
Imene Benyettou
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Omar Kharoubi
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Nouria Hallal
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Hadj Ali Benyettou
Labortory of Medical Analysis, Oran, Algeria
Kaddour Tair
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Mansoria Belmokhtar
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Abdelkader Aoues
Laboratory of Experimental Biotoxicology, Biodepollution and Phytoremediation, Faculty of Life and Natural Sciences,
University of Oran 1 Ahmed Ben Bella, Oran, Algeria
Mehmet Ozaslan
Department of Biology, University of Gaziantep, 27310 Gaziantep, Turkey
LiveDNA: 90.581
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 3 | Page No.: 106-117







ABSTRACT
Background and Objective: Aluminium (Al) is a widely recognized neurotoxin that inhibits more than 200 biologically important functions inducing severe behavioral abnormalities, impaired cognitive functions and metabolic dysfunction. The present study was designed to investigate oxidative stress of aluminium induced toxicity in developing rat brain and enlarged to investigate the possible ameliorating role of omega-3 and omega-6 fatty acids with low and high ratio. Methodology: Aluminium was administered intraperitoneally (100 mg kg–1 b.wt., twice a week) to female adult rats. Postnatally (30 days), pups were given orally by gavage the low and high ratio of omega-6/omega-3 for 5 weeks. The antioxidative and protective properties of omega-6/omega-3 ratio against the effects of aluminium toxicity (Al) on behavior, redox status and leptin level was analysed by using the one way ANOVA test. Results: The results revealed that AlCl3 significantly increased (p<0.05) the level of Thiobarbituric Acid Reactive Substances (TBARS), Nitric oxide (NO), the antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT) and Glutathione Peroxidase (GPx), while Glutathione Reductase (GR) was significantly decreased (p<0.05) in the cerebral cortex in intoxicated pup rats. Moreover, the leptin concentration was significantly increased (p<0.05) in cortex. On the other hand, the results exhibited that, omega-6/omega-3 when given in low or high dose were able to ameliorate the mentioned parameters approaching them to the normal ranges. Conclusion: It is concluded that aluminium may be attributed to induce disturbance of locomotor and stereotype behavior, anxiety, disturb antioxidant system and leptin level. Furthermore, the results suggested that the omega-3 and omega-6 could be able to antagonize Al neurotoxicity perhaps by its antioxidant properties.
PDF Abstract XML References Citation
Received: January 22, 2017;
Accepted: February 28, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Imene Benyettou, Omar Kharoubi, Nouria Hallal, Hadj Ali Benyettou, Kaddour Tair, Mansoria Belmokhtar, Abdelkader Aoues and Mehmet Ozaslan, 2017. Aluminium-Induced Behavioral Changes and Oxidative Stress in Developing Rat Brain and the Possible Ameliorating Role of Omega-6/Omega-3 Ratio. Journal of Biological Sciences, 17: 106-117.
DOI: 10.3923/jbs.2017.106.117
URL: https://scialert.net/abstract/?doi=jbs.2017.106.117
DOI: 10.3923/jbs.2017.106.117
URL: https://scialert.net/abstract/?doi=jbs.2017.106.117
INTRODUCTION
Aluminum (Al) is the third most abundant element therein, after oxygen and silicon. The human body gets Al access via gastrointestinal and respiratory tracts1 from many sources as it presents naturally in food and water and is added to drinking water, processed foods, cosmetics, toothpaste, antiperspirants and adjuvants in various parenteral preparations and pharmaceuticals agents2,3. Aluminum (Al) has been attributed no role by nature in living process but it disrupts the pro-oxidant/anti-oxidant balance of tissues leading to various biochemical, physiological and mental dysfunctions4. It is implicated in several neurodegenerative disorders including Alzheimer’s disease5, Parkinson’s disease and dementia6. Exposure to high levels of Al leads to neurofibrillary degeneration and the concentration of Al has been found to be augmented in degenerating neurons7. The brain is a target of Al toxicity which can alter Blood-Brain Barrier (BBB) mediating its transport to the brain8 and gets deposited in the cortex9 by altering the physiological ligands present at these barriers10.
The brain is particularly vulnerable to free radicals due to its low content of glutathione, the high proportion of polyunsaturated fatty acids (PUFAs) in its membranes and its metabolism of about 20% of total body oxygen11.
The recent studies suggested that Al exposure also results in the production of free radicals12-13 and have sever toxic manifestations on the Central Nervous System (CNS) that increased the susceptibility of CNS to lipid peroxidation (LPO)14-16. Moreover a decrease of total glutathione disturb the antioxidant enzymes activities in brain of rats17.
Before considering the effects of PUFAs on brain-mediated functions, the effects of PUFAs on brain structural components must be elucidated. Fatty acids and lipids are major components in brain structure. Very high levels of fatty acids and lipids can be found in two structural components; in the neuronal membrane and myelin sheaths18.
Actually, the brain and other neural tissues are particularly rich in long-chain polyunsaturated fatty acids (PUFAs), which serve as specific precursors for eicosanoids that play important roles in normal CNS cells development and function19-21.
Fatty acids considered as strong antioxidants are important biological constituents with metabolic, structural and signaling roles. The developing fetus requires substantial amounts of fatty acids to support rapid cellular growth and activity. The most biologically important polyunsaturated fatty acids (PUFAs) are omega-3 and omega-6 PUFAs which are eicosapentaenoic acid (EPA; 20:5n-3), docosahexaenoic acid (DHA; 22:6n-3), dihomo gamma linolenic acid (20:3n-6) and arachidonic acid (AA; 20:4n-6)22. Dietary balance between omega-6 and omega-3 PUFAs plays a major role in the regulation of brain function. Some studies suggest that omega-3 PUFA deficiency exacerbates the sensitivity of individuals to emotional disturbances, such as chronic stress. Omega-3 is central component of glial and neuronal membrane phospholipids and takes part in brain membrane remodeling and synthesis and in signal transduction23.
In view of the aforementioned considerations, this study was carried out to investigate the possible protective ability of omega-6 PUFAs/omega-3 PUFAs ratio on Al induced altered neuromuscular coordination and caused anxiety, oxidative stress and biochemical changes along with oxidative damage in developing rat brain.
MATERIALS AND METHODS
Chemicals: Aluminum chloride (AlCl3) was obtained from agents of sigma chemicals (St. Louis, France). All other reagents used were of high quality and analytic grade.
Omega-3 Magnum EPA/DHA Be-Life capsules (each capsule has 1000 mg of fish oil, 180 mg EPA, 120 mg DHA).
Omega-6 capsules evening primrose 1000 omega 6-9 Be-life (each capsule has polyunsaturated 840 mg EPA, oleic acid 972 mg, L.A. 6748 mg and G.L.A. 692 mg).
