
Hari Marta Saputra
Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Sayati Mandia
Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Bambang Retnoaji
Laboratory of Animal Structure and Development, Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Nastiti Wijayanti
Laboratory of Animal Physiology, Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 3 | Page No.: 77-85







ABSTRACT
Lead (Pb2+) is a toxic heavy metal causes oxidative stress that affect vital organs in the body, such as kidneys. Lead (Pb2+) induces formation of free radicals on cells, generation of Reactive Oxygen Species (ROS), Reactive Nitrogen Species (RNS) and decreases level of endogenous antioxidant defense mechanism. Oxidative stress triggers synthesis of Heat Shock Protein (HSP) as sublethal response. The imbalance level between oxidant and antioxidant defense mechanisms has been suggested to play an important role in cells death. Administration of exogenous antioxidant from plant, such as liverwort (Marchantia polymorpha L.) assumed can decrease free radical levels caused by heavy metal. The objective of this study was to investigate antioxidant activity of liverwort and to investigate whether methanol extract of M. polymorpha was affect HSP70 expression and NO production in base-line levels on HEK293 (human embryonic kidney) cell line. The DPPH was used to determine antioxidant activity, Griess assay to quantify nitric oxide and Western blotting was used to detect HSP70 expression. The result showed that the activity of antioxidant scavenger of liverwort is about 5.22 μg mL–1 of IC50 value. In addition, liverwort extract can increase cell viability and decrease NO production significantly between Pb(NO3)2 and treatment group and also decrease HSP70 expression attained to base-line level.
PDF Abstract XML References Citation
Received: January 12, 2016;
Accepted: February 16, 2016;
Published: March 15, 2016
How to cite this article
Hari Marta Saputra, Sayati Mandia, Bambang Retnoaji and Nastiti Wijayanti, 2016. Antioxidant Properties of Liverwort (Marchantia polymorpha L.) to Lead-induced Oxidative Stress on HEK293 Cells. Journal of Biological Sciences, 16: 77-85.
DOI: 10.3923/jbs.2016.77.85
URL: https://scialert.net/abstract/?doi=jbs.2016.77.85
DOI: 10.3923/jbs.2016.77.85
URL: https://scialert.net/abstract/?doi=jbs.2016.77.85
INTRODUCTION
Lead contamination in the environment is found from cigarette smoke, fumes, food or objects that contain lead (Suciani, 2007; Ashraf, 2011; Ariansya et al., 2012). Administration of lead into the body through the respiratory tract (inhalation) (Irwan et al., 2013) and gastrointestinal tract (Faix et al., 2005). Lead as a toxicant has been widely used to in vivo and in vitro experimental (Aykin-Burns et al., 2005; Casado et al., 2007; Jankeer and El-Nouri, 2009; Mehana et al., 2012; Missoun et al., 2010; Gurer et al., 2001; Ibrahim et al., 2012). Lead is toxicant causes damage to the brain, heart, liver, kidneys, gastrointestinal tract, respiratory tract and changes in blood profile in humans or animals model as well as rat and mice (Mugahi et al., 2003; Jankowska-Kulawy et al., 2008; Reza et al., 2009; Mehana et al., 2012; Missoun et al., 2010; Aziz et al., 2012).
Lead in the cell will trigger excessive formation of free radicals because of its ability to bind to the enzymes, which are forming natural antioxidants. The imbalance of oxidants and natural antioxidants in cells will form oxidative stress through two mechanisms, namely the formation of Reactive Oxigen Species (ROS) and decrease the synthesis of endogenous antioxidants enzyme (antioxidant defense system) such as superoxide dismutase (SOD) and catalase (CAT) (Flora et al., 2012). At high concentrations, ROS can be important mediators of DNA, protein and lipid damage. Cells under oxidative stress would trigger the activation of Heat Shock Factor (HSF) and begin the process of Heat Shock Proteins (HSP) synthesis as a sublethal response of cells to environmental stress (Filipovic et al., 2008). Therefore, to maintain cells homeostasis against free radicals excess, it is need a way of balancing the oxidant and antioxidant cell by adding an exogenous antioxidant compound (Gurer et al., 2001) of plants that have antioxidant activity, such as flavonoids and phenolic. One of Indonesian plant as potential source of exogenous antioxidants is liverwort (Marchantia polymorpha L.). In contrast to China and India, study on the antioxidant activity of liverworts in Indonesia has not been done.
