
Raviteja Pagolu
Faculty of Biotechnology, Jeju National University, 690-756 Jeju, Republic of Korea
Md. Shafiqul Islam Sikdar
Department of Agronomy, Hajee Mohammad Danesh Science and Technology University, 5200 Dinajpur, Bangladesh
Jung-Sup Kim
Faculty of Biotechnology, Jeju National University, 690-756 Jeju, Republic of Korea
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 6 | Page No.: 221-228







ABSTRACT
Background: Chorismate synthase (EC 4.6.1.4) is the last enzyme synthesizing chorismate which is the branch point for synthesis of three aromatic amino acids in shikimate pathway in bacteria and plants. Materials and Methods: A gene encoding chorismate synthase from rice (OsCS) was analyzed and characterized by bioinformatics and molecular genetics approaches in functional complementation. Results: The analysis of a cDNA sequence revealed that the open reading frame consisted of 438 amino acid residues with approximate molecular weight of 46.9 kDa which showed high homology with related sequences from bacteria and plants. The rice OsCS was expressed in an aroC mutant of Escherichia coli strain, which was able to complement the mutant in the absence of tryptophan and inhibited the chorismate synthase activity by its inhibitor, bathophenanthroline. Conclusion: These results indicate that OsCS encodes chorismate synthase and suggest the possible application in herbicide development.
PDF Abstract XML References Citation
Received: August 30, 2016;
Accepted: September 07, 2016;
Published: September 15, 2016
How to cite this article
Raviteja Pagolu, Md. Shafiqul Islam Sikdar and Jung-Sup Kim, 2016. Characterization and Functional Complementation with an Inhibitor of Rice Chorismate Synthase. Journal of Biological Sciences, 16: 221-228.
DOI: 10.3923/jbs.2016.221.228
URL: https://scialert.net/abstract/?doi=jbs.2016.221.228
DOI: 10.3923/jbs.2016.221.228
URL: https://scialert.net/abstract/?doi=jbs.2016.221.228
INTRODUCTION
Aromatic amino acids function the building blocks synthesizing proteins as well as precursors for secondary metabolism. The three aromatic amino acids, phenylalanine (Phe), tyrosine (Tyr) and tryptophan (Trp) are synthesized from a common precursor, chorismate (Fig. 1) that originates from the shikimate pathway1-3. The chorismate is a precursor for many secondary metabolites that play important roles in the plant’s interaction with coordinated regulation with genes of associated pathways in response to altered plant development and environmental responses4. In bacteria, this pathway is almost exclusively used to produce aromatic amino acids for protein synthesis. However, in higher plants, this pathway leads to the production of numerous aromatic secondary metabolites1,3 such as tetrahydrofolate5, phylloquinone and a number of pigments6 and also components of secondary metabolic function, such as alkaloids, isoflavonoids, lignin and indole-derivatives, play important roles in pathogen resistance. Secondary metabolites are nonessential components for the primary metabolic process but are required for plant development and environmental responses4.
Chorismate synthase (CS, EC 4.6.1.4), the seventh enzyme of the shikimate pathway, catalyzes the conversion of 5-enolpyruvylshikimate 3-phosphate (EPSP) into chorismate and requires reduced flavin mononucleotide (FMN) as a cofactor.
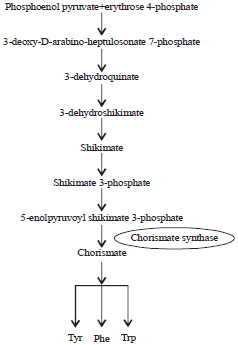 | |
| Fig. 1: | Schematic presentation of shikimate pathway |
The enzyme can bind first oxidized FMN and then EPSP to form a stable ternary complex which does not undergo turnover7.
This complex is considered to be a model of the ternary complex between enzyme, EPSP and reduced FMN immediately before catalysis commences. It is shown that the binding of oxidized FMN and EPSP to chorismate synthase affects the properties and structure of the protein. In plants, the shikimate pathway enzymes are localized in the chloroplasts8. The reaction of CS is unique in nature, involves a 1, 4 elimination of phosphate and loss of proton of the C-6 hydrogen. The formation of two out of three necessary double bonds to build an aromatic amino acid is aided by CS and activity of this enzyme requires reduced FMN molecule which is not consumed during the reaction9. The CS is insensitive to feedback inhibition of by the enzymes or final products of shikimate pathway10.
