
Nanung Agus Fitriyanto
Faculty of Animal Science, Gadjah Mada University, Jalan Fauna No. 3, Bulaksumur, Yogyakarta, Indonesia
LiveDNA: 62.12639
Atik Winarti
Faculty of Animal Science, Gadjah Mada University, Jalan Fauna No. 3, Bulaksumur, Yogyakarta, Indonesia
Fatih Akbar Imara
Faculty of Animal Science, Gadjah Mada University, Jalan Fauna No. 3, Bulaksumur, Yogyakarta, Indonesia
Yuny Erwanto
Faculty of Animal Science, Gadjah Mada University, Jalan Fauna No. 3, Bulaksumur, Yogyakarta, Indonesia
Takashi Hayakawa
Faculty of Applied Biological Sciences, Gifu University, Yanagido 1-1, Gifu, Japan
Tomoyuki Nakagawa
Faculty of Applied Biological Sciences, Gifu University, Yanagido 1-1, Gifu, Japan
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 1 | Page No.: 1-10







ABSTRACT
Background and Objective: A proper management processes for safe and efficient handling of animal manure is important to minimize the possibilities of global environmental problem due to the emission of unpleasant odor. This study was performed to identify strains bacteria screened from the soil in the odorous region of apoultry format Yogyakarta city, Indonesia, to evaluate their capability for growing in ammonium and animal’s urine high content medium and also to investigate the ability to reduce ammonium concentration from the liquid medium. Materials and Methods: The strain is originally obtained from soil sample collected at ammonia high-emitted of chicken hen production area at tropical country Indonesia. The isolation was using a 1/100 nutrient agar with high concentration of (NH4)2SO4 as an organic ammonium stressor. Selected strain was then identified based on morphology and biochemical identifications, as well as molecular identification. The reduction of ammonium was measured from cultured medium using nessler reagent spectroscopically. Results: High ammonium sulfate-responsive, strain LS3K was successfully isolated and identified based on morphology, physiology and biochemical identifications, as well as molecular identification based on 16S rRNA sequence. Based on the morphology, physiological observation and the 16S rRNA gene sequence, strain LS3T was identified as Pseudomonas spp. The content of DNA G+C of the strain was 55.2%. The 16S rRNA gene sequence analysis demonstrated that the isolate is showing high similarity to species of Pseudomonas tolaasii. The cells of strain LS3K are short rods, aerobic and motile. Colonies (l-2 mm in diameter) on plate culture are yellowish, circular and smooth. This bacterium has an optimal growth temperature 30°C. Conclusion: Strain LS3K can tolerate ammonium at 756.77 mM and has the ability for growing in animal’s manure contained a medium that should make it applicable for use in animal waste handling systems as well as for enhancing the capability of deodorization. Ammonium was oxidized 5.648 mg L–1 by strain LS3K during 30 h cultivation period.
PDF Abstract XML References Citation
Received: November 08, 2016;
Accepted: December 08, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nanung Agus Fitriyanto, Atik Winarti, Fatih Akbar Imara, Yuny Erwanto, Takashi Hayakawa and Tomoyuki Nakagawa, 2017. Identification and Growth Characters of Nitrifying Pseudomonas sp., LS3K Isolated from Odorous Region of Poultry Farm. Journal of Biological Sciences, 17: 1-10.
DOI: 10.3923/jbs.2017.1.10
URL: https://scialert.net/abstract/?doi=jbs.2017.1.10
DOI: 10.3923/jbs.2017.1.10
URL: https://scialert.net/abstract/?doi=jbs.2017.1.10
INTRODUCTION
As agricultural based industrial country, intensive and sustainable animal production is crucial for Indonesia. Waste from agrobiological processing, such as animal manure has significantly contributed to global environmental problems. Development of effective and efficient management processes for safe and efficient disposal of waste is crucial. Poultry excreta and dairy feces make a significant contribution to the emission of gaseous ammonia (NH3) as sources of greenhouse gas emission1,2.
Several researchers have investigated for mitigation of odor nuisance from animal waste. It includes dietary and management methods to minimize the generation and volatilization of ammonia from animal waste3, developing a deodorizing system4 and application of some plants to trap the ammonia emissions5. Among them, the addition of biological additives which is employ microorganisms has attracted attention for they are fast in action, ease of application and relatively lower expenses6,7. The treatment for ammonia removing is based on the alteration of volatile N into non-volatile N from comprising of nitrification and denitrification processes by heterotrophic nitrifying and denitrifying bacteria.
