
M. Yang
Food and Nutritional Toxicology and Safety Laboratory, Department of Food and Animal Sciences, Alabama A and M University, 35762 Normal AL, USA
R. Hardin
Food and Nutritional Toxicology and Safety Laboratory, Department of Food and Animal Sciences, Alabama A and M University, 35762 Normal AL, USA
S. Ogutu
Laboratory of Food Chemistry, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, 35762 Normal Al, USA
M. Verghese
Laboratory of Nutritional Biochemistry and Carcinogenesis, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, 35762 Normal Al, USA
J. Boateng
Food and Nutritional Toxicology and Safety Laboratory, Department of Food and Animal Sciences, Alabama A and M University, 35762 Normal AL, USA
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 6 | Page No.: 202-214







ABSTRACT
Background and Objective: Basil and ginger possess various beneficial effects such as antioxidant, anti-inflammatory and antimicrobial. However, studies on their bioefficacy do not consider changes to composition and chemical structure from processes such as digestion which may alter their bioactivity. The objective of this study was to evaluate the bioefficacy of bioaccessible fractions of basil and ginger in a HepG2 cell model before and after simulated in vitro digestion. Methodology: Digested and non-digested basil (BD and BND) and ginger (GD and GND) extracts were prepared and used for the determinations of Total Phenolic Content (TPC), Total Flavonoid Content (TFC), Ferric Reducing Antioxidant Power (FRAP), Oxygen Radical Absorbance Capacity (ORAC) Assay and Trolox Equivalent Antioxidant Capacity (TEAC). Lactate dehydrogenase (LDH) and 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) were used as measures of efficacy. Glutathione (GSH) and Glutathione-S-Transferase (GST) were also determined. Results: Results showed that TFC was significantly (p≤0.05) increased after in vitro digestion while, a 3 fold decrease was noted in TPC. Antioxidant activities were also decreased after in vitro digestion. The BD extracts exerted significant (p≤0.05) cytotoxicity (LDH) and reduced (p≤0.05) cell viability (MTT) compared to cells treated with BND extracts. The reverse was however, observed in cells treated with GD and GND. LDH in cells treated with GND ranged from 5-53% and 6-67%, respectively for 12 and 24 h compared to cells treated with GD which ranged from 4-18% and 9-28%, respectively for 12 and 24 h. The GSH levels and GST activities were significantly (p≤0.05) higher in cells treated with BND extracts compared to cells treated with BD extracts. However, results varied with ginger extracts. Conclusion: Although, the results indicated that digested and non-digested extracts of basil and ginger induced cytotoxicity and reduced cell viability in HepG2 cells, the distinct differences in the level of efficacy may reflect alterations in the polyphenolic composition caused by the digestion process.
PDF Abstract XML References Citation
Received: July 23, 2016;
Accepted: August 15, 2016;
Published: September 15, 2016
How to cite this article
M. Yang, R. Hardin, S. Ogutu, M. Verghese and J. Boateng, 2016. Preliminary Analysis of in vitro Digestion and Bioactivity Assessment of Basil and Ginger in Human Liver Cancer Cell Line. Journal of Biological Sciences, 16: 202-214.
DOI: 10.3923/jbs.2016.202.214
URL: https://scialert.net/abstract/?doi=jbs.2016.202.214
DOI: 10.3923/jbs.2016.202.214
URL: https://scialert.net/abstract/?doi=jbs.2016.202.214
INTRODUCTION
The medicinal properties of herbs and spices is beginning to garner attention due to their known beneficial effects for human health1-3. These health-promoting properties ascribed to culinary herbs and spices are due to bioactive polyphenol compounds in these plant materials. Some of these prominent dietary sources of bioactive polyphenols such as flavonoids and phenolic acids have prompted studies of their potential health properties4,5. This growing awareness of their health relevance has led to the importance of their multiple biological effects including inhibition of cellular proliferation, modulation of enzymatic activity, regulating xenobiotic enzymes, anti-inflammatory, antimicrobial, antioxidant and anticarcinogenic activities6,7.
Basil is one of the important culinary herbs and medicinal plants. Both fresh and dried basil leaves have various food applications including in sauces, salad dressings and confectionery products as well as flavorings in drinks. Basil contributes a source of aromatic compounds and essential oils because of its distinct flavor8. Aside from its culinary applications, basil has been used as therapeutic medicine for the treatment of several symptoms.