The CS has been purified and characterized from Escherichia coli 7,11,12 (E. coli), Neurospora crassa12,13, Euglena gracilis10, Bacillus subtilis14, Corydalis sempervirens15, Hordeum vulgare4, Verticillium longisporum16 and Xanthomonas oryzae17. There are little reports on the study of CS in plants and no report in rice. Therefore, in order to study the characterization and function of CS gene in rice as well as in the search of new targets for herbicide development in plants, an attempt by bioinformatics and molecular genetics approaches was performed. Here, the analysis and characterization of a gene for the CS enzyme with an inhibitor from rice (Oryza sativa), an important crop plant are reported.
MATERIALS AND METHODS
Mutant strains and plasmids: Two E. coli mutant strains which were used in this study namely a tryptophan mutant strain JW2326 (aroC) and a threonine mutant strain JW0001 (thrA). The genotypes of the mutant strains are JW2326 [Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ–, ΔaroC737::kan, rph-1, Δ(rhaD-rhaB)568, hsdR514] and JW0001 (thrA). Both mutant strains are obtained from National Bio-Resource Project (NBRP) Japan. The EST clone carrying the expected gene encoding for chorismate synthase, having the GenBank accession number of AK099850 and clone number of J013110C10 was collected from Rice Genome Resource Center (RGRC), Japan. The source of E. coli mutant strains were obtained from National Bioresource Project (NBRP), Japan.
DNA sequence analysis: The sequence analysis of rice EST clone (J013110C10) which was derived from rice cDNA library18 from developing seeds prepared in pBluescript SK-that was done by using different bioinformatics tools. The DNA sequencing and sequence analysis were described previously19. The nucleotide and amino acid sequences were compared with various sequences present in GenBank and were analyzed by using Nucleotide-BLAST20 and CLUSTALW a multiple sequence alignment program21 and GeneDoc. Motifs regions were searched by the Genome Net computation Service at Kyoto University (http://www.genome. ad.jp) and the phylogenic tree analysis was performed by MEGA 4 program22.
Polymerase chain reaction: Specific primers were designed from the sequence of OsCS information surrounding the translational start and stop codons of OsCS to amplify the full-length open reading frame (ORF) and to construct a recombinant DNA to express the gene product in E. coli. Polymerase chain reaction (PCR) was conducted in accordance with the method described previously19. The ORF region of chorismate synthase was amplified by own designed forward and reverses primers by using the PCR. After the purification of plasmid DNA from the bacterial culture harvested from LB ampicillin broth media the cDNA was amplified using the designed primers from the OsCS sequene; OsCS-F-(5’-GGAGCTCTCACCAACCAAACCAACAAC-3’) and OsCS-R-(5’-GGGTACCCTGTATAACGGATTCCTTGG-3’) both of the primers have the SacI and KpnI restriction sites. PCR was performed by using the MY Cycler TM PCR system (Bio Rad, United States) for 40 cycles carried out by denaturation at 95°C for 10 min, annealing at 58°C for 1 min and extension at 72°C for 1 min. The PCR products were analyzed by gel electrophoresis on 1% (w/v) agarose gel.
Growth curve determination in E. coli: To determine the growth of E. coli mutant with OsCS, the transformed mutant strain JW2326 harbouring pB:OsCS, control plasmid and wild type with control plasmid were inoculated in 1 mL of LB ampicillin (Amp) broth in different tubes and incubated for overnight at 37°C with shaking. The overnight culture was inoculated in the 100 mL of M9 minimal medium containing 1 mM IPTG, 20% glucose, Amp (50 μg mL–1) and all 19 amino acids except tryptophan. The E. coli growth was monitored at 37°C at 1 h interval by using measurement the optical density with a UV-spectrophotometer (UV1101, Biochrom, England) at 595 nm (OD595).