Experimental design: This study was carried out on 32 healthy female rats procured from central animal house, Laboratory of Experimental Biotoxicology of the University of Oran1 Ahmed Benbella, Algeria during the year 1999. The animals were housed in standard conditions with free access to food and water (12 h light/dark, 22±2°C). All the procedure performed on animals were approved and conducted in accordance with the National Institute of health Guide (Reg. No. 488/160/1999/CPCSEA).
The animals were 6 months old and their body weight was in the range of 160-200 g. They were acclimatized under hygienic conditions and were fed on standard rat feed and water ad libitum.
The females were checked daily for vaginal plaque and the day on which the vaginal plaque was seen was marked as day one.
The female rats were separated, divided into 4 groups and placed in separate cages for the gestation period. The rat litter obtained from the parents was allowed to survive on their mother’s milk for 4 weeks.
The litter size of the control subgroup was 8-12 rats which was decreased to 6-8 rats after aluminium treatment.
Weekly weight changes were recorded and the dose was adjusted accordingly. The rats were monitored for their health, general behaviour and daily food intake.
The animals were assigned to two groups: group A (the adult rats: 32 rats) served as developed brain model and group B (the rat pups: 48 rats) as developing brain model.
Group A was further divided into four subgroups: Subgroup A1 served as control and was given free access to water and diet, Subgroup A2 was treated with AlCl3 (100 mg kg–1 b.wt.) by intraperitoneal injection twice a week over a period of four weeks., subgroup A3 was administered orally with the low omega-6/omega-3 ratio (0.168) (0.168 = 2.1 and 12.5 g kg–1 b.wt., respectively) by gavage on alternate day as protective agent over a period of 5 weeks and with an intraperitoneally injection of aluminum chloride (100 mg kg–1 b.wt.,) over a period of 4 weeks and the subgroup A4 was received orally the high omega-6/omega-3 ratio (2.1) (2.1 = 10. 5 and 5 g kg–1 weight, respectively) by gavage on alternate day as protect agent over a period of 5 weeks and with an intraperitoneally injection of aluminum chloride (100 mg kg–1 b.wt.) over a period of 4 weeks.
Similarly in group B, the subgroup B1 (issued from control female) served as control, the subgroup B2 (issued from intoxicated female) received aluminium as aluminium chloride feeding mothers for 4 weeks.
The subgroup B3 (issued from intoxicated female) received aluminium as aluminium chloride feeding mothers for 4 weeks and was administered orally with the low omega-6/omega-3 ratio (0.168) by gavage on alternate day as protective agent over a period of 5 weeks.
The subgroup B4 (issued from intoxicated female) received aluminium as aluminium chloride feeding mothers for 4 weeks and was received orally the high omega-6/omega-3 ratio (2.1) by gavage on alternate day as protective agent over a period of 5 weeks.
The subgroup B5 (issued from intoxicated and treated with the low omega-6/omega-3 ratio female) received aluminium as aluminium chloride and low omega-6/omega-3 ratio feeding mothers for 4 weeks.
The subgroup B6 (issued from intoxicated and treated with the high omega-6/omega-3 ratio female) received aluminium as aluminium chloride and high omega-6/omega-3 ratio feeding mothers for 4 weeks.
Brain samples: At the end of the experiment and overnight fasting, the animals were anesthetized with sodium pentobarbital (60 mg kg–1 b.w.).
For evaluation of oxidative status, the brain was removed, washed with normal saline and all the extraneous materials were removed before weighing and the organ weight ratio was estimated. It was kept at ice-cooled conditions all the time. The brain was then gently removed, minced with ice-cold saline, blotted on filter paper and cortex was separated than any other area of the central nervous system24 and afterwards homogenized in 50 mmol L–1 phosphate buffer (pH 7.4). Homogenate was stored at -70°C until biochemical assay.
Body weight: A careful record of body weight changes of both group A and group B was kept throughout the study. The animals were weighed at the beginning of the experiment, then once a week and finally before sacrificing them.
Behavioral tasks: Different groups of rats were investigated for their spatial learning abilities. Ambulation, stereotypic behavior and anxiety were monitored in the open field and dark-light tests. Scoring procedures are briefly described below.
Locomotor activity (open field test (OFT)): The open field test provides simultaneous measures of locomotion and anxiety25. The open field used was a square wooden arena measured (90×90×25 cm). The floor was divided by white lines into 36 smaller squares (15×15 cm). The open field maze was cleaned between each rat to avoid odor cues. The rats were carried to the test room in their home cages and tested once at a time for 10 min each. The locomotor activity (horizontal) defined as number of squares crossed and rearing frequency (vertical) defined as number of times the animals stood on their hind legs26-27 were evaluated. Other elements of exploratory activity such rearing, grooming and sniffing were carefully observed and time spent performing each behavior was recorded. These parameters were defined as follows: rearing (standing on hind legs with paws pressed against the wall of the arena); sniffing (continuous placing nose against the floor for at least 2 sec) and grooming (using paws or tongue to clean/scratch body)28.
Light-dark transition task: The light and dark exploration tasks represent a natural conflict between the tendencies of rats to explore a novel environment versus the tendency of rats to avoid a brightly lit open field. The light-dark box apparatus consists of a rectangular box (44×8.5×25 cm) divided equally into a light, open compartment, connected by a door (17 cm in height) leading to a dark closed compartment in which the animal is placed. Each animal was placed facing the side away from the door and then released. During 15 min29, the time spent in dark and light compartments, respectively, was measured to evaluate degrees of anxiety.
Biochemical investigation: In the brains samples, the following parameters were investigated: TBARS, SOD, catalase, GPx , GR, NO and leptin.
Lipid peroxide assay: Lipid peroxide formation was assayed by the method of Ohkawa et al.30. The results were expressed as nmol of malondialdehyde (MDA) mg–1 of protein. The MDA is a degradation product of peroxidized lipids. The pink color of the TBA-MDA chromophore has been taken as an index of LPO (absorption maximum at 532 nm).
Determination of superoxide dismutase (SOD) activity: Superoxide dismutase (SOD) activities (total SOD: Mn SOD and Cu/Zn SOD) were determined using the pyrogallol assay following the procedure described by Marklund and Marklund31, based on the competition between pyrogallol oxidation by superoxide radicals and superoxide dismutation by SOD. Briefly, 100 μL of 20 mM pyrogallol solution (in HCl 0.01 M) was added to 2.8 μL of tris buffer (containing 50 mM of tris buffer and 1 mM of ethylene diamine tetra acetic) and 100 μL of tissues supernatants and then mixed. Measurement was taken at 420 nm after 1 min 30 sec and 3 min 30 sec using a visible spectrophotometer (DU 720 UV, Beckman Coulter, Palo Alto, CA). The percentage of inhibition of pyrogallol autoxidation was calculated and 1 U of enzymatic activity was defined as the quantity of enzyme necessary to achieve a 50% inhibition of autoxidation at 25°C mg–1 of protein.