Ginger is a popular spices used worldwide. Ginger has various food applications for example, in jams, pickles, chutneys, beverages and bakery and other food sectors9-12. It is recommended as an affective remedy for minor ailments such as gastrointestinal disorders, diarrhea, nausea, asthma, respiratory disorders, toothaches, gingivitis and arthritis in Asian folk medicine13,14.
The majority of studies on the bioefficacy of culinary herbs and spices such as basil and ginger have been conducted using cellular models. However, these studies do not take into consideration changes to the composition and chemical structure of these extracts due to human metabolic processes such as digestion which may alter these compounds. Such changes to their chemical structure can affect their bioavailability hence, bioactivity or efficacy. Secondly, there is little study on the effect of digestive processes on bioactive properties of culinary herbs and spices in general. Although, basil and ginger are only used in small quantities, they have shown several health benefits in many studies. However, data on digestion and subsequent impact on bioaccessibility and bioactivity of basil and ginger are limited. As such, more study is needed to understand if the digestion process limits their proposed health benefits. Therefore, the objective of this study was to evaluate the effects of simulated in vitro human digestion on the bioactivity (cytotoxicity and anti-proliferative properties as well as effects on antioxidant/detoxification enzymes) of basil and ginger. The outcome of this study will be useful in terms of expanding present knowledge on the in vivo therapeutic efficacy of these culinary herbs and spices.
MATERIALS AND METHODS
Materials: Basil (holy basil) and ginger were purchased from local store. Samples were stored in amber containers at -20°C until further analysis. All chemicals and reagents used were of analytical grade and were obtained from Sigma-Aldrich (St. Louis, MO) and Fisher Scientific (Suwanee, GA). Kits for cytotoxicity determinations were purchased from Roche applied Science (Indianapolis, IN).
Preparation of non-digested extracts of culinary herbs and spices: Phenolic extracts of basil and ginger were prepared using established methods. Briefly, 10 g of basil and ginger were added to 100 mL of saline buffer (pH 6.9) (Since, the gastrointestinal tract (GIT) has an aqueous environment with varying pH and would modify the amount of phenolic compounds, saline buffer was used to represent the GIT milieu) and extracted for 12 h at room temperature. The supernatants were collected via filtration using cheesecloth. Extractions were repeated 2 more times using the same procedures. Supernatants were pooled and then centrifuged at 3000×g for 20 min. The phenolic extractions were partially concentrated using a rotary evaporator (Buchi Rotavapor, Switzerland). The concentrates were then stored at -20°C until analysis. There were three replicates each. These fractions were labeled as non-digested.
Simulated human digestion of basil and ginger: A modified in vitro digestion method was used15. Basil and ginger were subjected to simulated gastrointestinal digestion which consisted of oral digestion phase (OD), gastric digestion phase (GD) and post gastric phase (PGD). Briefly, for OD basil and ginger (2 g each) was added to Simulated Salivary Fluid (SSF, pH 6.9; 0.15 M NaCl and 3 mM urea) and human salivary amylase (75 U mL–1) dissolved in simulated salivary fluid. The samples were macerated and after wards were incubated for 2 min to simulate the mechanical oral breakdown16. A blank consisting of saline was used. For GD phase, 10 mL of gastric solution (130 mM NaCl and 5 mM KCl) was added to samples after OD. Hydrochloric acid (HCl, 0.1 M) was added to each sample including the blank and pH was adjusted to pH 2.0. Pepsin solution (porcine pepsin in 0.1 M HCl) was added and all samples were incubated in an orbital water bath at 37°C for 2 h. For PGD phase, 7 mL of pancreatin/bile solution (pancreatin and bile extract in 0.1 M NaHCO3) was added and pH adjusted to 7 with NaOH (2 M). All samples were incubated with shaking for 2 h in water bath at 37°C. Afterwards, samples were heated for 15 min at 75°C to deactivate digestion enzymes and were cooled immediately on ice bath and centrifuged. Supernatants were collected and immediately kept at -80°C until analysis. The PGD (bioaccessible fraction) was used for analysis. This process was performed in triplicate.
Sample cleanup and purification of ginger and basil extracts: A clean-up procedure was incorporated into the study in order to measure the intended target groups of antioxidants and to understand the antioxidant activity of phenolic constituents. Briefly, bioaccessible and non-digested fractions of basil and ginger were loaded unto a C18 SPE cartridge after a conditioning procedure with 3 mL of acidified methanol (0.1% HCl in methanol) and 5 mL of acidified deionized water (H2O) (0.1% HCl in H2O)15. The retained phenolic compounds were eluted with acidified methanol (3×3 mL). The collected fractions were evaporated to remove solvents and re-dissolved in 50% dimethyl sulfoxide (DMSO) prior to analysis. For cell culture determinations the final concentration for DMSO did not exceed 0.05%.
