belongs to the group of sulfate-reduced bacteria. Ito et al.43 showed that different molecular analyses confirmed that Desulfobulbus was found to be the numerically important members of SRB in a wastewater biofilm with Desulfobulbus elongatus was the most frequently found. Sulfate-reduced bacteria group are strict anaerobes that often found in biotopes where conditions can temporarily exist44. Those bacteria do not aggregate on the surface until an anaerobic space is formed and defend themselves against free oxygen by several defense strategies include absorbing substances containing non free oxygen, aerotaxis and enzymatic systems that is dedicated to the reduction and the elimmination of oxygen and its reactive species44,45. Sulfate-reducing bacteria are heterotrophic bacteria that require organic compounds as nutrient source and utilize H2, lactate and acetate as electron donor40. Chemically, sulfate-reducing bacteria can reduce sulfate (SO4)2 to sulphide (S2–) and produce compounds such as hydrogen suphide (H2S) or iron suphide (FeS)46. Eventhough there are few in number as shown in Fig. 3, sulfate-reducing bacteria have been reported as the major part in biocorrosion of metallic surfaces and involved in the number of microbial corrosion problems on various systems and alloys47.
Although be in a consortium of biofilm that potential to cause corrosion, some microbes residing in those biofilm are also known to have a role as a pathogenic bacteria. B. caryophylli is pathogenic for carnations and causes onion rot48 whereas, Xenorhabdus bacterial group are known to be symbiotically associated with the entomopathogenic nematode Steinernema49. Other bacteria which are pathogenic to humans are Burkholderia pseudomallei which is an important cause of acute fulminant pneumonia, septicaemia and cystic fibrosis50, Yersinia pseudotuberculosis that is causing gastroenteritis and extra-intestinal infections51 and Enterobacter cancerogenus that has been found to be the cause of osteomyelitis52. The presence of those microbes in the biofilm, in addition to being ubiquitous in nature such as the genus of Burkholderia that occupy remarkably diverse ecological niches, ranging from contaminated soils to the respiratory tract of humans53, it can be caused also by various pollutions due to many kinds of anthropogenic activities such as domestic, agriculture, animal husbandry and industry at the upper part of the water source.
CONCLUSION
Biofilm formation depended on carbon source and its biodegradability. Bacterial community of biofilm from saguling hydro power has the ability evenly in the use of diverse carbon sources consistently. Carbon sources used by biofilm community at the high rate of reparation were N-acetyl-D-glucosamine from carbohydrates group, 2-hydroxybenzoic acid from carboxylic and ketonic acids, glycogen from polymers group and L-serine from the amino acid groups. In addition, besides being used with a high respiration rate, N-acetyl-D-glucosamine and 2-hydroxybenzoic acid also being used in large quantities for 168 h of incubation time. In this community, there were SRB group as the main group that causes microbial corrosion.
SIGNIFICANCE STATEMENTS
The information regarding the type and the rate of use of carbon sources can be used as a marker to identify metabolic pathways traversed by bacteria that involved in biofilm formation.
ACKNOWLEDGMENT
This study was funded by grants from ITB Research Program to Dea Indriani Astuti and BPP-DNDIKTI Scholarship to Keukeu Kaniawati Rosada.
REFERENCES
- Beech, I., H. Flemming and V. Scotto, 2000. Simple Methods for the Investigation of of the Role of Biofilms in Corrosion. In: Microbially Influenced Corrosion of Industrial Materials, Beech, I.B., A. Bergel, A. Mollica, H.C. Flemming, V. Scotto and W. Sand (Eds.). Biofilm Publication, USA., pp: 1-27.
- Beech, I.B., 2004. Corrosion of technical materials in the presence of biofilms-current understanding and state-of-the art methods of study. Int. Biodeterioration Biodegradation, 53: 177-183.
CrossRefDirect Link - Javaherdashti, R., 2009. A brief review of general patterns of MIC of carbon steel and biodegradation of concrete. IUFS J. Biol., 68: 65-73.
Direct Link - Coetser, S.E. and T.E. Cloete, 2005. Biofouling and biocorrosion in industrial water systems. Critical Rev. Microbiol., 31: 213-232.
CrossRefDirect Link - Flemming, H.C., T.R. Neu and D.J. Wozniak, 2007. The EPS matrix: The house of biofilm cells. J. Bacteriol., 189: 7945-7947.
CrossRefDirect Link - Horemans, B., P. Breugelmans, J. Hofkens, E. Smolders and D. Springael, 2013. Environmental dissolved organic matter governs biofilm formation and subsequent linuron degradation activity of a linuron-degrading bacterial consortium. Applied Environ. Microbiol., 79: 4534-4542.
CrossRefDirect Link - Czaczyk, K. and K. Myszka, 2007. Biosynthesis of Extracellular Polymeric Substances (EPS) and its role in microbial biofilm formation. Polish J. Environ. Stud., 16: 799-806.