The main secondary metabolite compounds in M. polymorpha are flavonoid, terpenoid and phenol. The primarily flavonoids in Marchantiaceae are luteolin (Gokbulut et al., 2012), quercetin, apigenin, O-glycoside, C-glycosides (Xiao et al., 2006a), marchantin A and C (Friederich et al., 1999) as well as polyphenols (Krishnan and Murugan, 2013). Liverworts are used as anti-microbial, anti-fungal (Veljic et al., 2010; Khanam et al., 2011), anti-cancer (Xiao et al., 2005; Huang et al., 2010; Hegazy et al., 2012) and anti-hepatitis B (Xiao et al., 2006b). However, there has been no report about antioxidant activity of liverwort against oxidative stress caused by lead. The secondary metabolite of M. polymorpha allegedly can decreased excessive free radicals in cells. The objective of this study was to investigate the antioxidant scavenging activity of M. polymorpha extract, its effect to expression of Heat Shock Protein 70 (HSP70) and NO concentration after Pb(NO3)2-induced oxidative stress on HEK293 cells.
MATERIALS AND METHODS
Marchantia polymorpha was collected in Mt. Merapi area, Yogyakarta, Indonesia on September, 2014. Vitamin C was used as antioxidant control. Treatment were divided into 6 groups: Untreated as negative control, antioxidant control (Vitamin C), positive control (10 μg mL–1 of lead nitrate for 24 h) and treatment groups (10 μg mL–1 of lead nitrate for 24 h then added by marchantia extract with several doses: 10, 30, 60 and 90 μg mL–1 for 24 h to determine cell viability and NO production and 5, 10 and 30 μg mL–1 to determine HSP70 expression).
Plant material and extraction: Marchantia polymorpha were washed with running water. Thallus (leaves) were separated from rhizoid. Thallus allowed to air-dry in shade for a week. During the drying process, the thallus wrapped with paper. The plant material was subsequently homogenised in electrical blender. About 350 g of powdered sample was extracted until exhaustion by maceration with 3 L of methanol 80% for 2×24 h. The extracts were filtered and re-extracted in same way three to four times until the colour of extract seen transparent. The filtrate were combined and evaporated using a rotary evaporator. Crude extract dissolved in dimethyl sulphoxide (DMSO) (Huang et al., 2010).
Antioxidant scavenging assay: A total of 1 mL of each sample extracts with various concentrations (0, 5, 10, 15, 25, 30, 50, 60, 75, 90 and 100 μg mL–1) were diluted using methanol and added to cuvette. One milliliter of 0.1 mM DPPH (Himaja et al., 2010) was added to each cuvette. Samples were incubated for 30 min at room temperature and placed in a dark room and absorbance was measure at 517 nm. Ascorbic acid in various concentrations (2.5, 5, 7.5, 10, 12.5 and 15 μg mL–1) were used as a standard positive control and methanol as blank, DPPH solution as negative control. The results were calculated as percentage scavenging of DPPH• (Charoensin, 2014):
![]()
Cell culture: Human embryonic kidney cell line (HEK293) was cultured in Dulbaccos’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum, 100 U mL–1 penicillin, 100 μg mL–1 streptomycin and 1% fungizon. Cell cultures were kept under a 5% CO2 humidified atmosphere at 37°C. The cells were seeded in 96-well cell culture plates to measure cell viability and NO concentration. In other hand, cells were seeded in 6 cm (in diameter) culture disk to measure expression of HSP70.
Cell viability assay: Cell survival was accessed by cleavage of the tetrazolium salt WST-1 to formazan by cellular enzymes. An expansion in the number of viable cells results in an increase in the overall activity of mitochondrial dehydrogenases in the sample. Quantification of the formazan dye produce by metabolically active cells by scanning multiwell spectrophotometer. Briefly, 1×103 cells in 100 μL of media to each well were seeded into 96-well cell culture plate and incubated for 72 h. The medium was then replaced with 100 mL fresh medium contained 10 μg mL–1 Pb(NO3)2 and incubation for 24 h. Marchantia extract was added in the next day appropriate to its concentrations for 24 h. Then after, the medium was replaced with 100 μL fresh medium and 10 μL cell proliferation reagent WST-1 (4-[3-(Iodophenyl) -2-(4-nitrophenyl)-2H-5-tetrazolio]-1,3-benzene disulfonate) (Roche). Incubate the cells for 1 h in a humidified atmosphere. Shake thoroughly for 1 min on a shaker. Measure the absorbance of samples againts a background control as blank using microplate (ELISA) reader at 450 nm. Cell viability is calculated by the formula:
![]()
Nitric oxide assay: The sulfanilamide solution and NED solution were equilibrated to room temperature (15-30 min). Add 50 μL of each sample (medium HEK293 cells that had been treated) to 96-well microplate in triplicate. Dispense 50 μL sulfanilamide solution to all samples and wells containing the dilution series for the nitrite standard reference curve. Incubate 5-10 min at room temperature, protected from light. Dispense 50 μL of NED solution to all wells, incubate at room temperature and protected from light. A purple/magenta color will begin to form immediately. Measure absorbance within 30 min in a plate reader in 595 nm. The NO concentrations are determined by nitrite standard curve.