Determination of Total Phenolic Content (TPC): The TPC of in vitro bioaccessible and non-digested basil and ginger extracts were determined using the folin-ciocalteu assay modified for the micro plate17. The TPC was expressed as mg Gallic acid equivalent per gram sample (mg GAE g–1 sample).
Determination Total Flavonoid Content (TFC): A colorimetric assay18 with modifications17 was used to quantify TFC. A standard curve was prepared for total flavonoid using catechin. The TFC was expressed as mg catechin equivalent per gram sample (mg CE g–1 sample).
Determination of antioxidant activity
Trolox Equivalent Antioxidant Capacity (TEAC) assay: The TEAC was determined following previous methods19. Samples were read at 734 nm. Readings were taken every minute for 6 min. The TEAC was expressed as Mmol trolox g–1 sample.
Ferric Reducing Antioxidant Power (FRAP) assay: The FRAP of in vitro bioaccessible and non-digested basil and ginger extracts was measured using methods described by Benzie and Strain20. The FRAP activity was expressed as mM Fe2+ g–1 sample.
Oxygen Radical Absorbance Capacity (ORAC) assay: For oxygen radical absorbance capacity, the protocol described previously by Cao et al.21 was used. The ORAC value was expressed as AUC and Net AUC. The AUC and the Net AUC of the standards and samples were determined using Eq. 1 and 2:
| (1) |
where, R1 is the fluorescence reading at the initiation of the reaction and Rn is the last measurement:
| (2) |
Cell line and culture conditions: Human liver cell line (HepG2), (ATCC, Manassas, VA) was grown in DMEM medium supplemented with 10% v/v fetal bovine serum, 1% v/v antibiotic solution (ATCC, Manassas, VA) at a final pH of 7.2-7.4. Cells were maintained at 37°C in an incubator under a 5% CO2/95% air atmosphere at constant humidity. The pH of the culture medium was determined using pH indicator paper. The HepG2 was seeded at 104 cells cm–2 density and allowed to adhere for 48 h, cells were used for cytotoxicity determinations on day 3 after seeding.
Lactate dehydrogenase (LDH) leakage assay: The HepG2 cells were incubated with different concentrations (12.5, 18.7, 25, 37.5, 50 and 75.0 μg mL–1) of in vitro bioaccessible and non-digested basil and ginger extracts under conditions previously described for 12 and 24 h. After incubation for these time periods, the culture medium was collected and centrifuged at 3000 rpm for 5 min in order to obtain a cell free supernatant. The activity of LDH in the supernatant was determined using a commercially available kit from Roche Diagnostics (Roche applied Science, Indianapolis, IN). Absorbance was measured at 490 nm.
(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay: The anti-proliferative properties of in vitro bioaccessible and non-digested basil and ginger extracts against HepG2 were determined by the MTT protocol as described by the manufacturer (Roche applied Science, Indianapolis, IN). Absorbance was measured at 550 and 690 nm.
Dose selection: For cell culture experiments dose levels were selected using a dose-dependent curve (12.5, 25, 37.5, 5.0 and 75.0 μg mL–1). The dose was then refined to three levels low (12.5 μg mL–1), medium (25 μg mL–1) and high (50 μg mL–1) for antioxidant/detoxification enzymes determination once a range of efficacy was established and consistent results were obtained after three separate experiments.
Preparation of cell lysate for antioxidant/detoxification enzymes determination: After treatment with basil and ginger extracts, HepG2 cells were washed with cold 1XPBS (pH 7.4) and harvested by gentle scraping. Cells were transferred into pre-chilled micro centrifuge tubes and centrifuged at 1600×g for 10 min. Cells were lysed following previous protocol22 and the supernatants which contained the cytosolic fraction were collected and used for enzyme analysis. Protein in the cell lysate was determined using BCA protein assay kit from Pierce (Rockford, IL).
Determination of endogenous antioxidant enzymes: Glutathione (GSH) from cell lysate (μmol min–1 mg–1 protein) was determined according to Griffith23. For Glutathione S-transferase (GST) activity, the method of Habig et al.24 was used. The 1-chloro-2, 4-dinitrobenzene was used as the substrate and GST was calculated as (nmol min–1 μg–1 protein). Samples were analyzed in triplicates.