Bathophenanthroline: Bathophenanthroline was used to check the inhibitory activity on CS. This compound is also known as 4, 7 diphenyl-1, 10 phenanthroline and has molecular weight of 332.40 (Sigma-Aldrich company, USA). About 100 mL of M9 minimal medium contains 1 mM IPTG, 20% glucose, Amp (50 μg mL–1) and all 19 amino acids except tryptophan including 0.005 mM bathophenanthroline. The media was turned into red color due to formation of iron complex. The activity of E. coli was also checked with this media simultaneously media without bathophenanthroline.
RESULTS
Sequence analysis of OsCS: The EST clone (clone ID: J013110C10) obtained from the RGRC was analyzed to determine the nucleotide sequence using the designed primers and bioinformatics tools. The results of data analysis with the cDNA sequence and complete rice genome indicated that an ORF was identical to that of the rice genomic region located on chromosome III (Os03g0254800). The cDNA sequence (OsCS) contained a full-length open reading frame consisting of 1,317 bp and encoded for 438 amino acid residues and approximate molecular weight of 46.9 kDa. The expected isoelectric point of the protein was 7.922. The similar sequences of different species such as rice, Zea maize, Hordeum vulgare, Arabidopsis and many bacteria were analyzed. The most of the regions of OsCS are highly conserved in plant species sharing similar domains, but OsCS share few conserved regions compared to similar proteins of bacteria. The comparison of the predicted amino acid sequence of OsCS with the deduced sequences from A. thaliana and E. coli showed high homology with identity values of 74 and 48%, respectively (Fig. 2). Motif analysis of the amino acid sequence of OsCS revealed that there are three motifs found in the amino sequence which are located at 64...79 (GESHGGGVGCVISGCP), 172...188 (GRSSARETIGRVAAGAL) and 379...395 (RHDPCVVPRAVPMVESM), respectively (Fig. 2). The patterns of the three motifs on the OsCS amino acid sequence are G-[DES]-S-H-[GC]-x(2)-[LIVM]-[GTIVLAMS]-x-[LIVTM]-[LIVM]-[DEST]-[GH]-x-[PV], [GE]-x(2)-S-[AG]-R-x-[ST]-x(3)-[VT]-x(2)-[GA]-[STAVY]-[LIVMF] and R-[SHF]-D-[PSV]-[CSAVT]-x(4)-[SGAIVM]-x-[IVGSTAPM]-[LIVM]-x-E-[STAHNCG]-[LIVMA], respectively. The phylogenetic tree derived from the related sequences showed that OsCS is grouped with Zea mays and Hordeum vulgare in monocotyledon plants and divergent and evolved from ancestor bacterial CS using the Mega 4.1 neighbor joining program22. The branching pattern and Numbers at the nodes indicate levels of bootstrap support based on neighbor joining analyses of 1000 resample data sets using the same program (Fig. 3).
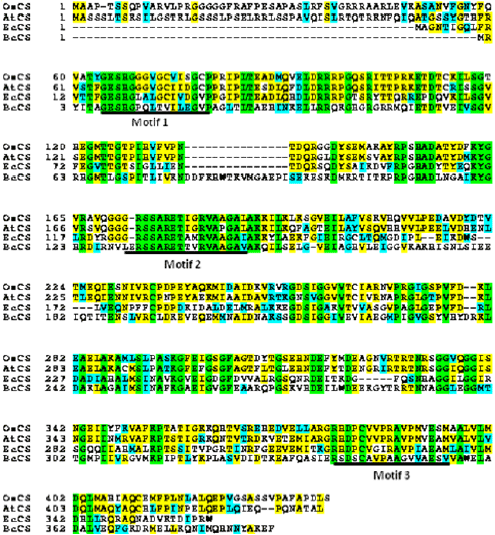 | |
| Fig. 2: | Multiple sequence alignment of CS was analyzed by using Boxshade program The amino acid sequences of chorismate synthase (CS) from different organisms that were collected from NCBI database for preparing amino acid sequence alignment of CS from Oryza sativa (OsCS accession No. AK099850), Arabidopsis thaliana (AtCS; NP_564534), Escherichia coli (EcCS; WP_020239114) and Bacillus cereus (BcCS; WP_046392823) |