Direct Link - Lee, J.W., W.G. Yeomans, A.L. Allen, F. Deng, R.A. Gross and D.L. Kaplan, 1999. Biosynthesis of novel exopolymers by Aureobasidium pullulans. Applied Environ. Microbiol., 65: 5265-5271.
Direct Link - Santos, S.M., J.M. Carbajo and J.C. Villar, 2013. The effect of carbon and nitrogen sources on bacterial cellulose production and properties from Gluconacetobacter sucrofermentans CECT 7291 focused on its use in degraded paper restoration. BioResources, 8: 3630-3645.
Direct Link - Allan, V.J.M., M.E. Callow, L.E. Macaskie and M. Paterson-Beedle, 2002. Effect of nutrient limitation on biofilm formation and phosphatase activity of a Citrobacter sp. Microbiology, 148: 277-288.
CrossRefDirect Link - Edwards, R.A. and D.M. Schifferli, 1997. Differential regulation of fasA and fasH expression of Escherichia coli 987P fimbriae by environmental cues. Mol. Microbiol., 25: 797-809.
CrossRefDirect Link - Beech, I.B. and J. Sunner, 2004. Biocorrosion: Towards understanding interactions between biofilms and metals. Curr. Opin. Biotechnol., 15: 181-186.
CrossRefDirect Link - Rosada, K.K., N.F. Afianti, D.I. Astuti, G. Suantika and P. Aditiawati, 2014. Bacterial community structures of planktonic cells and biofilm at saguling hydro ower using Denaturing Gradient Gel Electrophoresis (DGGE) J. Biol. Sci., 14: 414-424.
CrossRefDirect Link - Gryta, A., M. Frac and K. Oszust, 2014. The application of the Biolog EcoPlate approach in ecotoxicological evaluation of dairy sewage sludge. Applied Biochem. Biotechnol., 174: 1434-1443.
CrossRefDirect Link - Uehara, T. and J.T. Park, 2004. The N-acetyl-D-glucosamine kinase of Escherichia coli and its role in murein recycling. J. Bacteriol., 186: 7273-7279.
CrossRefDirect Link - Deveryshetty, J., V. Suvekbala, G. Varadamshetty and P.S. Phale, 2007. Metabolism of 2-, 3-and 4-hydroxybenzoates by soil isolates Alcaligenes sp. strain PPH and Pseudomonas sp. strain PPD. FEMS Microbiol. Lett., 268: 59-66.
CrossRefDirect Link - Wilson, W.A., P.J. Roach, M. Montero, E. Baroja-Fernandez and F.J. Munoz et al., 2010. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev., 34: 952-985.
CrossRefDirect Link - Lee, J.C.Y., A. Tsoi, G.D. Kornfeld and I.W. Dawes, 2013. Cellular responses to l-serine in Saccharomyces cerevisiae: Roles of general amino acid control, compartmentalization and aspartate synthesis. FEMS Yeast Res., 13: 618-634.
CrossRefDirect Link - Zhang, Y., Z. Lin, Q. Liu, Y. Li and Z. Wang et al., 2014. Engineering of Serine-Deamination pathway, Entner-Doudoroff pathway and pyruvate dehydrogenase complex to improve poly (3-hydroxybutyrate) production in Escherichia coli. Microbial Cell Factories, Vol. 13.
CrossRef - Rickard, A.H., A.J. McBain, A.T. Stead and P. Gilbert, 2004. Shear rate moderates community diversity in freshwater biofilms. Applied Environ. Microbiol., 70: 7426-7435.
CrossRefDirect Link - Prakash, B., B.M. Veeregowda and G. Krishnappa, 2003. Biofilms: A survival strategy of bacteria. Curr. Sci., 85: 1299-1307.
Direct Link - Kokare, C.R., S. Chakraborty, A.N. Khopade and K.R. Mahadik, 2009. Biofilm: Importance and applications. Ind. J. Biol. Technol., 8: 159-168.
Direct Link - Tolker-Nielsen, T. and S. Molin, 2000. Spatial organization of microbial biofilm communities. Microb. Ecol., 40: 75-84.
Direct Link - Manga, S.S., S.B. Oyeleke, A.D. Ibrahim, A.A. Aliero and A.I. Bagudo, 2012. Influence of bacteria associated with corrosion of metals. Cont. J. Microbiol., 6: 19-25.
Direct Link - Lee, W., Z. Lewandowski, S. Okabe, W.G. Characklis and R. Avcl, 1993. Corrosion of mild steel underneath aerobic biofilms containing sulfate‐reducing bacteria part I: At low dissolved oxygen concentration. Biofouling, 7: 197-216.
CrossRefDirect Link - Jung, M.Y., J.S. Kim and Y.H. Chang, 2009. Bacillus acidiproducens sp. nov., vineyard soil isolates that produce lactic acid. Int. J. Syst. Evol. Microbiol., 59: 2226-2231.