The ammonia emissions from animal industries may cause some environmental problems because of their odor, toxicity and contribution to eutrophication and acid rains8. Several potential harmful effects associated with exceeding high concentrations of both oxidized and reduced forms of N in the environment such as respiratory diseases caused by exposure to high levels of ammonia and eutrophication of surface water bodies resulting in harmful algal blooms and decreased water quality. The other adverse effects are nitrate contamination of drinking water, climatic changes associated with increases in nitrous oxide (N2O), ecosystem changes because of higher concentrations of N and soil acid iication through nitrification and leaching9.
In the environment, nitrification-denitrification denote as one of the important system involved in the N cycle. Ammonia-oxidizing bacteria (AOB) and archaea-oxidizing bacteria (AOA) oxidize NH3 to nitrite (NO2–) in various ecosystems. Whereas, NO2– is further oxidized to nitrate (NO3–) by nitrite oxidizing bacteria (NOB)4,8,10,11. The primary grouping of AOB belongs to the subclass β-proteo bacteria and comprises genera such as Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosovibri and Nitrosolobus12. Only a few species of marine Nitrosococcus belong to the Gammaproteo bacteria. The AOA currently was known, belong to the new phylum Thaumarchaeota13.
However, nitrification is also employed during heterotrophic growth of some bacteria, such as Bacillus methylotrophicus14, Alcaligenes faecalis15, Pseudomonas stutzeri 10 and Thiosphaera pantotropha16. Heterotrophic nitrification is thought to be performed in similar ways to the autotrophic process. The ![]() is initially converted to NH2OH by the enzyme ammonia monooxygenase and then followed by the oxidation of NH2OH to NO2– by the hydroxylamine oxidoreductase enzyme (HAO) and then NO2– is further oxidized to NO3– 17.
is initially converted to NH2OH by the enzyme ammonia monooxygenase and then followed by the oxidation of NH2OH to NO2– by the hydroxylamine oxidoreductase enzyme (HAO) and then NO2– is further oxidized to NO3– 17.
Composting is a conventional general method to decompose agriculture crop residues, animal feces, industrial organic waste as well as other organic matter. During aerobic composting, decomposer is important to improve the quality of compost product. Studies on nitrification during composting has been performed8,4. Besides bacteria and archaea, fungi are another microorganism of a decomposer for which the role and development are not clear in the composting system. It is well known that the composting self-heating pilemay reach a temperature too extreme for their survival18. They would thus be eliminated during the thermophilic stage and recovered when the temperature decreases19. Fungi may play a significant role during the maturation stage because of their ability to decompose some slowly biodegradable materials like cellulose and lignin18. Moreover, some fungi exhibit heterotrophic nitrification activity linked to lignin degradation. Their role involved in N transformations during composting needs to be considered20.
To investigate the N losses and NH3 emissions from aerobic treatment, understanding the N mobility and studying the microorganisms involved in N transformations are needed. The objective of this study was to isolate and identified strain microbial from soil at anodorous region that has a deal with nitrogen tolerant and to understand the role of isolated strain in nitrification system.
MATERIALS AND METHODS
Media and culture: A portion of 1/100 diluted nutrient agar with 500 mg L–1 (NH4)2SO4 was used for screening high ammonium-responsive microorganisms. Cultures were then performed aerobically for 7 days at 30°C. The liquid culture was carried out using 100 mL of 1/100 nutrient broth with various (NH4)2SO4 concentrationin 250 mL-Erlenmeyer flasks and cells were cultured at 30°C with a reciprocal shaker (120 rpm). Bacterial growth was monitored at Optical Density (OD) 600.
Screening of ammonium-responsive microorganisms: Indigenous isolates from Yogyakarta city as a tropical area was initially obtained from ammonia high-emitted area of a poultry farm. Soil samples (1.0 g) collected from various spots in a most odorous region were then suspended in 9.0 mL sterile pure water and diluted appropriately. A portion of the cell suspension was spread on a 1/100 nutrient agar plate with 500 mg L–1 (NH4)2SO4. Colonies appearing on the plate were then picked and purified. Each purified colony was inoculated on an agar plate with and without 500 mg L–1 (NH4)2SO4. Microorganisms displaying a proper growth on the agar with (NH4)2SO4 were selected as ammonium-responsive microorganisms. Isolates were purified by plating on 1/100 nutrient broth (0.01% meat extract, 0.01% polypeptone and 0.005% NaCl, pH 7.2) supplemented with 500 mg L–1 (NH4)2SO4 and continued by incubation at 30°C for 48 h in aerobic condition.
Identification of anammonium-responsive bacterium: Morphological, biochemical and physiological characteristic of the ammonium-responsive bacterium were investigated according to Bergey’s Manual of Systematic Bacteriology21. Genomic DNA of the bacterium was extracted by standard methods22. The 16S rRNA was amplified by PCR using universal primers. The purified PCR product was then sequenced using BigDye Terminator v3.1 Cycle Sequencing Kit from Applied Biosystems, USA with the ABI Prism 310 Genetic Analyzer (Applied Biosystems) and analyzed using the BLAST version 2.2.18 (BLASTN) to compare with the public database of DDBJ (http://blast.ddbj.nig.ac.jp).