Expression of OsCS can complement the aroC mutant of E. coli: The recombinant DNA, pB::OsCS was constructed using the ORF of a PCR-amplified OsCS fragment. After the transformation of E. coli with the recombinant DNA, OsCS activity was observed in vivo in a medium containing IPTG and 19 amino acids excluding Trp. Functional complementation was carried out using the CS mutant of E. coli strains JW2326 to prove the enzyme activity of the pB::OsCS product of OsCS. To evaluate the viability of E. coli cells by OsCS activity, the OsCS-expressing cells were cultured for overnight at 37°C with shaking and the diluted portion of culture was plated on agar medium containing the 19 amino acids except Trp with Amp (50 mg mL–1). The viable colonies greatly differed on the agar medium among the plasmids . The aroC mutant of E. coli JW2326 with OsCS could grow colonies under the conditions without Trp in contrast with the mutant with control plasmid showed no growth in the media without Trp due to no CS activity. This showed that the OsCS was able to functionally complement to the aroC mutant of E. coli. To determine the OsCS activity in detail, a growth activity was performed to check whether the gene would increase the sensitivity of bacterial cells to Trp. The pB::OsCS construct was transformed into the aroC mutant strain JW2326 of E. coli. A control plasmid was also transformed into the mutant strain to differentiate the activity of aroC in both cases. A transformed E. coli strain of thrB mutant (JW0001) with control plasmid was used as wild-type and negative control. The bacterial cells were grown initially incubated overnight at 37°C with 1 mL LB media with Amp and then inoculated in MM media having 1 mM IPTG, Amp (50 μg mL–1) and all 19 amino acids excluding Trp. The wild type thrB grew normally under MM media without Trp as it is evident by increased in the growth exponentially in the media with Trp.
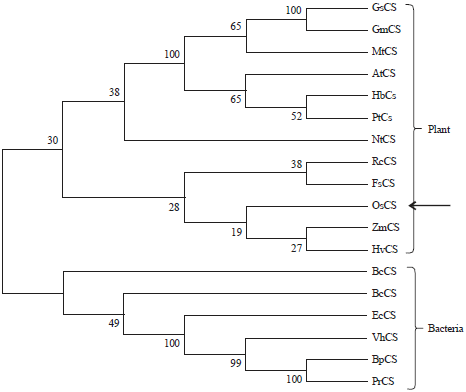 | |
| Fig. 3: | Phylogenetic tree: Phylogenetic analysis of OsTS related proteins using Clustal W and Mega 4.1 program. GenBank accession numbers of above protein sequences are as follows: KHN17131 (GsCS from Glycine soja), NP_001148583 (ZmCS; Zea mays), AES62175 (MtCS; Medicago truncatula); NP_564534 (AtCS; Arabidopsis thaliana), ADR70879 (HbCS; Hevea brasiliensis); XP_002312018 (PtCS; Populus trichocarpa), AGW47887 (NtCS; Nicotiana tabacum), XP_002529571 (RcCS; Ricinus communis), ABA54870 (FsCS; Fagus sylvatica), AK099850 (OsCS; Oryza sativa), NP_001148583 (ZmCS; Zea mays), ACF36173(HvCS; Hordeum vulgare), WP_046392823 (BcCS; Bacillus cereus), BAM51143 (BsCS; Bacillus subtilis), WP_020239114 (EcCS; Escherichia coli), CDQ35237 (VhCS; Virgibacillus halodenitrificans), WP_023559308 (BpCS; Brevibacillus panacihumi) and WP_020429593 (PrCS; Paenibacillus riograndensis) |
The E. coli mutant strains harboring control plasmid showed no growth in the media without Trp. The E. coli strain JW2326 expressing the pB:OsCS grew normally in the same media as it is evident by S-shaped growth curve. The E. coli mutant strains harboring control plasmid do not grow in the medium without Trp as they were not able to synthesize the Trp due to lacking of aroC gene. The wild type JW0001 harboring control plasmid grows normally in the media without Trp. The same E. coli strains JW2326 containing pB::OsCS grew well because the aroC mutants E. coli was able to synthesize Trp using rice CS expressed by the pB::OsCS plasmid (Fig. 4a). This is a consequence of pB::OsCS activity. From the above finding, it was concluded that OsCS expression could be functionally complement the aroC mutant of E. coli.