Catalase activity (CAT): Catalase was determined by means of the UV spectrophotometric method described by Luck32. H2O2 was used as substrate. The UV absorption of the H2O2 solution was recorded at 240 nm after reaction of H2O2 with catalase. From the decrease in optical density, the enzyme activity was calculated. The amount of H2O2 decomposed was calculated on the basis of the molar extinction coefficient of H2O2 (0.71 L mol–1 cm–1) and the results were expressed as mmol of H2O2 decomposed min–1 mg–1 protein.
Glutathione peroxidase activity (GPx): Glutathione peroxidase (GPX) is an enzyme family with peroxides activity whose main biological role is to protect the organism from oxidative damage and help to prevent lipid peroxidation of cellular membrane by removing the free peroxides in the cells. The enzyme was quantitated by Cayman kit.
Cayman’s GPx assay measures GPx activity indirectly by a coupled reaction with Glutathione Reductase (GR). Oxidized glutathione (GSSG), produced upon reduction of hydroperoxide by GPx, is recycled to its reduced state by GR and NADPH:
| (1) |
| (2) |
The oxidation of NADPH to NADP+ is accompanied by a decrease in absorbance at 340 nm. Under conditions in which the GPx activity is rate limiting, the rate of decrease in the A340 is directly proportional to the GPx activity in the sample. The Cayman GPx assay Kit can be used to measure all of the glutathione-dependent peroxidases in plasma, erythrocyte lysates, tissue homogenates and cell lysates.
Glutathione reductase activity (GR): Glutathione reductase is also called GSR; it is an enzyme that reduces glutathione disulfide (GSSG) to sulfhydryl form GSH, which is an important cellular antioxidant. The enzyme activity was quantitated by Cayman kit.
Cayman’s Glutathione Reductase Assay Kit measures GR activity by measuring the rate of NADPH oxidation :
| (3) |
The oxidation of NADPH to NADP+ is accompanied by a decrease in absorbance at 340 nm and is directly proportional to the GR activity in the sample. The Glutathione Reductase Assay Kit can be used to measure GR activity in plasma, erythrocyte lysates, tissue homogenates and cell lysates.
Nitrite estimation: Nitrite estimation is an indirect measurement of nitric oxide (NO) content in the biological samples. The level of NO was quantitated by cayman kit.
Cayman’s nitrate/nitrite colorimetric assay kit provides an accurate and convenient method for measurement of total nitrate/nitrite concentration in a simple two-step process. The first step is the conversion of nitrate to nitrite utilizing nitrate reductase. The second step is the addition of the Griess reagents which convert nitrite into a deep purple azo compound. Photometric measurement of the absorbance due to azo chromophore accurately determines nitrite (NO2‾) concentration.
The protein content in the different samples was estimated by the method of Lowry et al.33.
Leptin estimation: Leptin is a protein hormone with important effects in metabolism and regulating body weight. It is a single chain 16 kDa protein consisting of 146 amino acid residues and encoded by the obese (ob) gene. Leptin is expressed predominantly by adipocytes, small amounts of leptin are also secreted by cells in the epithelium of stomach and in the placenta. Leptin’s effect on body weight is mediated through effects on hypothalamic centers, where leptin receptors are highly expressed. Leptin has a dual action, it decreases the appetite and increases energy consumption. The level of brain leptin was quantitated by mouse/rat leptin enzyme immunoassay kit (SPI bio).
This Enzyme Immunometric Assay (EIA) is based on a double-antibody sandwich technique specific for mouse and rat leptin.
Statistical analysis: Results were expressed as the Mean±SEM in each group. Groups were compared together by one way analysis of variance (ANOVA) test followed by tukey-test. The acceptance level of significance was p<0.05.
RESULTS
Body and brain weight: In aluminium treated pup rats a significant increase (p<0.05) in body weight as well as brain weight was observed as compared to their control group (Table 1).
The results in Table 1 show the positive effect of treatment by omega-6/omega-3 ratio on body and whole brain weight, a significantly increased (p<0.05) gain on body in all treated groups (B3, B4, B5, B6) as compared as B1 and B2 groups. Moreover, in the brain weight, the results showed a significantl increase (p<0.05) in B5 and B6 groups as compared to B2 group.
On the other hand the treatment by omega-6/omega-3 ratio on brain weight in B3 and B4 groups did not show a positive effect, the brain weight in mentioned groups decreased as compared to aluminium treated group.
Effect of treatment on behavioural parameters: No significant difference was found between the locomotor activities score count of B2 and B3 groups compared to control (Fig. 1). However, in B4 group, the total locomotor activity score was significantly increased (by +92%) as compared to B1 and B2 groups. But a significant decrease (p<0.05) in the locomotor activity was observed in animals issued from A3 and A4 mother’s groups.
There was no variation between all groups in the grooming activity counts, except in B6 group where noted a significant decrease value (p<0.05) was compared to control and A1 group (77.1 and 79.4%, respectively) (Fig. 1).
Results from dark-light box test (Fig. 1) showed that Al exposed rat spent less time in dark than light, which has suggested for the anxiety of intoxicated animals. Conversely, B3, B4, B5 and B6 groups spent significant more time in the dark compartment, compared to aluminium intoxicated group.
Lipid peroxidation: Following aluminium treatment for 4 weeks, the MDA levels were significantly elevated (+25%) in the pups group as compared to controls in both cortex (p<0.05).
Co-administered omega-6/omega-3 ratio during AlCl3 intoxication, ameliorate the elevated MDA level in B3, B4, B5 and B6 groups.
Effect of treatment on antioxidant enzymes activities in cortex
SOD and catalase activities: In this study, the evaluation of superoxide dismutase activity was observed in aluminium treatment group B2 as compared to control group, while omega-6/omega-3 ratio exposure significantly reduced (p<0.05) in B3, B5 and B6 groups (Table 2). Treatment with omega-6/omega-3 ratio in B4 group showed a no significant amelioration in superoxide dismutase activity.
Statistically, CAT values remain significantly lower in B3, B4, B5 and B6 groups compared to intoxicated group (Table 2) by 31,9, 72, 7 and 36%, respectively. However, the result in Table 2 explains a significantly increased by 21% in the cortex CAT activity in Al intoxicated animal compared to the normal control group.