Genomic DNA isolation: For the genomic DNA isolation, an overnight cell culture was harvested and suspended in DNA extraction buffer (2.5 M NaCl, 500 mM EDTA, 1 M tris-HCl, 10% SDS pH 7.5). The solution was then mixed by inverting the eppendorf tube followed by incubation at 65°C for 5 min and method was repeated twice. After incubation, a portion of phenol:chloroform:isoamyl alcohol (P:C:I) was added to the solution followed by vortex for 5 min and centrifugation at 13000 rpm for 10 min to separate DNA from other cellular contaminants. The top layer of the solution was then transferred to new eppendorf tube and then added by 750 μL 100% ethanol and 30 μL 3 M NaCH3COO and incubated at -20°C for 30 min. Precipitated DNA was collected by centrifugation at 13000 rpm for 10 min, continued by washing with 250 μL 70% ethanol and re-suspended in tris-EDTA (TE) buffer. The isolated DNA was resolved by electrophoresis on a 0.7% agarose gel in TAE buffer at 100 V for 30 min and continued by ethidium bromide staining and visualized in UV-illuminator. The size was calculated by using λ-Eco T14I DNA marker (Takara).
Amplification of 16S rRNA gene by Polymerase Chain Reaction (PCR): The PCR was carried out to amplify about 1500 bp fragment of the 16S rRNA gene by using primers which designed based on the conserved region of 16S rRNA gene for bacteria. A combination of forward primers 16S forward (5-AGAGTTTGATCCTGGCTCAG-3) and 16S reverse (5-GGYTACCTTGTTACGACTT-3) was applied to amplify 16S rRNA sequence of bacteria. The PCR was performed using PCR thermal cycler (Applied Biosystem, USA). The total volume of 50 μL consisted of 35 μL dH2O (water freenuclease), 1 μL DNA template, 1.5 μL forward primer, 1.5 μL reverse primer and 5 μL 10x polymerase buffer, 1 μL taq polymerase (Takara, Japan) and 5 μL dNTP. The PCR condition was set as follows: Pre-denaturation at 94°C for 2 min, denaturation was performed at 94°C for 15 sec, continued by annealing at 50°C for 30 sec, elongation 68°C for 2 min (35 cycles) and post-elongation 68°C for 5 min. Amplified bands were resolved by electrophoresis in a 0.75% (w/v) agarose gel in TAE buffer and continued by ethidium bromide staining visualized in UV-illuminator. The size was calculated by using λ-Eco T14I DNA marker (Takara, Japan).
DNA sequencing and phylogenetic analysis: The purified DNA of selected isolates was sequenced using DNA analyzer (Applied Biosystems). Sequences were aligned and assembled using the sequencher 4.7 program (GENE CODES). The obtained sequences of the strain were used to search high similarity sequences deposited in the DNA database and the identities of isolates were calculated by the highest score (>98%). The phylogenetic tree was constructed with a neighbor-joining algorithm method which performed by using Clustal W and viewed using FigTree v1.4.0.
Scanning Electron Micrograph (SEM): Strain morphology observation was conducted by using SEM and research was initially by making the 1/100 diluted nutrient broth containing 3.5% agar and 0.1% gelatin. Glass fiber filter (GA100, ADVANTEC) was then prepared by cutting 5 mm2 and sterilized. The sterilized glass fiber was put on the 1/100 diluted nutrient agar medium and a small portion of the sterilized agar medium was dropped gently on the glass filter to make athin layer and solidified. Cells precultured in the nutrient broth were poured on the glass fiber. Then the glass fiber was incubated at 30°C for 24-48 h. Next, the cells on the glass filter were fixed with fixation solution that has consisted of 630 μL of 0.2 M HCl, 1.008 g of NaCl, 0.214 g of sodium cacodylate, 1 g of glutaraldehyde and filled up to 100 mL with distilled water. The glass filter was put into eppendorf tube (1.5 mL) and filled gently with 1 mL of fixation solution and stored in a refrigerator (4°C) for 30 min. Fixation solution was removed and then 60% acetone was poured into the same tube to remove water in the sample. The tube was then kept at room temperature for 15 min. The solution was removed and replaced by 80% acetone and further changed by 100% acetone. The treatment by 100% acetone was repeated twice. After removal of 100% acetone, 1 mL N-amyl acetate (Pentyl acetate) was added gently and stored at room temperature for 20 min. After dehydrating, the sample was vacuumed. After drying, the sample was coated with platinum using the JEOL JEC-3000FC auto fine coater. The sample was observed by SEM (JEOL JSM-6510LA).