Western blotting: Harvested cells were homogenized with lysis buffer (tris-HCl 0,0005 M, pH 7.1, 2% SDS, 10% glycerol, 0.2 M DTT, a punch of bromphenol blue, 4% protese inhibitor), followed by sonication and sentrifuged at 13000 rpm for 15 min. The protein concetration was measured by the Bradford method (Bio-Rad Protein Assay; Bio-Rad Laboratories Inc., Hercules, CA, USA). An equal amount of proteins (5 μg) were separated by 10% SDS-PAGE. Proteins were electrotransferred to an immobilion polyvinylidene difluoride (PVDF) membrane. This was followed by 1 h blocking with solution composed of 20 mM tris-HCl (pH 7.6), 125 mM HCl, 0.2% tween 20, aquadest and 5% skim milk. Specific antibodies used were anti-HSP70 (1:1000 of a rabbit polyclonal antibody) and anti-α-tubulin (1:1000 of a mouse monoclonal antibody). Detection was made using HRP conjugated mouse anti-rabbit and goat anti-mouse IgG (1:10000) and chemiluminescence/ECL (Amersham, GE Healthcare, Buckinghamshire, UK). Densitometric analysis of bands was performed using ImageJ software.
Statistical analysis: Statistically analysis was performed using one-way ANOVA and Duncan for further analysis (p<0.05). Statistical data analysis was performed with SPSS software ver. 20. Densitometric of HSP70 bands was analysis using description method.
RESULTS
Antioxidant scavanging activity of M. polymorpha extract: The antioxidant scavenging activity of M. polymorpha extract and vitamin C to radical DPPH were exhibited in Table 1 and 2. The IC50 value is found to be 5.22 and 0.34 μg mL–1, respectively for M. polymorpha extract and vitamin C.
| Table 1: | Antioxidant activity of Marchantia extract |
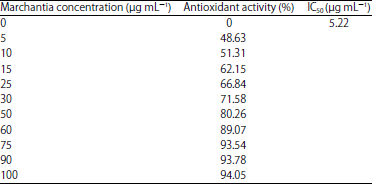 | |
| Values are the mean of three observations | |
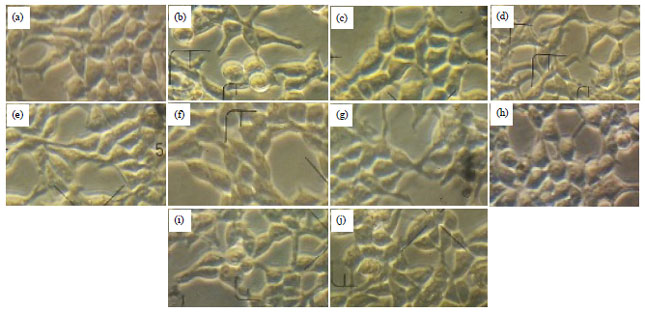 | |
| Fig. 1(a-j): | Morphology of HEK293 cells after treatments with Pb(NO3)2, various concentration of Marchantia and vitamin C for 24 h, (a) Control, (b) Pb(NO3)2 10 μg mL–1, (c) Vitamin C 10 μg mL–1, (d) Pb(NO3)2 10 μg mL–1+vitamin C 10 μg mL–1, (e) Marchantia 5 μg mL–1, (f) Pb(NO3)2 10 μg mL–1+Marchantia 5 μg mL–1, (g) Marchantia 10 μg mL–1, (h) Pb(NO3)210 μg mL–1+Marchantia 10 μg mL–1, (i) Marchantia 30 μg mL–1 and (j) Pb(NO3)2 10 μg mL–1+Marchantia 30 μg mL–1 (magnification, 400x) |
| Table 2: | Antioxidant activity of vitamin C |
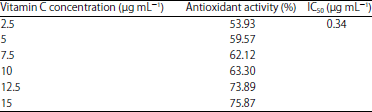 | |
| Values are the mean of three observations | |
| Table 3: | Viabilities of HEK293 cells in the treatment of lead and Marchantia |
 | |
| Values are the Mean±SD of three observations (p<0.05 compared to the percentage viability of the control group) | |
| Table 4: | Viabilities of HEK293 cell in the treatment of Marchantia |
 | |
| Values are the Mean±SD of three observations (p<0.05 compared to the percentage viability of the control group) | |
Morphology of HEK293: Figure 1 shows the morphology of HEK293 cells changed after treated with lead. The morphological changes were observed under microscope (magnification, 400x) in lead group neither in other groups compared to the control group.