Morphological evaluation of apoptosis: The HepG2 cells (1×104) were grown on coverslips and treated with or without three dose levels (low = 12.5, medium = 25 and high = 50 μg mL–1) of basil and ginger extracts for 24 h. The medium was discarded and cells were washed once with PBS. Cells were fixed with MeOH: Acetic acid (3:1) and stained with 50 mg mL–1 Hoechst 33242 dye at 37°C for 20 min. The morphological changes of apoptotic cells were also observed using phase contrast inverted microscope.
Data/statistical analysis: All experiments were performed three times (three independent experiments). Data for the chemical analysis and antioxidant assays are presented as the arithmetic Mean±Standard Deviation (SD) and those for the cytotoxicity assays are presented as the mean arithmetic percentages relative to the control±SD. Experimental data were analyzed with a one-way analysis of variance (ANOVA) followed by Tukey’s multiple range test for significant differences. The p-value of the effect had to be ≤0.05 to be considered significant. The statistical analysis was conducted using a SAS 9.3 version (SAS Institute, Inc., Cary, NC).
RESULTS
Total phenolic and flavonoid content in vitro digested and non-digested basil and ginger extracts: Total Phenolic Contents (TPC) (mg GAE g–1) of in vitro digested and non-digested basil and ginger extracts showed a wide variation among the samples (Table 1). The TPC of BD was significantly (p≤0.05) decreased compared to BND. The TPC was decreased by nearly 70% after in vitro digestion. A similar trend was also observed for ginger where, TPC after in vitro digestion was significantly (p≤0.05) decreased compared to GND (from 3.43 in GND to 0.92 mg in GD). Among the culinary samples, there were non-significant differences in TPC either before or after in vitro digestion. On the other hand, Total Flavonoids Contents (TFC) (mg CE g–1) of in vitro digested basil and ginger extracts was significantly (p≤0.05) increased compared to non-digested samples. The TFC in BD was approximately 3 times higher (15.37) compared to BND (4.31) and in GD TFC was 2 times higher (2.77) compared to GND (1.06).
Antioxidant activity of in vitro digested and non-digested basil and ginger extracts: Antioxidant activity of in vitro digested and non-digested basil and ginger extracts are shown in Table 2. The results indicated that FRAP (mM trolox g–1) activities of in vitro digested fractions of basil and ginger were significantly (p≤0.05) decreased compared to non-digested fractions.
| Table 1: | Total phenolic content and total flavonoid content of culinary herb and spice samples |
 | |
Values (n = 6) are expressed as Means±SD from three independent experiments, Means in a each row with different letters ab and xyDiffer significantly (p≤0.05) using Tukey’s studentized range test, TPC: Total phenolic content, TFC: Total flavonoid content, BND: Non-digested basil, BD: Digested basil, GND: Non-digested ginger and GD: Digested ginger | |
| Table 2: | Antioxidant activity of culinary herb and spice samples |
 | |
Values (n = 6) are expressed as Means±SD from three independent experiments, Means in a row with different letters ab and xyDiffer (p≤0.05) using Tukey’s studentized range test, FRAP is expressed in µmol FESO4 g–1, TEAC is expressed in mM trolox g–1, ORAC is expressed in µmol trolox equivalents g–1, FRAP: Ferric reducing antioxidant power, TEAC: Trolox equivalent antioxidant capacity, ORAC: Oxygen radical antioxidant capacity, BND: Non-digested basil, BD: Digested basil, GND: Non-digested ginger and GD: Digested ginger | |
The FRAP activity after in vitro digestion was decreased by 82% and by 47%, respectively for basil and ginger. A similar trend was observed with TEAC where, BND (12.16) and GND (6.81) had significantly (p≤0.05) higher TEAC activity compared to BD (7.26) and GD (4.43). For basil, TEAC was decreased by almost 40% for ginger, TEAC activity was decreased by 35% after in vitro digestion. The present results indicated that while both FRAP and TEAC activities were significantly (p≤0.05) decreased after in vitro digestion, ORAC activity showed non-significant differences among all the samples.