Bathophenanthroline inhibits the OsCS activity: Bathophenanthroline, a compound which has crystalline nature, was used as an inhibitor of OsCS activity. The bathophenanthroline (0.005 mM) was added to M9 minimal medium containing 1 mM IPTG, 20% glucose, Amp (50 μg mL–1) and all 19 amino acids except Trp. The E. coli aroC mutant strain (JW2326) and wild type (JW0001) cultures were inoculated to check the inhibitory activity into the medium with bathophenanthroline. The inhibitory activity was determined the growth curve by measuring the optical density of bacterial culture at 595 nm at 1 h intervals for 16 h. The growth of mutant strain JW2326 harboring aroC gene was decreased due to the inhibitory effect of bathophenanthroline to CS enzyme. The growth with bathophenanthroline was rapidly decreased relative to the growth in the absence bathophenanthroline in MM media without Trp. The JW2326 strain harboring control plasmid showed no growth in the media containing bathophenanthroline. The wild type JW0001 strain harboring control plasmid have sustained growth rate in the media as relative to growth in the MM media without Trp (Fig. 4b).
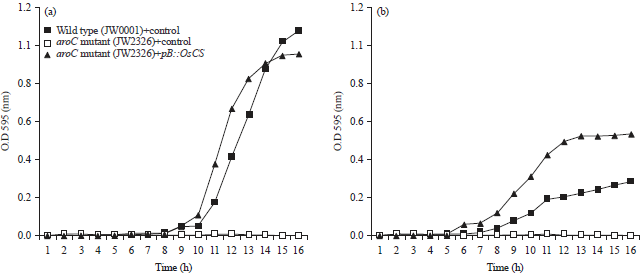 | |
| Fig. 4(a-b): | (a) Growth curves of the aroC mutant strain JW2326 harboring pB:OsCS. Bacterial cells were grown at 37°C in MM containing 19 amino acids excluding Trp. Growth was monitored via optical density measurements at 595 nm (O.D 595) and (b) Growth curve of E. coli mutant strains JW2326 harboring pB:OsCS, control plasmid and wild type JW0001 harboring control plasmid. The bacterial strains were grown in M9 minimal media without tryptophan containing 0.005 mM bathophenanthroline. Growth was monitored via optical density measurements at 595 nm (O.D 595) |
From the consequence, it was concluded that OsCS expression was able be functionally complement the aroC mutants E. coli and OsCS enzyme activity could be inhibited by bathophenanthroline.
DISCUSSION
Seven enzymes of shikimate pathway are effective targets for the development of antimicrobial and herbicidal compounds as a crucial pathway for synthesis of aromatic amino acid in bacteria and plants but not in mammals1,23,24. An inhibitor of the pathway has a damaging effect on plant metabolism and possibly is a useful commercial herbicide with low mammalian toxicity15. The shikimate pathway is targeted for the development of herbicides. For example, glyphosate, a potent inhibitor, blocks the synthesis of aromatic amino acids23 and inhibits the growth of both Gram-negative and Gram-positive bacteria in vitro25,26. Glyphosate is being widely used as a nonselective, broad-spectrum, post-emergence herbicide that is readily translocated in plants and is biodegradable by soil microorganisms27. Recent study showed that bacterial CS is an attractive target for new antibacterial drugs28. Through the effort in the search of new targets for herbicide development in plants, these results showed that OsCS has enzyme activity in the biosynthesis of amino acids and is possible to an attractive target for new herbicides. In the complementation experiment on the E. coli with an inhibitor to CS, the growth of wild type JW0001 (control) is lower than JW2326 (mutant strain complemented with OsCS) when treated with bathophenanthroline. This phenomenon might be occurred by the difference in the sensitivity of the enzyme inhibitor against E. coli and rice chorismate synthases. It might be possible to apply a new herbicide development targeting OsCS which functions crucial role in aromatic amino acid synthesis in plants. Furthermore, it is necessary to study the shikimate pathway in rice so as to better uncover the formation and regulation mechanism of diverse flavonoids in rice crop. To elucidate the impact of the shikimate pathway on the production of specialized metabolites in rice, the manipulation of the rice gene encoding CS enzyme that controlling the conversion of primary to specialized metabolism could be an attractive tool for improving rice aroma and flavor qualities.