| Table 1: | Effect of aluminium and omega-6/omega-3 ratio on body and brain weight |
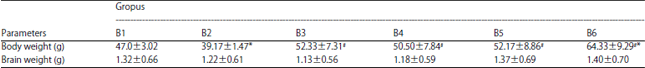 | |
| Values are Mean±SE (n = 8). *p<0.05, All groups were compared to control. #p<0.05, All groups are compared to Al group (Tukey-test) | |
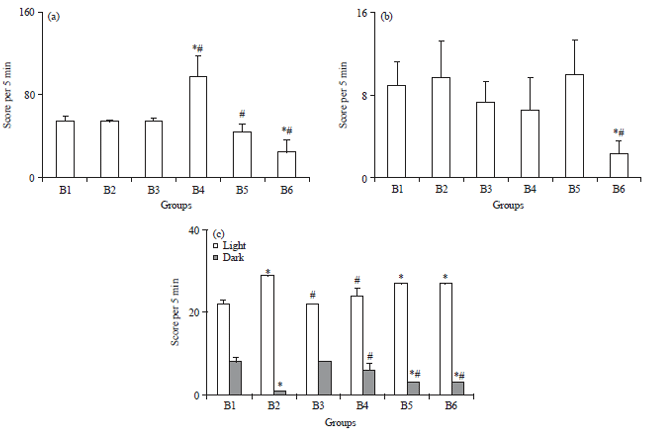 | |
| Fig. 1(a-c): | Effect of Al-exposure on (a) Locomotor activity, (b) Frequency of grooming response and (c) Frequency of sniffing response |
| Values are Mean±SE (n = 8).*p<0.05, All group were compared to control. #p<0.05, All group are compared to Al group (Tukey-test) | |
| Table 2: | Level of TBARS, NO, catalase, SOD, GPx and GR activity in cortex rats |
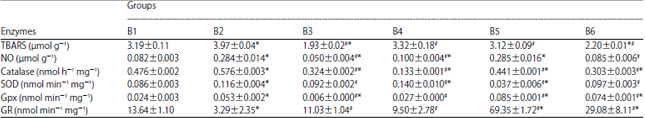 | |
Values are Mean±SE (n = 8). *p<0.05, All groups were compared to control. #p<0.05, All groups are compared to Al group (Tukey-test), TBARS: Thiobarbituric acid reactive substances, NO: Nitric oxide, SOD: Superoxide dismutase, GPx: Glutathione peroxidase, GR: Glutathione reductase | |
GPx and GR activities: Exposure to Al produced changes in GPx and GR activities in the cortex. A significant increase (p<0.05) in GPx and a significant decrease (p<0.05) in GR activities were recorded in intoxicated group compared to control.
Administration of omega-6/omega-3 ratio showed amelioration in GPx and GR by significantly increased (p<0.05) their values in B3, B4, B5 and B6 in cortex when compared to those in Al treated group except, GPx activity in B3 and B4 groups.
Nitrite estimation: The result showed that the is a significant higher (p<0.05) level of nitrite in the brain of AlCl3 treated animals as compared to control group. Nitrite level was found to be significantly decreased (p<0.05) in all omega-6/omega-3 ratio treated groups except B5 group.
Leptin level: The Al group presented in Fig. 2, showed the significantly higher (p<0.05) value of leptin level compared to control; however, B5 showed a very higher value compared to control and Al group by +557 and 77%, respectively. Treatment by omega-6/omega-3 ratio in B3, B4 and B6 groups showed a significant decrease (p<0.05) in leptin level as compared to aluminium intoxicated group.
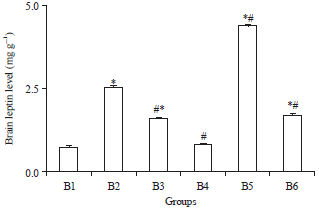 | |
| Fig. 2: | Brain leptin level after 4 weeks of treatment |
Values are Mean±SE (n = 8). *p<0.05, All groups were compared vs. control. #p<0.05, All groups are compared vs. Al group (Tukey-test) | |
DISCUSSION
The present study evaluated the effect of aluminum on the development of different behaviors, the antioxidant status and leptin level in brain cortex and revealed the ability of two feeding ratio of omega-6/omega-3 (low and high ratio) to protect brain against the deleterious effect of aluminum.
The cortex is region known to be particularly susceptible in Alzheimer’s disease and have an important role in learning and memory functions34.
The importance of neurobehavioural studies in risk assessment lies in the fact that behaviour can be regarded as the net output of the sensory, motor and cognitive functions occurring in the nervous system and can serve as potentially sensitive end points of chemically induced neurotoxicity35. Several studies suggest a general decline in learning abilities mediated by aluminium toxicity36. The present study assessed whether omega-6/omega-3 ratio treatment inhibits the Al-affects the motor functions as well as increased anxiety level. Co-administration of omega-6/omega-3 ratio with Al treatment was effective in preventing anxiety-like and motor activities behaviors observed in open field and light/dark box tasks. The method of inducing omega-3 deficiency via diet is a powerful tool to investigate the role of omega-3 in various brain functions. Studies have shown that omega-3-deficient rats exhibited poor learning and memory performances in a variety of tests, such as Morris Water Maze37,38 and olfactory-based learning and memory tasks, mainly in complex (vs. simple) learning39,40. In addition, sensory deficits were evident in those rats, such as visual problems41,42. Study of Salem et al.43 is of specific interest in this aspect, omega-3-deficient rats showed very poor performance in spatial tasks and in olfactory-cued reversal learning tasks. However, Salem et al.43 did not find any difference in the hippocampus gross morphology. Several hypotheses can be offered to explain the finding of poor learning. Omega-3 deficiency induces a significant decrease (p<0.05) in the neuron size in the hippocampus, hypothalamus and cortex44 brain areas that mediate spatial and serial learning. In addition, omega-3 deficiency induces a significant reduction (p<0.05) in cerebral catecholamines45, in glucose transport capacity and glucose utilization in the brain46, in the cyclic AMP level in the hippocampus47 and brain phospholipid synthesis capacity in brain and hypothalamus48,49. Each one of those changes (in the levels of catecholamines, glucose, cAMP and phospholipids) can induce learning deficit. The above-mentioned studies demonstrate two points; firstly, the essentiality of omega-3 fatty acids to the structure and normal function of the brain. Secondly, they demonstrate the importance of the ratio. It is impossible to induce omega-3 deficiency without offsetting the ratio between omega-6 and omega-3 in the diet18. Many studies examined the effects of various fatty acids on learning and memory, but very few examined the ratio between various PUFA. The results of the present study are supported by those described in previous study50 when the ratio of omega-6 and omega-3 may be a key factor in modulating behavioral and neuropharmacological effects of polyunsaturated fatty acids. Therefore, identify the optimal ratio. Obtained results are similar to Yehuda and Carasso50 who found that omega-6/omega-3 ratio was the most effective in improving learning performance (as assessed by the Morris Water Maze and passive avoidance), elevating pain threshold, improving sleep and improving thermoregulation. The ratio omega-6/omega-3 was also able to correct learning deficits induced by the neurotoxins AF64A and 5,7-dihydroxytryptamine51, treatments that decrease the acetylcholine and serotonin brain levels. Similarly, it overcame learning deficit induced by 6-OH-DA (e.g., reduction in brain dopamine level)52. Treatment with a single fatty acid was less successful53,54.
In the present experiment, there was a significant increase (p<0.05) in lipid peroxidation after aluminium exposure in pups group, measured in terms of TBARS levels in the rat brain. Nehru and Anand55 also reported a significant increase (p<0.05) in whole brain thiobarbituric acid reactive substances after stimulation by aluminium salts.