Assessment of growth and ammonium reduction: Shaking culture experiments were conducted to observe the effect of (NH4)2SO4 addition at a different concentration of 0, 2.5, 5, 7 and 10% of total medium. Ammonium was mixed with auto claved sterile 1/100 meat extract medium to 100 mL in total volume. One percent of overnight strain preculture were then inoculated to medium in 250 mL erlenmeyer flasks and incubated at 30°C. The growth of the isolate was observed spectrophotometry at a wavelength of 600 nm (Shimadzu UV-1601PC spectrophotometer). Furthermore, the ability of the strain in reduce ammonia was performed by centrifuged culture samples at 8000 rpm for 15 min (Eppendorf centrifuge 5804R). Ammonia concentration was determined using nessler assay at a wavelength 425 nm.
RESULTS AND DISCUSSION
Isolation and selection of microorganisms: About 100 strains were isolated from soils from two laying hens industry that categorizes as high ammonia emission region and strain LS3K belong to that group. Because environmental conditions are usually nutritionally inadequate, a 1/100 nutrient agar was used for screening heterotrophic ammonium-responsive strains. We have isolated a soil bacterium, which located at high ammonia odorous region of laying hens farm in Yogyakarta city, Indonesia as tropical area. The isolated strain was responsive to the addition of 500 mg L–1 (NH4)2SO4 as the addition of nitrogen source in nutrient agar, from soil at Yogyakarta city.
The growth of strain LS3K at 1/00 nutrient broth was promoted by the addition of 500 mg L–1 in culture medium at 120 rpm shaker at 30°C after incubation at 4 days. Colony diameter of the strain was also confirmed increased in size when growing in 1/100 nutrient agar.
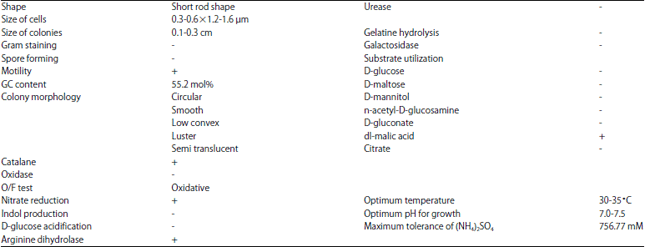
Morphological and physiological characteristics: The morphology of strain LS3K colonies on a nutrient agar plate was light yellow, circular, low convex, smooth and semi-translucent (Fig. 1). Spores are not formed. Morphological observation using scanning electron microscope (JEOL JSM-6510LA) of pure strains was in Fig. 2. It is found that cells of LS3K were a short rod, 0.3-0.6×1.2-1.6 μm in size. The morphological, biochemical and physiological characteristics of strain LS3K were shown in Table 1. The cells of the strain are Gram-negative bacteria, confirmed as oxidase negative, nitrate reduction and non-indole production.
| Table 1: | Morphological, biochemical and physiological characteristics of Pseudomonas sp., LS3K |
 | |
 | |
| Fig. 1: | Colony of strain LS3K growing on 1/100 meat extract medium contain 0.01% yeast extract, 0.01% polypepton and 0.005% NaCl with 1.5% agar. Medium was supplemented with 500 mg L–1 (NH4)2SO4 |
 | |
| Fig. 2: | Scanning electron micrograph of strain LS3K grown in 1/100 meat extract medium |
Furthermore, catalase reaction, starch hydrolysis and H2S production were positive, while citrate utilization was negative. Arginine dihydrolase was positive. These following substrates were not utilized for respiration: D-glucose, D-maltose and D-mannitol but dl-malic acid was used as a substrate for respiration. The n-acetyl-D-glucosamine, maltose and D-gluconate were also not used as a substrate by the strain. According to the data mentioned, strain LS3K was confirmed belong to genus Pseudomonas.
Phylogenetic analysis of 16S rRNA: The partial 16S rRNA sequence of strain LS3K was determined and the phylogenetic tree was constructed based on the 16S rRNA sequence (Fig. 3). The sequence was deposited in the GenBank database with accession No. LC034239. From comparative ribosomal DNA gene sequence as well as phylogenetic analysis, LS3K was deduced to be located in the genus Pseudomonas spp., close to Pseudomonas kuykendallii H2T (JF749828) and Pseudomonas tolaasii ND6-2T (AF348507) (Fig. 3). Some species close to the strain LS3K are Pseudomonas geniculata isolate OTU-a9T (KJ147059) and Pseudomonas kuykendallii H2T (JF749828) were also suggested take a role in organic wastes processing, that suppose they act some part in the composting reaction.
 | |
| Fig. 3: | Neighbour-joining phylogenetic analysis based on 16S rRNA gene sequences showing the positions of strains LS3KT among the type strains of selected Pseudomonas spp., available from GenBank NCBI/EMBL/DDBJ (accession numbers are given in parentheses). Bar, 0.5 substitutions per nucleotide position |
By examining physiological and biochemical characteristics and comparing the 16S rRNA gene sequence, the strain was identified as a strain of Pseudomonas and designed as Pseudomona sp., LS3K.