Cytotoxicity effect of in vitro digested and non-digested basil and ginger extracts in HepG2 cells: Cytotoxicity LDH (%) of in vitro digested and non-digested basil and ginger extracts in HepG2 liver cells are presented in Fig. 1. Results indicated that LDH (%) was significantly (p≤0.05) different between the in vitro digested and non-digested basil and ginger extracts. The BD and BND extracts exhibited a significant decrease (p≤0.05) in cytotoxicity at the lowest (12.5 μg mL–1) concentration compared to cells exposed to the highest (75 μg mL–1) concentration. For BND extracts, a dose dependent increase in cytotoxicity was noted at 12 and 24 h exposures with a 2 fold increase in cytotoxicity with increasing time. However, there was no specific dose-response relationship in cells treated with BD extracts although, cells exhibited a higher cytotoxicity compared to cells exposed to BND. In cells treated with BD, LDH (%) ranged from 8.55-24.03 and 11.50-30.72 for 12 and 24 h, respectively. Cells treated with GND extracts showed significant (p≤0.05) cytotoxic properties against HepG2 cells with increasing concentration. In cells treated with GND cytotoxicity ranged from 5.52-52.21% after 12 h incubation and from 6.13-66.97% after 24 h incubation. Unlike the results with basil, HepG2 cells treated with GD exhibited a significantly (p≤0.05) lower cytotoxicity (4-18.62% (12 h) and 8.86-28.06% (24 h)) compared to cells treated with GND.
Antiproliferation/viability effect of in vitro digested and non-digested basil and ginger extracts in HepG2 cells: In an additional step, anti-proliferative properties of ginger and basil (in vitro digested and non-digested) extracts were explored (Fig. 2).
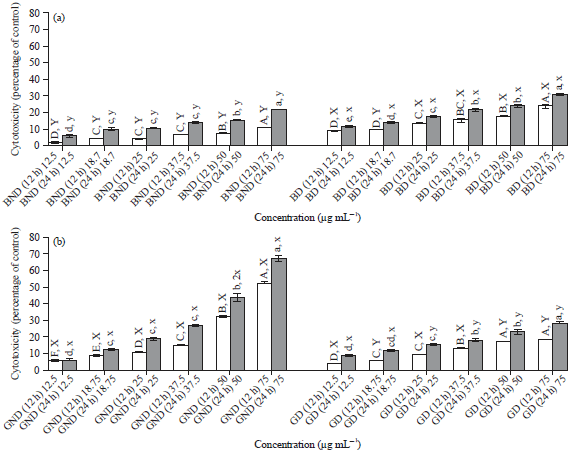 | |
| Fig. 1(a-b): | Cytotoxicity of HepG2 cells after basil treatments as measured by lactate dehydrogenase (LDH). The cells were exposed to basil and ginger extracts for 12 or 24 h as indicated. ABCDEF, abcd, XY, xyMeans on a bar with different letters differ (p≤0.05), abcCompare different concentration for 12 h, ABCCompare different concentration for 24 h, XYCompare treatments for 12 h and xyCompare treatments for 24 h. The values are expressed as Means±SD from two independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test, BND: Non-digested basil, BD: Digested basil, GND: Non-digested ginger and GD: Digested ginger |
 | |
| Fig. 2(a-b): | Cell viability of HepG2 cells after basil treatments as measured by MTT (3-[4,5-dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium bromide) assay. The cells were exposed to the compounds for 12 or 24 h as indicated. ABCD, abc, XY, xyMeans on a bar with different letters differ (p≤0.05), ABCCompare different concentration for 12 h, abcCompare different concentration for 24 h, XYCompare treatments for 12 h and xyCompare treatments for 24 h. The values are expressed as Means±SD from two independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test, BND: Non-digested basil, BD: Digested basil and GND: Non-digested ginger |
As indicated, cell proliferation/viability was significantly (p≤0.05) decreased with increasing concentration and time. At the highest concentration (75 μg mL–1), BND and BD extracts caused 54.16 and 50.61% reductions, respectively in cell viability after 12 h exposure and after 24 h, cell viability was decreased by 50.50 and 45.27%, respectively. A similar trend was observed in cells treated with ginger extracts. Cell viability in HepG2 cells treated with GND extracts ranged from 94.45-42.23% and 75-34.86% after 12 and 24 h, respectively. In cells treated with GD extracts, cell viability after 12 and 24 h exposure ranged from 90.62-58.31% and 88.25-53.24%, respectively. These results corroborate those from cytotoxicity, whereby with an increase cytotoxicity there was a decrease in cell proliferation/viability.