CONCLUSION
The OsCS from rice encodes chorismate synthase and its activity is inhibited by its inhibitor, bathophenanthroline.
ACKNOWLEDGMENTS
We wish to thank Rice Genome Resource Center (RGRC) and National Bio-Resource Project (NBRP), Japan for providing an EST clone J013110C10 and an aroC mutant (JW2326) of E. coli, respectively. This study was supported by the research grant of Jeju National University in 2012.
SIGNIFICANCE STATEMENT
| • | Characterization of new gene encoding chorismate synthase from rice by bioinformatics |
| • | Functional complementation to aroC mutant of E. coli |
| • | Inhibition of OsCS activity by an inhibitor, bathophenanthroline |
REFERENCES
- Bentley, R. and E. Haslam, 1990. The shikimate pathway a metabolic tree with many branches. Crit. Rev. Biochem. Mol. Biol., 25: 307-384.
CrossRefPubMedDirect Link - Herrmann, K.M., 1995. The shikimate pathway: Early steps in the biosynthesis of aromatic compounds. Plant Cell, 7: 907-919.
CrossRefDirect Link - Wakasa, K. and A. Ishihara, 2009. Metabolic engineering of the tryptophan and phenylalanine biosynthetic pathways in rice. Plant Biotechnol., 26: 523-533.
CrossRefDirect Link - Hu, P., Y. Meng and R.P. Wise, 2009. Functional contribution of chorismate synthase, anthranilate synthase and chorismate mutase to penetration resistance in barley-powdery mildew interactions. Mol. Plant Microbe Interact., 22: 311-320.
CrossRefPubMedDirect Link - Basset, G.J.C., S. Ravanel, E.P. Quinlivan, R. White and J.J. Giovannoni et al., 2004. Folate synthesis in plants: the last step of the p-aminobenzoate branch is catalyzed by a plastidial aminodeoxychorismate lyase. Plant J., 40: 453-461.
CrossRefPubMedDirect Link - Gross, J., W.K. Cho, L. Lezhneva, J. Falk and K. Krupinska et al., 2006. A plant locus essential for phylloquinone (vitamin K1) biosynthesis originated from a fusion of four eubacterial genes. J. Biol. Chem., 281: 17189-17196.
CrossRefPubMedDirect Link - Macheroux, P., S. Bornemann, S. Ghisla and R.N.F. Thorneley, 1996. Studies with flavin analogs provide evidence that a protonated reduced FMN is the substrate-induced transient intermediate in the reaction of Escherichia coli chorismate synthase. J. Biol. Chem., 271: 25850-25858.
CrossRefPubMedDirect Link - Mousdale, D.M. and J.R. Coggins, 1985. Subcellular localization of the common shikimate-pathway enzymes in Pisum sativum L. Planta, 163: 241-249.
CrossRefPubMedDirect Link - Pitchandi, P., W. Hopper and R. Rao, 2013. Comprehensive database of Chorismate synthase enzyme from shikimate pathway in pathogenic bacteria. BMC Pharmacol. Toxicol., Vol. 14.
CrossRef - Schaller, A., M. van Afferden, V. Windhofer, S. Bulow, G. Abel, J. Schmid and N. Amrhein, 1991. Purification and characterization of chorismate synthase from Euglena gracilis. Plant Physiol., 97: 1271-1279.
CrossRefPubMedDirect Link - Morell, H., M.J. Clark, P.F. Knowles and D.B. Sprinson, 1967. The enzymic synthesis of chorismic and prephenic acids from 3-enolpyruvylshikimic acid 5-phosphate. J. Biol. Chem., 242: 82-90.