Growth and ammonia removal by strain LS3K: To take the role as ammonia mitigation agent, Pseudomona sp., LS3K should have the ability to grow in manure that contains a high concentration of animal manure and urine in the environment. Thus, research regarding the high concentration effect of (NH4)2SO4 and cow’s urine on the growth of strain LS3K was observed in 1/100 meat extract medium at 120 rpm aerobic shaking condition. Figure 4 shows the addition of (NH4)2SO4 at different concentration gave different growth profiles of strain LS3K. The growth of the strain in liquid medium with 0, 2.5 and 5%, ammonium addition was slightly lower comparing with control (without (NH4)2SO4). Furthermore, the addition of 7 and 10% of ammonium in medium gave impact on the delayed growth of strain LS3K as well as prolongation of adaptation phase until 18 h. Different phenomena were observed when strain LS3K is growing with various concentration of animal manure. Chicken manure which collected from one of poultry industry at Yogyakarta city gave a significant effect on the growth profiles of strain LS3K comparing with control, observed spectrophotometry until 48 h of cultivation period. High concentration of animal’s manure showed increasing force on the growth of strain LS3K.
Ammonium removal ability of strain LS3K in 1/100 meat extract culture medium was investigated in shaking condition at 30°C and the patterns of ammonium removal were shown in Fig. 5. Strain LS3K could oxidize 5.648 mg L–1 ammonium after 30 h cultivation time.
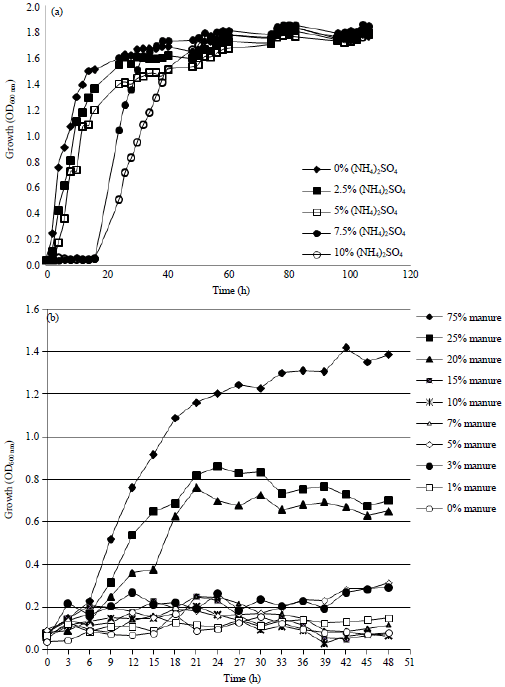 | |
| Fig. 4(a-b): | Growth of strain LS3K on (a) Ammonium and (b) Manure supplemented medium. Strain LS3K was cultivated in 1/100 dilution meat extract medium added by a various concentration of ammonium and animal’s manure |
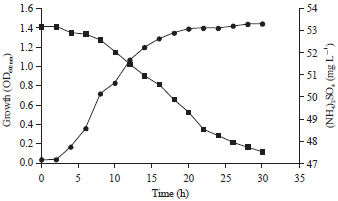 | |
| Fig. 5: | Growth curve of strain LS3K in the presence of 7% (NH4)2SO4 (black circle) and reduction of (NH4)2SO4 (black square) during cultivation period |
Biological treatment processes for ammonia emission mitigation from animal production are either based on the as similation and immobilization of volatile N or the transformation of volatile nitrogen into non-volatile inorganic nitrogen as fertilizer9. Transformation of volatile N to non-volatile is an important biological treatment process comprising of coupled nitrification and denitrification processes, which occur in the various environment. As a tropical country, Indonesia has continuously warm conditions that supposed to be a potential habitat for nitrifying-denitrifying bacteria. Especially in the high concentration of odorous ammonia area, we have tried to find microbes that adapted to this kind of condition. Furthermore, we had isolated soil bacterium from the odorous region of layer hens farm at Yogyakarta city and based on biochemical and physiological observation as well as gene analysis of 16S rRNA; the strain was identified as Pseudomona sp., LS3K, which can grow in high concentration ammonium medium. This is interesting due to the poison character of ammonium in a high dose to some microbes; however, strain LS3K have the ability to encounter that stressor. This strain was expected to be involved most treatments with some variation of physical, chemical or components of both physical and chemical unit processes to provide suitable conditions for the processes to occur efficiently and cost-effectively in mitigation of ammonia.