Effect of in vitro digested and non-digested basil and ginger extracts on glutathione S-transferase (GST) activity: The present results showed that there were non- significant differences in GST (nmol mL–1 μg–1 protein) activity in cells treated with BND extracts and the control untreated cells (Fig. 3). However, GST activity was significantly (p≤0.05) increased after 24 h in cells treated with high (50 μg mL–1) concentration of BND extracts. On the other hand, GST activity in cells treated with BD extracts was significantly (p≤0.05) decreased (with time and concentration) compared to the control untreated cells. There was atleast a 2 folds decrease in GST activity in cells treated with medium (25 μg mL–1) and high (50 μg mL–1) concentrations of BD extracts compared to the control untreated cells. The results also showed that GST activity was higher in cells treated with BND compared to cells treated with BD. The GST activity in cells treated with low (12.5 μg mL–1) and medium (25 μg mL–1) concentrations of GND extracts was similar to the control (p≤0.05) although, a significant decrease (p≤0.05) was noted in cells treated with high concentration of the extract. In cells treated with GD extracts, GST activity was significantly (p≤0.05) decreased after 12 h incubation when compared to the control.
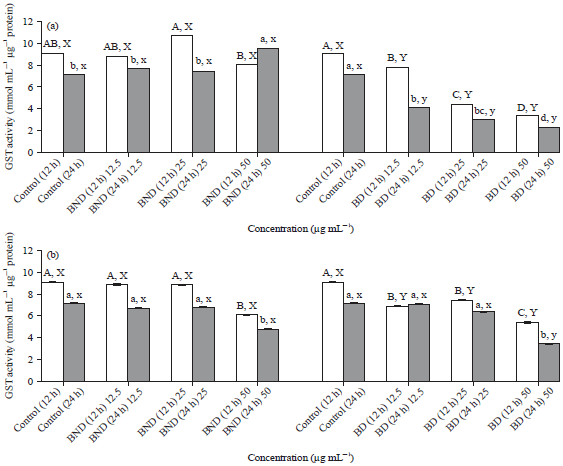 | |
| Fig. 3(a-b): | Effect of non-digested and digested basil extracts on Glutathione S-transferase (GST) activity in HepG2 cells. ABC, abc, XY, xyMeans on a bar with different letters differ (p≤0.05), ABCCompare different concentration for 12 h, abcCompare different concentration for 24 h, XYCompare treatments for 12 h and xyCompare treatments for 24 h. The values are expressed as Means±SD from two independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test, BND: Non-digested basil, BD: Digested basil, GND: Non-digested ginger and GD: Digested ginger |
But, there was non-significant difference in GST activity after 24 h, except in cells treated with high concentrations of the extract, where at least a 2 folds decrease in GST activity was noted when compared to the control.
Effect of in vitro digested and non-digested basil and ginger extracts on glutathione (GSH) levels: Results shown in Fig. 4 indicated that there were non-significant differences in GSH (μM min–1 mg–1 protein) levels in the cells treated with BND (at all concentrations and exposure time) when compared to the control untreated cells. By contrast, GSH levels in cells treated with BD extracts were significantly (p≤0.05) decreased in comparison to the control untreated cells. The GSH levels were decreased by approximately 33-64% (12 h) and 68-76% (24 h) compared to the control untreated cells. Overall, GSH levels were significantly (p≤0.05) increased in cells treated with BND compared to BD. For GND, GSH levels were significantly (p≤0.05) decreased in cells treated with medium (25 μg mL–1) and high (50 μg mL–1) concentrations compared to the control by a range of approximately 1.5-2.8 folds and 1.6-3 folds after 12 and 24 h incubation, respectively. After 12 h incubation, GSH level was significantly (p≤0.05) increased in cells treated with GD extracts (except in cells treated with high concentration) when compared to the control untreated cells but decreased (p≤0.05) after prolong (24 h) incubation.
Effect of in vitro digested and non-digested basil and ginger extracts on cellular morphology of HepG2 cells: Cellular apoptotic morphology via fluorescence microscopy indicate untreated cells stained with Hoechst 33258 (Fig. 5) cells were large and rounded with uniformly light blue nuclei were observed under fluorescence microscope while, apoptotic cells exhibited bright blue because of chromatin condensation. The study results showed that apoptotic cells induced by digested and non-digested basil and ginger extracts displayed condensed and fragmented nuclei.