PubMedDirect Link - White, P.J., G. Millar and J.R. Coggins, 1988. The overexpression, purification and complete amino acid sequence of chorismate synthase from Escherichia coli K12 and its comparison with the enzyme from Neurospora crassa. Biochem. J., 251: 313-322.
CrossRefPubMedDirect Link - Welch, G.R., K.W. Cole and F.H. Gaertner, 1974. Chorismate synthase of Neurospora crassa: A flavoprotein. Arch. Biochem. Biophys., 165: 505-518.
CrossRefDirect Link - Hasan, N. and E.W. Nester, 1978. Purification and properties of chorismate synthase from Bacillus subtilis. J. Biol. Chem., 253: 4993-4998.
PubMedDirect Link - Schaller, A.,V. Windhofer and N. Amrhein, 1990. Purification of chorismate synthase from a cell culture of the higher plant Corydalis sempervirens Pers. Arch. Biochem. Biophys., 282: 437-442.
CrossRefPubMedDirect Link - Singh, S., S.A. Braus-Stromeyer, C. Timpner, V.T. Tran and G. Lohaus et al., 2010. Silencing of Vlaro2 for chorismate synthase revealed that the phytopathogen Verticillium longisporum induces the cross-pathway control in the xylem. Applied Microbiol. Biotechnol., 85: 1961-1976.
CrossRefPubMedDirect Link - Song, E.S., Y.J. Park, T.H. Noh, Y.T. Kim, J.G. Kim, H. Cho and B.M. Lee, 2012. Functional analysis of the aroC gene encoding chorismate synthase from Xanthomonas oryzae pathovar oryzae. Microbiol. Res., 167: 326-331.
CrossRefDirect Link - Osato, N., M. Itoh, H. Konno, S. Kondo and K. Shibata et al., 2002. A computer-based method of selecting clones for a full-length cDNA project: Simultaneous collection of negligibly redundant and variant cDNAs. Genome Res., 12: 1127-1134.
CrossRefDirect Link - Sikdar, S.I. and J. Kim, 2010. Functional analysis of a gene encoding threonine synthase from rice. Afr. J. Biotechnol., 9: 1122-1129.
Direct Link - Wheeler, D.L., D.M. Church, S. Federhen, A.E. Lash and T.L. Madden et al., 2003. Database resources of the national center for biotechnology. Nucleic Acids Res., 31: 28-33.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Kumar, S., M. Nei, J. Dudley and K. Tamura, 2008. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Briefings Bioinf., 9: 299-306.
CrossRefPubMedDirect Link - Kishore, G.M. and D.M. Shah, 1988. Amino acid biosynthesis inhibitors as herbicides. Annu. Rev. Biochem., 57: 627-663.
CrossRefPubMedDirect Link - Roberts, F., C.W. Roberts, J.J. Johnson, D.E. Kyle and T. Krell et al., 1998. Evidence for the shikimate pathway in apicomplexan parasites. Nature, 393: 801-805.
CrossRefDirect Link - Amrhein, N., J. Johanning, J. Schab and A. Schulz, 1983. Biochemical basis for glyphosate-tolerance in a bacterium and a plant tissue culture. FEBS Lett., 157: 191-196.
CrossRefDirect Link - Fischer, R.S., J.L. Rubin, C.G. Gaines and R.A. Jensen, 1987. Glyphosate sensitivity of 5-enol-pyruvylshikimate-3-phosphate synthase from Bacillus subtilis depends upon state of activation induced by monovalent cations. Arch. Biochem. Biophys., 256: 325-347.
PubMedDirect Link - Schnurer, Y., P. Persson, M. Nilsson, A. Nordgren and R. Giesler, 2006. Effects of surface sorption on microbial degradation of glyphosate. Environ. Sci. Technol., 40: 4145-4150.
CrossRefPubMedDirect Link - Hossain, M.M., P.K. Roy, A.T. Mosnaz, S.K. Shakil, M.M. Hasan and S.H. Prodhan, 2015. Structural analysis and molecular docking of potential ligands with chorismate synthase of Listeria monocytogenes: A novel antibacterial drug target. Indian J. Biochem. Biophys., 52: 45-59.
PubMedDirect Link