We suggest that strain LS3K has involved to the nitrification-denitrification in its habitat of the odorous region. During nitrification, this strain suggested transformed total ammonia calnitrogen to oxidize into N (nitrite and nitrate). These compounds are then biologically reduced to environmentally N gas (N2) by denitrifying bacteria as our next concern in performing research.
The reaction rate of nitrification of strain LS3K has observed in laboratory scale. Moreover, the nitrification rate of this strain to oxidize ammonium is low compared to that of an other strain such as Pseudomonas stutzeri YZN-00110 or Alcaligenes faecalis15. Strain LS3K could oxidize ammonium 5.648 mg L–1 during 30 h of cultivation period. As the acceleration for growing is quite low, we suggest that by prolonged the cultivation, the number of ammonium that can be oxidized by strain LS3K will increase. Nitrification is the more critical step and usually receives more attention for the biological treatment of waste waters for removal of NH3. The selected bacterium LS3K was highly tolerant of (NH4)2SO4, suggesting that strain LS3K is adaptable to animal wastes being composted or in animal’s manure.
On the other hands, ammonia (NH3) is a colorless gas, which is lighter than air and possesses a strong, penetrating odor in the environment. Gaseous NH3 is emitted from livestock production due to the excretion of nitrogen (N) from undigested protein as well as the amino acid from animal feed. Livestock manures which contain N in organic forms is potential as plant fertilizer as well as growth promoters. The excess of N in the feed and inefficient utilization of crude protein or amino acids in animal diets is the source of N in excreted urine and feces. Most of the N (up to 97%) is excreted as urea in the urine of ruminant and non-ruminant livestock such as sheep, cows and pigs; while the rest is excreted as undigested organic N in the feces6,23. Within hours to a few days, urea is then hydrolyzed to ![]() in a process catalyzed by the microbial enzyme urease originating mainly from feces9. In contrast, the microbial breakdown of organic N in feces into
in a process catalyzed by the microbial enzyme urease originating mainly from feces9. In contrast, the microbial breakdown of organic N in feces into ![]() in a process named as ammonification or mineralization requires months or even years to effect. The
in a process named as ammonification or mineralization requires months or even years to effect. The ![]() resulting from either urea hydrolysis or organic N decomposition or both is the one susceptible to volatilization from manure depending on pH and temperature conditions.
resulting from either urea hydrolysis or organic N decomposition or both is the one susceptible to volatilization from manure depending on pH and temperature conditions.
In laboratory scale study conducted, the optimum conditions for emission of odor from animal manure were observed at pH 8.6-10, temperature 35-40°C and moisture content 42-63%24. Treatments using microorganisms have been extensively used in municipal livestock waste for degradation of organic matter25. Furthermore, microbial treatments are emerging to treat livestock waste since degradation of organic matter in livestock waste relies on microorganisms26. On the other hand, using microorganisms as additive to encounter odor as well as ammonia emission from livestock production is become promising methods due to cheap and fast in action. Biological treatment processes for the ammonia mitigation from animal production are either based on the assimilation and immobilization of volatile N or the transformation of volatile N into non-volatile inorganic N as fertilizer9. The conversion of volatile N to non-volatile is a major biological treatment process comprising of coupled nitrification and denitrification processes that occur in the various environment. Nitrification is one of the key processes involved in the N cycle. In various ecosystems, ammonia-oxidizing bacteria oxidize NH3 to nitrite (NO2–), whereas NO2– is further oxidized to nitrate (NO3–) by nitrite-oxidizing bacteria. The major grouping of ammonia-oxidizing bacteria belongs to the subclass β-proteobacteria27.
The key in using bacterial cultures for deodorization of manure is to have the added bacteria become the predominant strain in the manure. For the added bacteria to the best growth, the real environment should appropriate from the optimum growth range for the bacteria. The indigenous flora (not only for those reducing odors) of the wastes always grew better than the inoculated microorganisms. In addition, the selected microorganisms may even use other organic compounds in preference to the malodorous substances when inoculated in wastes. Since predominance of the added bacteria is critical to the treatment, the quantity of bacterial material is questionable. Usually, the indigenous microorganisms are present in high concentration and are able to grow rapidly. Therefore, massive inoculation has to be exercised to accelerate the development of the added bacteria. Such massive inoculation can be achieved only on a laboratory scale, not at the farm level where the number of manure to be treated are considerable. According to Liao and Bundy28, microbial digestive additives contain bacteria or enzymes that eliminate odors and suppress gaseous pollutants by their biochemical digestive processes. Bourque et al.29 conducted research on microbiological degradation of odorous substances of swine manure on a laboratory scale under aerobic conditions29. The bacterial culture under study was inoculated into sterilized animal manure and incubated for a maximum of 6 days at 29°C. It is found that three bacterial species (Acinetobacter calcoaceticus, Alcaligenes faecalis and Arthrobacter flavescens) could completely degrade all types of VFAs in swine manure while Corynebacterium glutamicum and Micrococcus spp., could only degrade acetic and propionic acids. Another laboratory experiment has reported that Acinetobacter calcoaceticus could degrade VFAs in both sterilized and non-sterilized animal slurry incubated at 22°C within pH 6.2-8.6 for 21 days30.