The ionic radii of Al3+ most closely resemble those of Fe3+; therefore, the appearance of Al3+ in Fe3+ sites is probable56. Aluminium is known to be bound by the Fe3+ carrying protein transferrin thus reducing the binding of Fe2+. The increase in free intracellular Fe2+ causes the peroxidation of membrane lipids and thus causes membrane damage.
Moreover, Fleming and Josh57 have reported that the amount of aluminium found in ferritin extracted from Alzheimer’s disease affected brains was 5.6 times higher than in ferritin from matched control samples. The increase may have been due to a general increase in the availability of aluminium to the brain of patients with Alzheimer’s disease and raised the possibility that aluminium releases iron as Fe3+.
The more pronounced effects of aluminium exposure on rat pups as compared to adult rats might be related to the cell biology of the BBB. The BBB is formed by brain capillary endothelial cells. In the late embryonic and early postnatal period, these cells respond to inducing factors found in the brain environment by adopting a set of defined characteristics, including high electrical-resistance tight junctions58. The presence of aluminium at the postnatal stage might affect the formation of BBB in pups, resulting in a higher exposure to the neurotoxic metal compared to the same dose given to the adult group and resulting in increased lipid peroxidation.
The increased LPO is, at least in part, due to an inhibition of SOD in the brain. The result is a substantial increase in the rate of phospholipid peroxidation in brain cells, leading to membrane damage and neuron death.
The SOD presents the first line of defence against superoxide, as it dismutases the superoxide anion to H2O2 and O259. Because the SOD enzyme generates H2O2, it works in collaboration with H2O2 removing enzymes. Catalase converts H2O2 to water and oxygen. Catalase is present in the peroxisomes of mammalian cells and probably serves to destroy H2O2 generated by oxidase enzymes located within these subcellular organelles.
However, the brain is an organ that is especially susceptible to peroxide damage because of several factors, such as high lipid content, high oxygen turnover, low mitotic rate as well as low antioxidant concentration. These factors may explain why aluminium exposure affects the brain more than any other organ.
Glutathione peroxidase (GPX) is an enzyme family with peroxides activity whose main biological role is to protect the organism from oxidative damage and help to prevent lipid peroxidation of cellular membrane by removing the free peroxides in the cell60.
Oxidative stress has been implicated in the pathogenesis of a number of disorders and its extent of injury is generally related to an increase or decrease of one or more free radical scavenging enzymes of which Gpx is one60.
Glutathione reductase (GSR) is an enzyme that reduces Glutathione disulfide (GSSG) to sulfhydryl form GSH, which is an important cellular antioxidant.
In the present study, aluminium exposure produced significant increase (p<0.05) of glutathione peroxidase and a significant decrease (p<0.05) in glutathione reductase activities in aluminium group when compared to control group.
This finding is evidenced by Yuan et al.61 who demonstrated that the positive correlation between Al content and GPX activity in neonatal rats administered daily AlCl3 at dose level of [0, 7 and 35 mg kg–1 b.wt., control, low Al (LA) and high Al (HA), respectively] via intraperitoneal injection for duration of 14 days via drinking water for 6 weeks.
High Al significantly increased (p<0.05) activity of GPX in specific brain regions such as, hippocampus, diencephalon, cerebellum, brain stem as compared with LA and control groups, Gpx activities confirmed Al induced production of the ROS free radical H2O2 and OH which participated in oxidative stress in the brain.
Moreover, the decrease in glutathione reductase activity after aluminium exposure might have resulted from the oxidative modification of genes that control these enzymes. The decrease of the antioxidant versus oxidant ratio plays a crucial role in generating a condition of oxidative stress.
These resuts are in agreement with studies of Savory et al.62 and Johnson et al.63 who showed that aluminum exposure enhanced the neuronal lipid peroxidative damage with concomitant alterations in the enzymatic defense system.
The present experimental results found a significant higher (p<0.05) level of nitrite in the brain of AlCl3 treated animals as compared to normal control group. Nitrite level estimation is an indirect method to estimate the NO level. Nitrite is formed by the oxidation of NO and the reaction is catalyzed by the cytochrome c oxidase in the tissues and multi-copper oxidase ceruloplasmin in the plasma64. Therefore, high content of nitrite indicates elevated level of nitric oxide in the brain. The NO readily reacts with superoxide radicals to form peroxynitrite. Eventually, peroxynitrite can elicit cellular damages by initiating lipid peroxidation and protein adducts formation which leads to neuronal cell damage65. In the present study, omega-6/omega-3 ratio has shown significant decrease (p<0.05) in the NO levels. All of these records are in agreement with a decrease in the total antioxidant capacity.
The dietary supplementation with PUFAs administration has been conducted to evaluate the curative effect of omega-6/omega-3 ratio against AlCl3 induced oxidative stress and dysfunction in brain. Although certain compounds have been tested for the detoxification of aluminium66-68, there is no previous study carried out with omega-6/omega-3 ratio. Taken together, these findings constitute evidence that the antioxidative properties of the omega-6/omega-3 ratio contribute to the prevention of damage induced by AlCl3 in brain rats. The current results indicated an increased TBARS levels in the brain in response to AlCl3 treatment, implying the increased oxidative damage to the tissues. Omega-6/omega-3 ratio treatments returned the increased TBARS levels back to the control levels (Table 2), this result is in accordance with the findings that omega-3 induced reduction of the increased TBARS concentrations in the cerebral cortex (Co) and hippocampus (Hip) of rats intoxicated with AlCl369.
The results of El-Habibi et al.69 exhibited that, omega-3 when given in low or high dose along with AlCl3 was able to ameliorate the lipid peroxidation and antioxidant enzymes activities in dose relating manner approaching them to the normal ranges. Similarly, the dietary supplementation with PUFAs or its chronic administration shortly prior to a severe ischemic insult may ameliorate some of the symptoms associated with cerebral injury probably by increasing antioxidative capacity, lowering lipid peroxidation, inducing chaperon molecules or stabilizing membrane integrity70,71.
In the present study, AlCl3 caused oxidative stress and consequently disturbed the activities of the antioxidant enzymes (SOD, CAT, GPx and GR). After treatment of rats with AlCl3 plus omega-6/omega-3 ratio the activities of these antioxidant enzymes were normalized to their control values. Kondo et al.72 showed who omega-6/omega-3 ratio exercise their activity through the scavenging of hydroxyl, superoxide free radicals and lipid peroxides. The antioxidant activities of omega-6/omega-3 ratio are related to their ability to chelate metal ions and scavenge singlet oxygen, superoxide anions, proxy radicals, hydroxyl radicals and peroxynitrite. Such results were confirmed by the studies of Kim et al.73 who showed that SOD deficiency exacerbated cerebral infarction and that chronic daily administration of omega-3 for 6 weeks increased SOD activity and elevated reduced GSH, leading to effective reduction of the brain lipid peroxidation product (MDA).