HEK293 cell viability: Marchantia polymorpha extract known have been flavonoid, terpenoid and phenolic compounds which are implicated to free radical levels, the altered cell viability was found after treatment with Pb(NO3)2 and continued with various concentration of M. polymorpha extract are increased cell viability in HEK293 (Fig. 2). In addition, there is significant increase in cell viability (treatment lead+M. polymorpha 10 μg mL–1, lead+M. polymorpha 30 μg mL–1 and lead+M. polymorpha 60 μg mL–1) compared to control and lead group (Table 3). By contrast, HEK293 (treatment with various concentration of M. polymorpha extract alone) showed a dramatically reduced rate of proliferation and increased rate of cell death (Table 4).
Nitric oxide concentration (NO) HEK293 cells: The NO concentration in sample medium of HEK293 cell was assesed. The increased of NO concentration in lead group (15.53 μM) related to decreased of cell viability (59.13%) (Fig. 3).
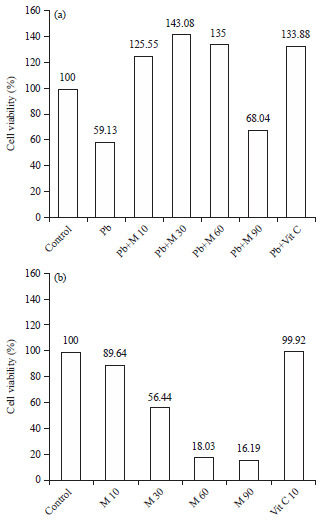 | |
| Fig. 2(a-b): | Viability of HEK293 cells after treatment, (a) Pb (NO3)2, Marchantia extract and vitamin C and (b) Marchantia extract and vitamin C |
| Table 5: | NO concentration of HEK293 in the treatment of lead and Marchantia |
 | |
| Values are the Mean±SD of three observations (p<0.05 compared to the percentage concentration of NO control group) | |
| Table 6: | NO concentration of HEK293 in the treatment of Marchantia |
 | |
| Values are the Mean±SD of three observations (p<0.05 compared to the percentage concentration of NO control group) | |
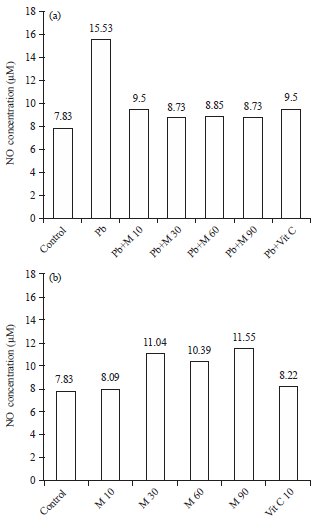 | |
| Fig. 3(a-b): | NO levels HEK293 cells after treatment (a) Pb(NO3)2, Marchantia extract and vitamin C and (b) Treatment Marchantia extract and vitamin C |
There is significant effect of M. polymorpha extract due to NO concentration after administration of lead continued with M. polymorpha extract (Table 5) compared to lead group (NO concentration and cell viability, 7.83 μM and 100%, respectively). Further, there is no significant effect in treatment groups compared to control group. In M. polymorpha alone, there is increased NO concentration in all treatments compared to control group (Table 6).
HSP70 expression: To further investigate the oxidative stress caused by lead, which is induced transcription of hsp genes, it is performed Western blot to detected expression of HSP70 in HEK293 cells. There was dramatically over-expression of HSP70 in lead group and reduced in treatment groups. In addition, HSP70 expression is higher in M. polymorpha extract alone than control group (Fig. 4 and 5).
 | |
| Fig. 4: | Expression of Heat Shock Protein 70 (HSP70) HEK293 cells after treatment Pb (NO3)2, Marchantia extract and vitamin C |
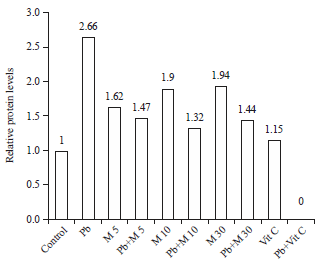 | |
| Fig. 5: | Expression of Heat Shock Protein 70 (HSP70) HEK293 cells after treatment Pb(NO3)2, Marchantia extract and vitamin C |
DISCUSSION
Measurement of antioxidant scavenging activity of M. polymorpha extract occured by reacting DPPH-methanol solution with M. polymorpha extract. Absorbance of DPPH-Marchantia extract solution was measure at 517 nm. Further, IC50 value obtained from linear regression.