Since the source of the odorous compounds is mainly microbial in origin, a sustained, rational research initiative is required using well-developed classical anaerobic microbiology technology, combined with modern molecular techniques and the latest analytical/sensory methodology, to determine the fundamentals controlling the production of malodor31.
This strain was expected to be involved most treatments with several variations of physical, chemical or components of both physical and chemical unit process in providing suitable conditions for the processes to occur efficiently and cost-effectively in mitigation of ammonia. We suggest that during nitrification, this strain transformed total ammoniacal nitrogen to oxidize into N (nitrite and nitrate). These compounds are then biologically reduced to environmentally benign N gas (N2) by denitrifying bacteria as our next concern in performing research.
CONCLUSION
The indigenous ammonia-oxidizing bacterium has successfully isolated from biological deodorization plants at poultry farm Yogyakarta Indonesia as tropicalarea and it was identified as Pseudomonas sp., LS3T. The strain could growth both in ammonium high contain medium and animals urine medium, with efficiently remove ammonium during the nitrification process. It indicates this strain could serve as microbial manure deodorization additive.
ACKNOWLEDGMENTS
This study was financially supported by grant aid from Directorate General of Higher Education (DGHE), Ministry of Education and Culture, the Republic of Indonesia through The Program Penelitian Unggulan UGM 2013 under LPPM management with grant number LPPM-UGM/370/LF/2013.
REFERENCES
- Chadwick, D., S. Sommer, R. Thorman, D. Fangueiro, L. Cardenas, B. Amon and T. Misselbrook, 2011. Manure management: Implications for greenhouse gas emissions. Anim. Feed Sci. Technol., 166: 514-531.
CrossRefDirect Link - Posmanik, R., A. Gross and A. Nejidat, 2014. Effect of high ammonia loads emitted from poultry-manure digestion on nitrification activity and nitrifier-community structure in a compost biofilter. Ecol. Eng., 62: 140-147.
CrossRefDirect Link - Elwinger, K. and L. Svensson, 1996. Effect of dietary protein content, litter and drinker type on ammonia emission from broiler houses. J. Agric. Eng. Res., 64: 197-208.
CrossRefDirect Link - Xie, K., X. Jia, P. Xu, X. Huang and W. Gu et al., 2012. Improved composting of poultry feces via supplementation with ammonia oxidizing archaea. Bioresour. Technol., 120: 70-77.
CrossRefDirect Link - Adrizal, P.H. Patterson, R.M. Hulet, R.M. Bates and D.A. Despot et al., 2008. The potential for plants to trap emissions from farms with laying hens: 2. Ammonia and dust. J. Applied Poultry Res., 17: 398-411.
CrossRefDirect Link - McCrory, D.F. and P.J. Hobbs, 2001. Additives to reduce ammonia and odor emissions from livestock wastes: A review. J. Environ. Qual., 30: 345-355.
PubMedDirect Link - Satoh, K., R. Takizawa, M. Sarai, N. Sato, R. Takahashi and T. Tokuyama, 2004. Two kinds of ammonia-oxidizing bacteria isolated from biologically deodorizing plants in cold district. J. Biosci. Bioeng., 98: 207-210.
CrossRefDirect Link - Zeng, Y., A. De Guardia, C. Ziebal, F.J. De Macedo and P. Dabert, 2012. Nitrification and microbiological evolution during aerobic treatment of municipal solid wastes. Bioresour. Technol., 110: 144-152.
CrossRefDirect Link - Ndegwa, P.M., A.N. Hristov, J. Arogo and R.E. Sheffield, 2008. A review of ammonia emission mitigation techniques for concentrated animal feeding operations. Biosyst. Eng., 100: 453-469.
CrossRefDirect Link - Zhang, J., P. Wu, B. Hao and Z. Yu, 2011. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001. Bioresour. Technol., 102: 9866-9869.
CrossRefDirect Link - Zeng, G., J. Zhang, Y. Chen, Z. Yu and M. Yu et al., 2011. Relative contributions of archaea and bacteria to microbial ammonia oxidation differ under different conditions during agricultural waste composting. Bioresour. Technol., 102: 9026-9032.
CrossRefDirect Link - Ma, Y., L. Wang and L. Qian, 2008. Community structure of β-Proteobacterial ammonia-oxidizing bacteria in prawn farm sediment. Progr. Nat. Sci., 18: 679-684.