It has been reported that leptin levels has also been shown to increase generation of reactive oxygen species74. The present study found significant increase (p<0.05) in leptin level in aluminium intoxicated group and statistically, a significant relationship and a positive correlation between the higher level of leptin and TBARS in aluminium group (R = 0.360) was found. While omega-6/omega-3 ratio treatment significantly decreased (p<0.05) the leptin levels in the cortex brain. Given the fact that leptin play important roles in brain development and activity, several studies evaluated its effects on brain size and function. Leptin receptors are widely distributed in the brain, including expression in the cortex, amygdala, hippocampus and thalamus, with highest
levels in hypothalamic nuclei such as the arcuate nucleus and paraventricular nucleus75,76. The earliest and most robust role of leptin in the brain is its anorexigenic action in the hypothalamus76. Besides further confirming the correlation between leptin levels and brain77, certain studies have also associated low leptin levels with cognitive impairment, particularly AD78,79. The leptin plays an important role to promote significant cognitive improvements80.
There is increasing evidence that omega-6/omega-3 ratio plays a significant role in neurological development, disease prevention and alleviating aluminium neurotoxicity. Omega-3 and 6 fatty acids proved to be beneficial in decreasing the levels of free radicals improving the antioxidant system and neurotransmitter contents. Consequently, the study recommended that attention should be paid to reduce the sources of exposures to aluminium and using omega-3 and 6 as additional to conventional medicine for treatment of neurodegenerative and psychotic disorders.
CONCLUSION
Aluminium has adverse effects on human health. Results of the present study reported that aluminium chloride (AlCl3) is capable to cause marked alterations in some behavior and biochemical parameters by inducing an oxidative damage and inhibiting the antioxidant enzymes activities. Whereas the administration of omega-6/omega-3 (with high and low ratio) combined to aluminium minimize its hazards. In addition, omega-6/omega-3 proved to be beneficial in decreasing free radicals and lipids level and increasing antioxidant enzymes activities. Consequently, aluminium exposure should be reduced by paying more attention to its sources in food, water and personal care products. As well as omega-3 and omega-6 consumption with both high and low ratio could attenuate aluminium toxicity.
SIGNIFICANCE STATEMENT
It is first study about omega-6/omega-3 ratio taken together, these findings constitute evidence that the antioxidative properties of the omega-6/omega-3 ratio contribute to the prevention of damage induced by AlCl3 in brain pup rats.
REFERENCES
- Yokel, R.A., 2000. The toxicology of aluminum in the brain: A review. Neurotoxicology, 21: 813-828.
PubMedDirect Link - Becaria, A., A. Campbell and S.C. Bondy, 2002. Aluminum as a toxicant. Toxicol. Ind. Health, 18: 309-320.
CrossRefDirect Link - Pournourmohammadi, S., P. Khazaeli, S. Eslamizad, A. Tajvar, A. Mohammadirad and M. Abdollahi, 2008. Study on the oxidative stress status among cement plant workers. Hum. Exp. Toxicol. 27: 463-469.
CrossRefDirect Link - Nehru, B. and P. Bhalla, 2006. Reversal of an aluminium induced alteration in redox status in different regions of rat brain by administration of centrophenoxine. Mol. Cell. Biochem., 290: 185-191.
CrossRefDirect Link - Domingo, J.L., 2006. Aluminum and other metals in Alzheimer's disease: A review of potential therapy with chelating agents. J. Alzheimer's Dis., 10: 331-341.
CrossRefDirect Link - Julka, D. and K.D. Gill, 1996. Effect of aluminum on regional brain antioxidant defense status in Wistar rats. Res. Exp. Med., 196: 187-194.
CrossRefPubMedDirect Link - Bharathi, N.M. Shamasundar, T.S. Sathyanarayana Rao, M.D. Naidu, R. Ravid and K.S.J. Rao, 2006. A new insight on Al-maltolate-treated aged rabbit as Alzheimer's animal model. Brain Res. Rev., 52: 275-292.
CrossRefDirect Link - Zatta, P., T. Kiss, M. Suwalsky and G. Berthon, 2002. Aluminium(III) as a promoter of cellular oxidation. Coord. Chem. Rev., 228: 271-284.
CrossRefDirect Link - Platt, B., G. Fiddler, G. Riedel and Z. Henderson, 2001. Aluminium toxicity in the rat brain: Histochemical and immunocytochemical evidence. Brain Res. Bull., 55: 257-267.
CrossRefPubMedDirect Link - Yokel, R.A. and P.J. McNamara, 2001. Aluminium toxicokinetics: An updated mini review. Pharmacol. Toxicol., 88: 159-167.
CrossRefDirect Link - Gupta, V.B., G. Anitha, M.L. Hegda, L. Zecca and R.M. Garruto et al., 2005. Aluminium in Alzheimer's disease: Are we still at a crossroad? Cell. Mol. Life Sci., 62: 143-158.
CrossRefPubMedDirect Link - Lebel, C.P. and S.C. Bondy, 1991. Oxygen radicals: Common mediators of neurotoxicity. Neurotoxicol. Teratol., 13: 341-346.
CrossRefPubMedDirect Link - Colomina, M.T., J.L. Roig, D.J. Sánchez and J.L. Domingo, 2002. Influence of age on aluminum-induced neurobehavioral effects and morphological changes in rat brain. Neurotoxicology, 23: 775-781.
CrossRefPubMedDirect Link - Fraga, C.G., P.I. Oteiza, M.S. Golub, M.E. Gershwin and C.L. Keen, 1990. Effects of aluminum on brain lipid peroxidation. Toxicol. Lett., 51: 213-219.
CrossRefDirect Link - Oteiza, P.I., C.L. Keen, B. Han and M.S. Golub, 1993. Aluminum accumulation and neurotoxicity in Swiss-Webster mice after long-term dietary exposure to aluminum and citrate. Metabolism, 42: 1296-1300.
CrossRefPubMedDirect Link - Sharma, P., Z.A. Shah, A. Kumar, F. Islam and K.P. Mishra, 2007. Role of combined administration of Tiron and glutathione against aluminum-induced oxidative stress in rat brain. J. Trace Elem. Med. Biol., 21: 63-70.
CrossRefPubMedDirect Link - Gendron, T.F., E. Brunette, J.S. Tauskela and P. Morley, 2005. The dual role of prostaglandin E2 in excitotoxicity and preconditioning-induced neuroprotection. Eur. J. Pharmacol., 517: 17-27.
CrossRefDirect Link - Heird, W.C. and A. Lapillonne, 2005. The role of essential fatty acids in development. Annu. Rev. Nutr., 25: 549-571.
CrossRefDirect Link - Matsuoka, Y., T. Furuyashiki, K. Yamada, T. Nagai and H. Bito et al., 2005. Prostaglandin E receptor EP1 controls impulsive behavior under stress. Proc. Natl. Acad. Sci. USA., 102: 16066-16071.