The results showed IC50 value of M. polymorpha extract to be 5.22 μg mL–1 while, the IC50 value of vitamin C to be 0.344 μg mL–1. Gokbulut et al. (2012) collected samples of M. polymorpha in the province of Anatolia, Turkey, get the IC50 value of M. polymorpha extract with methanol solvent to be 0.4495±0.029 μg mL–1 and ethyl acetate solvent to be 0.2756±0.001 μg mL–1. The IC50 value of methanolic extract of M. polymorpha from Yogyakarta. Indonesia have a higher antioxidant activity than methanolic extracts of M. polymorpha from Turkey. Secondary metabolites Marchantia extract consisting of flavonoids, terpenoids and phenolic. The most flavonoids of Marchantia genus are luteolin (Gokbulut et al., 2012), quercetin, apigenin, O-glycoside, C-glycosides (Xiao et al., 2006a), marchantin A and C (Friederich et al., 1999) as well as polyphenols (Krishnan and Murugan, 2013). These compounds are assumed reduce free radicals or ROS in HEK293 cell induced by lead as well as taurine (an antioxidant) against lead-induced oxidative stress (Gurer et al., 2001).
Cell viability of HEK293 was decreased in lead group (59.13%) compared to control group (100%). On the other hand, cell viability was increased in treatment groups (125.55, 143.08 and 135%, respectively in concentration 10, 30 and 60 μg mL–1) (Table 3). Beside this, cell viabiliy was significantly decreased in Marchantia alone (Table 4). Increased cell viability was also found in cells exposed to Pb(NO3)2, followed by administration of vitamin C 10 μg mL–1 to be 133.88%. The contrary results, cell viability exposed to Pb (NO3)2 and Marchantia extract at concentration of 90 μg mL–1 was decreased compared to control and increased when compared to the treated with Pb(NO3)2 about 68.04%. Based on statistical test there was no significant difference between treatment group of Pb(NO3)2 and treatment group Pb(NO3)2+90 μg mL–1 extract Marchantia. The same results was found among treatment groups Pb(NO3)2+10 μg mL–1, Pb(NO3)2+30 μg mL–1 and Pb(NO3)2+60 μg mL–1 extract Marchantia.
Lead has several effect cause dysfunction of physiology and biochemistry in animals and humans. Lead induced cell damage through lipid peroxidation, protein damage and DNA damage leading to cell death also known as apoptosis. The Pb2+ entry into the cell by utilizing the protein binding of metal ions such as Ca2+ and Zn2+, which is flexible and able to bind to the calcium and zinc binding-site. The Pb2+ has the ability to replace Ca2+, Zn2+ and other divalent cations in cellular systems that have an effect on changing the environment inside the cell. The Pb2+ will be accumulated in some cell organelles, such as mitochondria, endoplasmic reticulum and the nucleus. The Pb2+ in the mitochondrial inner membrane will cause damage, degradation of the energy metabolism and forming generation of free radical. Moreover, Pb2+ inhibits income (uptake) Ca2+ from the cytoplasm into the mitochondria. The opening of the pore-permeability transition induce cytochrome C release into the cytoplasm, which is trigger apoptosis by activating caspase-9 and caspase-3 (Yedjou et al., 2010).
Administration of Pb can decreased cell viability in nerve cell culture (PC-12) and increased after given N-acetylcysteine (NAC) as an antioxidant agent (Aykin-Burns et al., 2005). In this study, cell viability of HEK293 decreased was found after exposure to Pb(NO3)2 and increased after given Marchantia extract. Marchantia extract has several active compound that play an important role for the regulation of excess free radicals cause oxidative stress on HEK293 cells after exposure to Pb(NO3). Active compounds of M. polymorpha extract from Yogyakarta, Indonesia known as flavonoids, phenolics and terpenoids as a TLC analysis.
Antioxidant compounds have capability to stabilizing free radicals formed due to induction of Pb(NO3)2, which causes oxidative stress that leads to apoptosis. Flavonoids, phenolics and terpenoids in M. polymorpha extract can reduce free radicals in HEK293 cells by stabilizing the atoms bonding, which do not have a paired electrons in its outer orbital. On the other hand, Marchantia extract alone in HEK293 cells also decreased HEK293 cell viability. Terpenoid in the extract allegedly played an important role against cell apoptosis. Terpenoids compounds can induce cell apoptosis (Lu et al., 2012) through the activation of p53 (Liu et al., 2012).