CrossRefDirect Link - Muyzer, G. and A.J.M. Stams, 2008. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol., 6: 441-454.
CrossRefDirect Link - Eker, B. and E. Yuksel, 2005. Solutions to corrosion caused by agricultural chemicals. Trakia J. Sci., 3: 1-6.
Direct Link - Philips, S., H.J. Laanbroek and W. Verstraete, 2002. Origin, causes and effects of increased nitrite concentrations in aquatic environments. Rev. Environ. Sci. Biotechnol., 1: 115-141.
CrossRefDirect Link - Zhu, X.Y., J. Lubeck and J.J. Kilbane, 2003. Characterization of microbial communities in gas industry pipelines. Applied Environ. Microbial., 69: 5354-5363.
CrossRefDirect Link - Gavrila, L., D. Gavrila and A.I. Simion, 2003. Microbiologically induced corrosion and it's mitigation in cooling cycles. Sci. Study Res., 5: 127-135.
Direct Link - Nemati, M., G.E. Jenneman and G. Voordouw, 2001. Impact of nitrate-mediated microbial control of souring in oil reservoirs on the extent of corrosion. Biotechnol. Prog., 17: 852-859.
CrossRefDirect Link - Liao, J., H. Fukui, T. Urakami and H. Morisaki, 2010. Effect of biofilm on ennoblement and localized corrosion of stainless steel in fresh dam-water. Corrosion Sci., 52: 1393-1403.
CrossRefDirect Link - Beech, I.B. and C.C. Gaylarde, 1999. Recent advances in the study of biocorrosion-An overview. Rev. Microbiol., 30: 177-190.
CrossRefDirect Link - Bermont-Bouis, D., M. Janvier, P.A.D. Grimont, I. Dupont and T. Vallaeys, 2007. Both sulfate‐reducing bacteria and Enterobacteriaceae take part in marine biocorrosion of carbon steel. J. Applied Microbiol., 102: 161-168.
CrossRefDirect Link - Parot, S., O. Nercessian, M.L. Delia, W. Achouak and A. Bergel, 2009. Electrochemical checking of aerobic isolates from electrochemically active biofilms formed in compost. J. Applied Microbiol., 106: 1350-1359.
CrossRefDirect Link - Ito, T., S. Okabe, H. Satoh and Y. Watanabe, 2002. Successional development of sulfate-reducing bacterial populations and their activities in a wastewater biofilm growing under microaerophilic conditions. Applied Environ. Microbiol., 68: 1392-1402.
Direct Link - Dolla, A., M. Fournier and Z. Dermoun, 2006. Oxygen defense in sulfate-reducing bacteria. J. Biotechnol., 126: 87-100.
CrossRefDirect Link - Sun, C., J. Xu and F. Wang, 2011. Interaction of sulfate-reducing bacteria and carbon steel Q 235 in biofilm. Ind. Eng. Chem. Res., 50: 12797-12806.
CrossRefDirect Link - Lane, R.A., 2005. Under the microscope: Understanding, detecting and preventing microbiologically influenced corrosion. J. Failure Anal. Prevention, 5: 10-12.
CrossRefDirect Link - Enning, D. and J. Garrelfs, 2014. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Applied Environ. Microbiol., 80: 1226-1236.
CrossRefDirect Link - Ballard, R.W., N.J. Palleroni, M. Doudoroff, R.Y. Stanier and M. Mandel, 1970. Taxonomy of the aerobic pseudomonads: Pseudomonas cepacia, P. marginata, P. alliicola and P. caryophylli. Microbiology, 60: 199-214.
CrossRefDirect Link - Tailliez, P., S. Pages, N. Ginibre and N. Boemare, 2006. New insight into diversity in the genus Xenorhabdus, including the description of ten novel species. Int. J. Syst. Evolut. Microbiol., 56: 2805-2818.
CrossRefDirect Link - O'Carroll, M.R., T.J. Kidd, C. Coulter, H.V. Smith, B.R. Rose, C. Harbour and S.C. Bell, 2003. Burkholderia pseudomallei: Another emerging pathogen in cystic fibrosis. Thorax, 58: 1087-1091.
CrossRefDirect Link - Galindo, C.L., J.A. Rosenzweig, M.L. Kirtley and A.K. Chopra, 2011. Pathogenesis of Y. enterocolitica and Y. pseudotuberculosis in Human Yersiniosis. J. Pathogens, Vol. 2011.
CrossRef - Garazzino, S., A. Aprato, A. Maiello, A. Masse, A. Biasibetti, F.G. De Rosa and G. Di Perri, 2005. Osteomyelitis caused by Enterobacter cancerogenus infection following a traumatic injury: Case report and review of the literature. J. Clin. Microbiol., 43: 1459-1461.
CrossRefDirect Link - Coenye, T. and P. Vandamme, 2003. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol., 5: 719-729.
CrossRefDirect Link