CrossRefDirect Link - Haggarty, P., 2010. Fatty acid supply to the human fetus. Annu. Rev. Nutr., 30: 237-255.
CrossRefDirect Link - Rapoport, S.I., 2001. In vivo fatty acid incorporation into brain phosholipids in relation to plasma availability, signal transduction and membrane remodeling. J. Mol. Neurosci., 16: 243-261.
CrossRefDirect Link - Urano, S., Y. Asai, S. Makabe, M. Matsuo, N. Izumiyama, K. Ohtsubo and T. Endo, 1997. Oxidative injury of synapse and alteration of antioxidative defense systems in rats and its prevention by vitamin E. FEBS J., 245: 64-70.
CrossRefDirect Link - Millan, M.J., 2003. The neurobiology and control of anxious states. Progr. Neurobiol., 70: 83-244.
CrossRefPubMedDirect Link - Suarez-Fernandez, M.B., A.B. Soldado, A. Sanz-Medel, J.A. Vega, A. Novelli and M.T. Fernandez-Sanchez, 1999. Aluminum-induced degeneration of astrocytes occurs via apoptosis and results in neuronal death. Brain Res., 835: 125-136.
CrossRefDirect Link - Colomina, M.T., D.J. Sanchez, M. Sanchez-Turet and J.L. Domingo, 1999. Behavioral effects of aluminum in mice: Influence of restraint stress. Neuropsychobiology, 40: 142-149.
CrossRefDirect Link - Cauli, O. and M. Morelli, 2002. Subchronic caffeine administration sensitizes rats to the motor-activating effects of dopamine D1 and D2 receptor agonists. Psychopharmacology, 162: 246-254.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Marklund, S. and G. Marklund, 1974. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem., 47: 469-474.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Bihaqi, S.W., M. Sharma, A.P. Singh and M. Tiwari, 2009. Neuroprotective role of Convolvulus pluricaulis on aluminium induced neurotoxicity in rat brain. J. Ethnopharmacol., 124: 409-415.
CrossRefDirect Link - Evans, H.L., 1995. Markers of neurotoxicity: From behavior to autoantibodies against brain proteins. Clin. Chem., 41: 1874-1881.
Direct Link - Shi-Lei, S., M.A. Guang-Yu, L.H. Bachelor, Z.Y. Bachelor, H.M. Dong and X.H. Xu, 2005. Effect of naloxone on aluminum-induced learning and memory impairment in rats. Neurol. India, 53: 79-82.
CrossRefPubMedDirect Link - Moriguchi, T., R.S. Greiner and N. Salem Jr., 2000. Behavioral deficits associated with dietary induction of decreased brain docosahexaenoic acid concentration. J. Neurochem., 75: 2563-2573.
CrossRefDirect Link - Wainwright, P.E., 2002. Dietary essential fatty acids and brain function: a developmental perspective on mechanisms. Proc. Nutr. Soc., 61: 61-69.
CrossRefPubMedDirect Link - Greiner, R.S., T. Moriguchi, B.M. Slotnick, A. Hutton and N. Salem Jr., 2001. Olfactory discrimination deficits in n-3 fatty acid-deficient rats. Physiol. Behav., 72: 379-385.
CrossRefDirect Link - Catalan, J., M. Toru, B. Slotnick, M. Murthy, R.S. Greiner and N. Salem Jr., 2002. Cognitive deficits in docosahexaenoic acid-deficient rats. Behav. Neurosci., 116: 1022-1031.
CrossRefDirect Link - Jeffrey, B.G., D.C. Mitchell, R.A. Gibson and M. Neuringer, 2002. n-3 fatty acid deficiency alters recovery of the rod photoresponse in rhesus monkeys. Invest. Ophthalmol. Visual Sci., 43: 2806-2814.
Direct Link - Jeffrey, B.G., D.C. Mitchell, J.R. Hibbeln, R.A. Gibson, A.L. Chedester and N. Salem Jr., 2002. Visual acuity and retinal function in infant monkeys fed long-chain PUFA. Lipids, 37: 839-848.
CrossRefDirect Link - Salem, Jr. N., T. Moriguchi, R.S. Greiner, K. McBride, A. Ahmad, J.N. Catalan and B. Slotnick, 2001. Alterations in brain function after loss of docosahexaenoate due to dietary restriction of n-3 fatty acids. J. Mol. Neurosci., 16: 299-307.
CrossRefDirect Link - Ahmad, A., M. Murthy, R.S. Greiner, T. Moriguchi and N. Salem Jr., 2002. A decrease in cell size accompanies a loss of docosahexaenoate in the rat hippocampus. Nutr. Neurosci., 5: 103-113.
CrossRefDirect Link - Takeuchi, T., Y. Fukumoto and E. Harada, 2002. Influence of a dietary n-3 fatty acid deficiency on the cerebral catecholamine contents, EEG and learning ability in rat. Behav. Brain Res., 131: 193-203.
CrossRefDirect Link - Da Silva, A.X., F. Lavialle, G. Gendrot, P. Guesnet, J.M. Alessandri and M. Lavialle, 2002. Glucose transport and utilization are altered in the brain of rats deficient in n-3 polyunsaturated fatty acids. J. Neurochem., 81: 1328-1337.
CrossRefDirect Link - Nanjo, A., A. Kanazawa, K. Sato, F. Banno and K. Fujimoto, 1999. Depletion of dietary n-3 fatty acid affects the level of cyclic AMP in rat hippocampus. J. Nutr. Sci. Vitaminol., 45: 633-641.
CrossRefDirect Link - Gazzah, N., A. Gharib, M. Croset, P. Bobillier, M. Lagarde and N. Sarda, 1995. Decrease of brain phospholipid synthesis in free-moving n-3 fatty acid deficient rats. J. Neurochem., 64: 908-918.
CrossRefDirect Link - Murthy, M., J. Hamilton, R.S. Greiner, T. Moriguchi, N. Salem and H.Y. Kim, 2002. Differential effects of n-3 fatty acid deficiency on phospholipid molecular species composition in the rat hippocampus. J. Lipid Res., 43: 611-617.
Direct Link - Yehuda, S. and R.L. Carasso, 1993. Modulation of learning, pain thresholds and thermoregulation in the rat by preparations of free purified alpha-linolenic and linoleic acids: Determination of the optimal omega 3-to-omega 6 ratio. Proc. Natl. Acad. Sci. USA., 90: 10345-10349.
Direct Link - Yehuda, S., R.L. Carraso and D.I. Mostofsky, 1995. Essential fatty acid preparation (SR-3) rehabilitates learning deficits induced by AF64A and 5,7-DHT. Neuroreport, 6: 511-515.
Direct Link - Yehuda, S., S. Rabinovitz and D.I. Mostofsky, 1999. Treatment with a polyunsaturated fatty acid prevents deleterious effects of Ro4-1284. Eur. J. Pharmacol., 365: 27-34.