Nitric oxide concentrations of HEK293 cells after treated with Pb(NO3)2 for 24 h followed by several concentrations of Marchantia and vitamin C for 24 h are presented in Table 5 and 6. Nitric oxide concentration in Pb(NO3)2 group is 15.52 μM significantly than the control group 7.83 μM. This result indicates HEK293 cells are in nitrosative stress (a high concentration of NO inside cell) and decreased after treated with Marchantia extract. The data showed, NO concentration befall to base-line level (toward to control group). There is no significant difference in NO concentration between the treatment groups compared to control group but significantly different when compared to lead group. This indicates that Marchantia extract can reduce NO concentration in HEK293 cells were exposed to Pb(NO3)2. In Marchantia extract alone (30, 60 and 90 μg mL–1), NO concentrations were increased compared to control group in the amount of 11.04, 10.39 and 11.55 μM, respectively. Nitric oxide concentration in group Marchantia alone (10 μg mL–1) and vitamin C (10 μg mL–1) presents the concentration of NO to be 8.09 and 8.22 μM. Nitric Oxide (NO) is a free radical made from Reactive Nitrogen Species group. The NO• is a small molecule that has an unpaired electron in the orbital, therefore, NO• also called free radicals. The NO• is a product of Nitric Oxide Synthases (NOS), which acts on the metabolism of arginine into citrulline with by product through electron oxidative reactions. In normal physiological processes, NO• have function as neurotransmitters, blood pressure regulation, defense mechanisms, regulation of immunity and smooth muscle relaxation. Administration of Pb2+ can increase the production of NO• (Matovic et al., 2015). Excess of NO• production is known as nitrosative stress. The NO• can bind to O•- forming reactive peroxynitrite anion/peroxynitrite anion (ONOO-), which is very reactive and cause DNA fragmentation and lipid oxidation (Valko et al., 2007).
One of the stress protein is Heat Shock Protein 70 (HSP70). To investigate whether HSP70 was expressed, it is determined the HSP70 protein levels in HEK293 using Western blot analysis. Figure 4 and 5 shows that Pb2+ caused significant induction of HSP70 protein levels, in a manner related to what investigated at morphology changes, cell viability and NO production. There is significant of HSP70 protein levels between lead group compared to control group. The HSP70 were translated more reducing in treatment groups (approach to base-line levels). Surprisingly, in Marchantia alone HSP70 were translated more higher, which are related to increased cell death and NO production. Unfortunately, it did not found HSP70 in Pb(NO3)2+vitamin C, although treated with vitamin C shows no differences to control group.
The HSP70 is expressed more higher when the cell is in unbenefit condition due to environmental stress, such as the amount of Pb2+ in cells that can cause oxidative stress and trigger increased expression of protein stress (Forreiter and Nover, 1998; Ramp et al., 2007; Mandrekar et al., 2008).
Stressful conditions cause unfolding, misfolding or aggregation of proteins that will trigger the stress response induce the transcription process of genes into proteins. Transcription of HSP gene mediated by Heat Shock Factor (HSF1) by Heat Shock Elements (HSEs) in the promoter region of the HSP gene. In normal conditions, HSF1 is located in the cytoplasm as latent monomeric molecules. Under stress conditions, HSF1 will be hiperphosphorilation by members of the subfamily MAPK (mitogen-activated protein kinase), such as ERK1, JNK/SAPK, p38 protein kinase. The HSF1 will be joined by two other HSF1 monomers to form a trimer that has ability to bind to DNA in the region HSES (Dubey et al., 2015). The HSP will be assisted of regulation in misfolding, unfolding and aggregation of proteins inside the cells and avoid apoptosis process by inhibiting caspases regulation (Jolly and Morimoto, 2000).
CONCLUSIONS
Lead induces formation of free radicals on cells, generation of Reactive Oxygen Species (ROS), Reactive Nitrogen Species (RNS) and caused oxidative stress in HEK293 cell. Lead decreased cell viability, increased NO production and trigger HSP70 protein expression more higher. Marchantia polymorpha L. can reduce formation of free radicals, ROS, RNS and oxidative stress. Further, it also impacte to increased cell viability, decreased NO production and HSP70 protein expression.
REFERENCES
- Ariansya, K.A., K. Yuliati and R.J. Siti Hanggita, 2012. Analisis kandungan logam berat (Pb, Hg, Cu dan As) pada kerupuk kemplang Di desa tebing gerinting utara, kecamatan indralaya selatan, kabupaten ogan ilir. Fishtech, 1: 69-77.
Direct Link - Ashraf, M.W., 2011. Concentrations of cadmium and lead in different cigarette brands and human exposure to these metals via smoking. J. Arts Sci. Commerce, 2: 140-147.
Direct Link - Aykin-Burns, N., E.A. Franklin and N. Ercal, 2005. Effects of N-acetylcysteine on lead-exposed PC-12 cells. Arch. Environ. Contam. Toxicol., 49: 119-123.