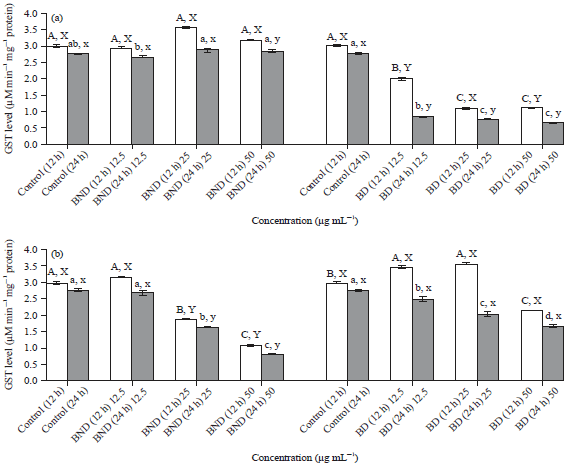 | |
| Fig. 4(a-b): | Effect of non-digested and digested basil extracts on Glutathione (GSH) levels in HepG2 cells. ABC,abcMeans on a bar with different letters differ (p≤0.05), ABCCompare different concentration for 12 h, abcCompare different concentration for 24 h, XYCompare treatments for 12 h and xyCompare treatments for 24 h. The values are expressed as Means±SD from two independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test, BND: Non-digested basil, BD: Digested basil, GND: Non-digested ginger and GD: Digested ginger |
Cells stained with geimsa and viewed via phase contrast microscopy (Fig. 6) indicated that at high concentrations, cells displayed DNA fragmentation, organelle swelling and shrunken nuclei.
DISCUSSION
Nutritional efficacy of food products is crucial especially, when studying the role of bioactive compounds in human health. According to Carbonell-Capella et al.25 before concluding on any potential health effect on any food products, it is imperative to determine whether the digestion process affects bioactive compounds and their stability as this, in turn will affect their possible beneficial effects including antioxidant, anti-inflammatory, antitumor and regulating xenobiotic enzymes26. In this study, TPC which is representative for phytochemicals were found at varying levels after in vitro digestion of basil and ginger. Conversely, TFC were significantly (p≤0.05) higher in basil and ginger after in vitro digestion. The present results are in line with Bouayed et al.27 who also indicated in an in vitro study that the phenolic content in digested fruit beverages were decreased by atleast 47% compared to non-digested samples. In another study, only 62% of polyphenols in grapes were bioaccessible following gastric and intestinal digestion28. Previous studies have also indicated a significant loss in TPC contents in green tea29, bean seed coats30 and spent coffee grounds31 after in vitro GI digestion. According to Bouayed et al.27 the nature of extractable phytochemicals, their stability and their antioxidant activity depend on many factors such as the food matrix, pH, temperature presence of inhibitors or enhancers of absorption, presence of enzymes, host and other related factors. As such, it is speculated that the transition from the acidic gastric environment to the mild alkaline post gastric environment caused a decrease in the amount of bioaccessible phenolics.
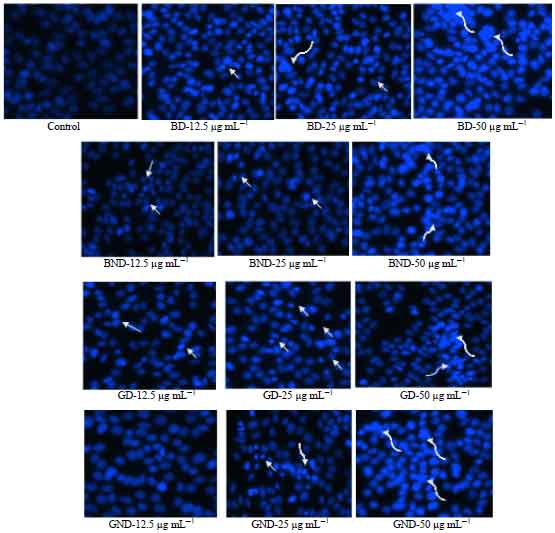 | |
| Fig. 5: | Morphological changes during basil and ginger extracts induced cytotoxicity in HepG2 cells as detected by Hoechst 33258 staining. The HepG2 cells were treated with basil and ginger extracts at 12.5, 25 and 50 µg mL–1 for 24 h. Control (live) cells were large and rounded with uniformly light blue nuclei were observed under fluorescence microscope while, apoptotic cells exhibited bright blue because of chromatin condensation. The short arrow marks indicate condensation. The curve arrow indicates fragmentation |
The FRAP and TEAC activities were significantly (p≤0.05) decreased in basil and ginger after in vitro digestion. According to Chen et al.32, the FRAP and TEAC activities of Chimonanthus praecox (L.) Link and Redartfulplum tea were decreased after in vitro digestion. Rodriguez-Roque et al.33 demonstrated that the antioxidant activities of blended fruit juice were unstable under intestinal conditions which has an alkaline pH. This can lead to decreased antioxidant activity. Likewise, in vitro digestion perhaps, influenced the FRAP and TEAC activities of digested basil and ginger. Rice-Evans et al.34 reported that the chemical structure of phenolic compounds could play a critical role in free radical scavenging activity. The study results revealed that antioxidant activity of basil and ginger extracts was not well correlated with the content of the phenolics and flavonoids compounds. Although, polyphenols provide the major antioxidant potency of the culinary herbs and spices used in this study, the present observations indicate that digestion may have altered the antioxidant properties due in part to the variations in polyphenol contents.