CrossRefDirect Link - Ikemoto, A., M. Ohishi, Y. Sato, N. Hata, Y. Misawa, Y. Fujii and H. Okuyama, 2001. Reversibility of n-3 fatty acid deficiency-induced alterations of learning behavior in the rat: Level of n-6 fatty acids as another critical factor. J. Lipid Res., 42: 1655-1663.
Direct Link - Umezawa, M., K. Kogishi, H. Tojo, S. Yoshimura and N. Seriu et al., 1999. High-linoleate and high-α-linolenate diets affect learning ability and natural behavior in SAMR1 mice. J. Nutr., 129: 431-437.
Direct Link - Nehru, B. and P. Anand, 2005. Oxidative damage following chronic aluminium exposure in adult and pup rat brains. J. Trace Elem. Med. Biol., 19: 203-208.
CrossRefDirect Link - Martin, R.B., J. Savory, S. Brown, R.L. Bertholf and M.R. Wills, 1987. Transferrin binding of Al3+ and Fe3+. Clin. Chem., 33: 405-407.
Direct Link - Fleming, J. and J.G. Josh, 1987. Ferritin: Isolation of aluminum-ferritin complex from brain. Proc. Nalt. Acad. Sci. USA., 84: 7866-7870.
Direct Link - Rubin, L.L. and J.M. Staddon, 1999. The cell biology of the blood-brain barrier. Annu. Rev. Neurosci., 22: 11-28.
CrossRefDirect Link - Fridovich, I., 1983. Superoxide radical: An endogenous toxicant. Annu. Rev. Pharmacol. Toxicol., 23: 239-257.
CrossRefPubMedDirect Link - Kankofer, M., 2002. Superoxide dismutase and glutathione peroxidase activities in bovine placenta: Spectrophotometric and electrophoretic analysis. Rev. Med. Vet., 153: 121-124.
Direct Link - Yuan, C.Y., Y.J. Lee and G.S.W. Hsu, 2012. Aluminum overload increases oxidative stress in four functional brain areas of neonatal rats. J. Biomed. Sci., Vol. 19.
CrossRefDirect Link - Savory, J., M.M. Herman and O. Ghribi, 2003. Intracellular mechanisms underlying aluminum-induced apoptosis in rabbit brain. J. Inorg. Biochem., 97: 151-154.
CrossRefDirect Link - Johnson, V.J., S. Kim and R.P. Sharma, 2005. Aluminum maltolate induces apoptosis and necrosis in neuro-2a cells: Potential role for p53 signaling. Toxicol. Sci., 83: 329-339.
CrossRefDirect Link - Shiva, S., 2013. Nitrite: A physiological store of nitric oxide and modulator of mitochondrial function. Redox Biol., 1: 40-44.
CrossRefDirect Link - Kumar, A., S. Lalitha and J. Mishra, 2013. Possible nitric oxide mechanism in the protective effect of hesperidin against pentylenetetrazole (PTZ)-induced kindling and associated cognitive dysfunction in mice. Epilepsy Behav., 29: 103-111.
CrossRefDirect Link - Yousef, M.I., 2004. Aluminum-induced changes in hemato-biochemical parameters, lipid peroxidation and enzyme activities of male rabbits: Protective role of ascorbic acid. Toxicology, 199: 47-57.
CrossRefDirect Link - Yousef, M.I., A.M. El-Morsy and M.S. Hassan, 2005. Aluminium-induced deterioration in reproductive performance and seminal plasma biochemistry of male rabbits: Protective role of ascorbic acid. Toxicology, 215: 97-107.
CrossRefPubMedDirect Link - Yousef, M.I., K.I. Kamel, M.I. El-Guendi and F.M. El-Demerdash, 2007. An in vitro study on reproductive toxicity of aluminium chloride on rabbit sperm: The protective role of some antioxidants. Toxicology, 239: 213-223.
CrossRefPubMedDirect Link - El-Habibi, E.S.M., W.M. El-Kholy, H.M. Serag and E.S.A.G. El-Mola, 2011. Aluminium-induced brain oxidative stress in male rats and the possible ameliorating role of omega-3. Egypt. J. Exp. Biol. (Zool.), 7: 329-342.
Direct Link - Choi-Kwon, S., K.A. Park, H.J. Lee, M.S. Park and J.H. Lee et al., 2004. Temporal changes in cerebral antioxidant enzyme activities after ischemia and reperfusion in a rat focal brain ischemia model: Effect of dietary fish oil. Dev. Brain Res., 152: 11-18.
CrossRefDirect Link - Cao, D., B. Yang, L. Hou, J. Xu and R. Xue et al., 2007. Chronic daily administration of ethyl docosahexaenoate protects against gerbil brain ischemic damage through reduction of arachidonic acid liberation and accumulation. J. Nutr. Biochem., 18: 297-304.
CrossRefDirect Link - Kondo, T., A.G. Reaume, T.T. Huang, E. Carlson and K. Murakami et al., 1997. Reduction of CuZn-superoxide dismutase activity exacerbates neuronal cell injury and edema formation after transient focal cerebral ischemia. J. Neurosci., 17: 4180-4189.
Direct Link - Kim, G.W., T. Kondo, N. Noshita and P.H. Chan, 2002. Manganese superoxide dismutase deficiency exacerbates cerebral infarction after focal cerebral ischemia/reperfusion in mice: Implications for the production and role of superoxide radicals. Stroke, 33: 809-815.
CrossRefDirect Link - Teryaeva, N.B., 2015. Leptin as a neuroprotector and a central nervous system functional stability factor. Neurosci. Behav. Physiol., 45: 612-618.
CrossRefDirect Link - Couce, M.E., B. Burguera, J.E. Parisi, M.D. Jensen and R.V. Lloyd, 1997. Localization of leptin receptor in the human brain. Neuroendocrinology, 66: 145-150.
CrossRefDirect Link - Meister, B., 2000. Control of food intake via leptin receptors in the hypothalamus. Vitamins Hormones, 59: 265-304.
CrossRefDirect Link - Rajagopalan, P., A.W. Toga, C.R. Jack, M.W. Weiner and P.M. Thompson, 2013. Fat-mass-related hormone, plasma leptin, predicts brain volumes in the elderly. Neuroreport, 24: 58-62.
CrossRefPubMedDirect Link - Power, D.A., J. Noel, R. Collins and D. O'Neill, 2001. Circulating leptin levels and weight loss in Alzheimer's disease patients. Dement. Geriatr. Cogn. Disord., 12: 167-170.
CrossRefPubMedDirect Link - Holden, K.F., K. Lindquist, F.A. Tylavsky, C. Rosano, T.B. Harris and K. Yaffe, 2009. Serum leptin level and cognition in the elderly: Findings from the Health ABC Study. Neurobiol. Aging, 30: 1483-1489.
CrossRefDirect Link - Farr, S.A., W.A. Banks and J.E. Morley, 2006. Effects of leptin on memory processing. Peptides, 27: 1420-1425.
CrossRefDirect Link