CrossRefDirect Link - Casado, M.F., A.L. Cecchini, A.N.C. Simao, R.D. Oliveira and R. Cecchini, 2007. Free radical-mediated pre-hemolytic injury in human red blood cells subjected to lead acetate as evaluated by chemiluminescence. Food Chem. Toxicol., 45: 945-952.
CrossRefDirect Link - Charoensin, S., 2014. Antioxidant and anticancer activities of Moringa oleifera leaves. J. Med. Plants Res., 8: 318-325.
CrossRefDirect Link - Dubey, A., K.S. Prajapati, M. Swamy and V. Pachauri, 2015. Heat shock proteins: A therapeutic target worth to consider. Vet. World, 8: 46-51.
Direct Link - Faix, S., Z. Faixova, K. Boldizarova and P. Javorsky, 2005. The effect of long-term high heavy metal intake on lipid peroxidation of gastrointestinal tissue in sheep. Veterinarni Medicina, 9: 401-405.
Direct Link - Filipovic, D., L. Gavrilovic, S. Dronjak, M. Demajo and M.B. Radojcic, 2008. Liver glucocorticoid receptor and heat shock protein 70 levels in rats exposed to different stress models. Physiol. Res., 57: 205-213.
PubMedDirect Link - Flora, G., D. Gupta and A. Tiwari, 2012. Toxicity of lead: A review with recent updates. Interdiscip. Toxicol., 5: 47-58.
CrossRefPubMedDirect Link - Forreiter, C. and L. Nover, 1998. Heat induced stress proteins and the concept of molecular chaperones. J. Biosci., 23: 287-302.
CrossRefDirect Link - Friederich, S., M. Rueffer, Y. Asakawa and M.H. Zenk, 1999. Cytochromes P-450 catalyze the formation of marchantins A and C in Marchantia polymorpha. Phytochemistry, 52: 1195-1202.
CrossRefDirect Link - Gokbulut, A., B. Satilmis, K. Batcioglu, B. Cetin and E. Sarer, 2012. Antioxidant activity and luteolin content of Marchantia polymorpha L. Turk. J. Biol., 36: 381-385.
CrossRefDirect Link - Gurer, H., H. Ozgunes, E. Saygin and N. Ercal, 2001. Antioxidant effect of taurine against lead-induced oxidative stress. Arch. Environ. Contam. Toxicol., 41: 397-402.
CrossRefPubMedDirect Link - Hegazy, M.E.F., A.M. Gamal-Eldeen, A.M. El-Halawany, A.E.H.H. Mohamed and P.W. Pare, 2012. Steroidal metabolites transformed by Marchantia polymorpha cultures block breast cancer estrogen biosynthesis. Cell Biochem. Biophys., 63: 85-96.
CrossRefDirect Link - Himaja, M., R. Anand, M.V. Ramana, M. Anand and A. Karigar, 2010. Phytochemical screening and antioxidant activity of rhizome part of Curcuma zedoaria. Int. J. Res. Ayurveda Pharmacy, 1: 414-417.
Direct Link - Huang, W.J., C.L. Wu, C.W. Lin, L.L. Chi and P.Y. Chen et al., 2010. Marchantin A, a cyclic bis(bibenzyl ether), isolated from the liverwort Marchantia emarginata subsp. tosana induces apoptosis in human MCF-7 breast cancer cells. Cancer Lett., 291: 108-119.
CrossRefDirect Link - Ibrahim, N.M., E.A. Eweis, H.S. El-Beltagi and Y.E. Abdel-Mobdy, 2012. Effect of lead acetate toxicity on experimental male albino rat. Asian Pac. J. Trop. Biomed., 2: 41-46.
CrossRefDirect Link - Irwan, A.B. Birawida and Anwar, 2013. [The relationship analysis of lead (Pb) levels on the dust inhalation levels of lead (Pb) in primary school children in Tamalate district, Mariso and Ujung Tanah subdistrict Makassar city]. http://repository.unhas.ac.id/bitstream/handle/123456789/5697/jurnal.pdf?sequence=.
- Jankeer, M.H. and A.A. El-Nouri, 2009. Histological study of the liver and kidney of albino mice Mus musculus exposed to lead. J. Raf. Sci., 20: 42-51.
Direct Link - Jankowska-Kulawy, A., S. Gul-Hinc, H. Bielarczyk, J.B. Suszkiw, T. Pawelczyk, A. Dys and A. Szutowicz, 2008. Effects of lead on cholinergic SN56 neuroblastoma cells. Acta Neurobiologiae Experimentalis, 68: 453-462.