CrossRefDirect Link - Sauder, L.A., K. Engel, J.C. Stearns, A.P. Masella, R. Pawliszyn and J.D. Neufeld, 2011. Aquarium nitrification revisited: Thaumarchaeota are the dominant ammonia oxidizers in freshwater aquarium biofilters. PLoS One, Vol. 6.
CrossRef - Zhang, Q.L., Y. Liu, G.M. Ai, L.L. Miao, H.Y. Zheng and Z.P. Liu, 2012. The characteristics of a novel heterotrophic nitrification-aerobic denitrification bacterium, Bacillus methylotrophicus strain L7. Bioresour. Technol., 108: 35-44.
CrossRefDirect Link - Joo, H.S., M. Hirai and M. Shoda, 2005. Nitrification and denitrification in high-strength ammonium by Alcaligenes faecalis. Biotechnol. Lett., 27: 773-778.
CrossRefDirect Link - Hooijmans, C.M., S.G.M. Geraats, E.W.J. van Niel, L.A. Robertson, J.J. Heijnen and K.C.A.M. Luyben, 1990. Determination of growth and coupled nitrification/denitrification by immobilized Thiosphaera pantotropha using measurement and modeling of oxygen profiles. Biotechnol. Bioeng., 36: 931-939.
CrossRefDirect Link - Zhao, B., Q. An, Y.L. He and J.S. Guo, 2012. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresour. Technol., 116: 379-385.
CrossRefDirect Link - Selvam, A., Z. Zhao, Y. Li, Y. Chen, K. Leung and J. Wong, 2013. Degradation of tetracycline and sulfadiazine during continuous thermophilic composting of pig manure and sawdust. Environ. Technol., 34: 2433-2441.
CrossRefDirect Link - Sonoki, T., T. Furukawa, K. Jindo, K. Suto, M. Aoyama and M.A. Sanchez‐Monedero, 2013. Influence of biochar addition on methane metabolism during thermophilic phase of composting. J. Basic Microbiol., 53: 617-621.
CrossRefDirect Link - Lin, S.Y., Y.C. Liu, A. Hameed, Y.H. Hsu, W.A. Lai, F.T. Shen and C.C. Young, 2013. Azospirillum fermentarium sp. nov., a nitrogen-fixing species isolated from a fermenter. Int. J. Syst. Evolutionary Microbiol., 63: 3762-3768.
CrossRefDirect Link - Mackie, R.I., O.G. Stroot and V.H. Varel, 1998. Biochemical identification and biological origin of key odor components in livestock waste. J. Anim. Sci., 76: 1331-1342.
CrossRefDirect Link - Ohta, Y. and M. Ikeda, 1978. Deodorization of pig feces by actinomycetes. Appl. Environ. Microbiol., 36: 487-491.
PubMedDirect Link - Low, E.W. and H.A. Chase, 1999. Reducing production of excess biomass during wastewater treatment. Water Res., 33: 1119-1132.
CrossRefDirect Link - Sund, J.L., C.J. Evenson, K.A. Strevett, R.W. Nairn, D. Athay and E. Trawinski, 2001. Nutrient conversions by photosynthetic bacteria in a concentrated animal feeding operation lagoon system. J. Environ. Quality, 30: 648-655.
CrossRefDirect Link - Spieck, E., E. Bock and H.P. Koops, 2005. Nitrifying Bacteria, the Lithoautotrophic Ammonia-Oxidizing Bacteria, the Lithoautotrophic Nitriteoxidizing Bacteria in Part A. In: Bergey's Manual of Systematic Bacteriology, Brenner, D.J., N.R. Krieg, J.T. Staley and G.M. Garrity (Eds.). Vol. 2, 2nd Edn., Springer, New York.
- Liao, C.M. and D.S. Bundy, 1994. Bacteria additives to the changes in gaseous mass transfer from stored swine manure. J. Environ. Sci. Health Part B: Pestic. Food Contamin. Agric. Wastes, 29: 1219-1249.
CrossRefDirect Link - Bourque, D., J.G. Bisaillon, R. Beaudet, M. Sylvestre, M. Ishaque and A. Morin, 1987. Microbiological degradation of malodorous substances of swine waste under aerobic conditions. Appl. Environ. Microbiol., 53: 137-141.
Direct Link - Jolicoeur, P. and A. Morin, 1987. Isolation of Acinetobacter calcoaceticus strains degrading the volatile fatty acids of swine wastes. Biol. Wastes, 19: 133-140.
CrossRefDirect Link - Zhu, J., 2000. A review of microbiology in swine manure odor control. Agric. Ecosyst. Environ., 78: 93-106.
CrossRefDirect Link