Direct Link - Jolly, C. and R.I. Morimoto, 2000. Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J. Natl. Cancer Inst., 92: 1564-1572.
CrossRefDirect Link - Khanam, R., B.L. Chaudhary, S. Khanam and P. Kumar, 2011. Antibacterial activity of Marchantia palmata. Nees. Asian J. Biochem. Pharmaceut. Res., 1: 27-36.
Direct Link - Krishnan, R. and K. Murugan, 2013. Polyphenols from Marchantia polymorpha L. a bryophyta: A potential source as antioxidants. World J. Pharm. Pharmaceut. Sci., 2: 5182-5198.
Direct Link - Liu, Y., R.J. Whelan, B.R. Pattnaik, K. Ludwig and E. Subudhi et al., 2012. Terpenoids from Zingiber officinale (Ginger) induce apoptosis in endometrial cancer cells through the activation of p53. PLoS One, Vol. 7.
CrossRef - Lu, J.J., Y.Y. Dang, M. Huang, W.S. Xu, X.P. Chen and Y.T. Wang, 2012. Anti-cancer properties of terpenoids isolated from Rhizoma curcumae-A review. J. Ethnopharmacol., 143: 406-411.
CrossRefDirect Link - Mandrekar, P., D. Catalano, V. Jeliazkova and K. Kodys, 2008. Alcohol exposure regulates heat shock transcription factor binding and heat shock proteins 70 and 90 in monocytes and macrophages: Implication for TNF-α regulation. J. Leukocyte Biol., 84: 1335-1345.
CrossRefDirect Link - Matovic, V., A. Buha, D. Dukic-Cosic and Z. Bulat, 2015. Insight into the oxidative stress induced by lead and/or cadmium in blood, liver and kidneys. Food Chem. Toxicol., 78: 130-140.
CrossRefDirect Link - Mehana, E.E., A.R.M.A. Meki and K.M. Fazili, 2012. Ameliorated effects of green tea extract on lead induced liver toxicity in rats. Exp. Toxicol. Pathol., 64: 291-295.
CrossRefDirect Link - Missoun, F., M. Slimani and A. Aoues, 2010. Toxic effect of lead on kidney function in rat Wistar. Afr. J. Biochem. Res., 4: 21-27.
Direct Link - Mugahi, M.N., Z. Heidari, H.M. Sagheb and M. Barbarestani, 2003. Effects of chronic lead acetate intoxication on blood indices of male adult rat. DARU J. Pharmaceut. Sci., 11: 147-151.
Direct Link - Ramp, U., C. Mahotka, S. Heikaus, T. Shibata, M.O. Grimm, R. Willers and H.E. Gabbert, 2007. Expression of heat shock protein 70 in renal cell carcinoma and its relation to tumor progression and prognosis. Histol. Histopathol., 22: 1099-1107.
PubMedDirect Link - Reza, B., N. Ali, M. Mustafa, A. Alireza and K. Ali, 2009. Cardiac responsiveness to beta-adrenergics in rats with lead-induced hypertension. Biol. Med., 1: 75-81.
Direct Link - Valko, M., D. Leibfritz, J. Moncol, M.T.D. Cronin, M. Mazur and J. Telser, 2007. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol., 39: 44-84.
CrossRefPubMedDirect Link - Veljic, M., C. Ana, S. Marina, P. Janackovic and P.D. Marin, 2010. Antibacterial and antifungal activity of the liverwort (Ptilidium pulcherrimum) methanol extract. Arch. Biol. Sci. Belgrade, 62: 381-395.
Direct Link - Xiao, J.B., F.L. Ren and M. Xu, 2006. Flavones from Marchantia convoluta: Isolation of apigenin7O-β-D-glucuronide and 5-hydroxyl-7-methoxyl-2-methylchromone. J. Pharmaceut. Allied Sci., 3: 310-313.
Direct Link - Xiao, J.B., F.L. Ren and M. Xu, 2005. Anti-hepatitis B virus activity of flavonoids from Marchantia convoluta. Iran. J. Pharmacol. Therapeut., 4: 128-131.
Direct Link - Xiao, J.B., X.Q. Chen, Y.W. Zhang, X.Y. Jiang and M. Xu, 2006. Cytotoxicity of Marchantia convoluta leaf extracts to human liver and lung cancer cells. Braz. J. Med. Biol. Res., 39: 731-738.
CrossRefPubMedDirect Link - Yedjou, C.G., J.N. Milner, C.B. Howard and P.B. Tchounwou, 2010. Basic apoptotic mechanisms of lead toxicity in human leukemia (HL-60) cells. Int. J. Environ. Res. Public Health, 7: 2008-2017.
CrossRefDirect Link