Herbs and spices have been studied for their effects against cell viability and cytotoxicity. It has been indicated in many studies that the anticancer/tumor, anti- inflammatory properties of herbs and spices are attributed to their ability to inhibit cancer cell growth or proliferation and to cause cancer cells to undergo cell damage via., apoptosis. In the present study, it is showed that in vitro digested basil and ginger extracts as well as the non-digested extracts induced HepG2 cells to undergo cytotoxicity and inhibited cell proliferation.
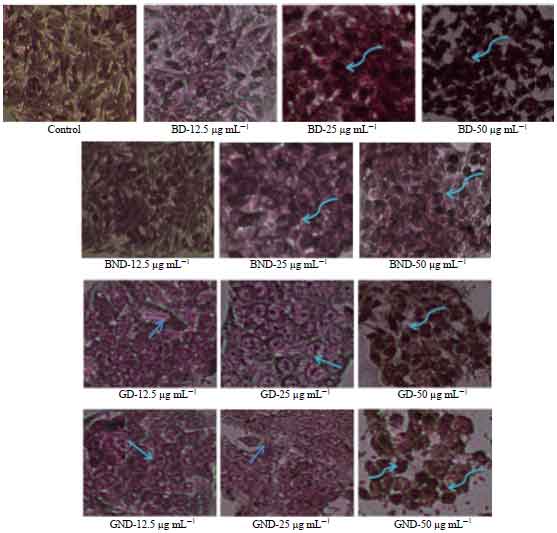 | |
| Fig. 6: | Effect of non-digested and digested basil and ginger extracts on apoptotic morphological changes in HepG2 cells detected by geimsa staining after 24 h of incubation. The arrow indicate cell blebbing, curved arrow indicate chromatin condensation and DNA fragmentation, BD: Basil digested, BND: Basil non-digested, GD: Digested ginger and GND: Ginger non-digested |
The present study is in line with other studies that have shown cytotoxicity and anti-proliferative effects of extracts derived from spice extracts. Recently, Offei-Oknye et al.35 demonstrated the cytotoxic properties via LDH of different forms (fresh, oven dried and freeze dried) of ginger extracts in HepG2 cells. Padma et al.36 and Wang et al.37 also reported results indicating the anti-proliferative and cytotoxic effects of ginger in Hela cells. In the former study, one of the major mechanisms ascribed to the presented effect was suggested to be the involvement of reactive oxygen species in apoptosis induction. This could be attributed to the presence of different classes of compounds in ginger, mainly gingerols and shogaols. For example, gingerol was found to exert cytotoxic and anti-proliferative inhibitory effects on the viability and DNA synthesis of human promyelocytic leukemia (HL-60) cells37,38.
The present study is also in line with a study reported by Lantto et al.39 who indicated that basil extracts, exhibited significant increase in cytotoxicity as assessed by LDH assay after 12 h of treatment in SH-SY5Y neuroblastoma cells. The researchers also reported atleast a 50% decrease in cell viability in SH-SY5Y cells exposed to basil extracts. According to the researchers, basil had an influence on the membrane integrity and amount of p53 in SH-SY5Y cells. In this study, a 50% decrease in cell viability in HepG2 cells exposed to BND and BD extracts (at highest concentrations). Another study revealed that extracts and the essential oil of basil significantly inhibited the aggressiveness of pancreatic cancer cells through MTT and even inhibited the growth of implanted cancerous cells in rodent models40. The present study provides evidence that bioaccessible fractions of ginger and basil acted as a potent growth inhibitory compound in HepG2 and thus, might suggest the possibility of chemoprevention potential in liver cancer. This study showed that HepG2 cells were more sensitive to GND compared to GD. However, the effects were reversed when cells were treated with basil extracts.
Results from this study indicated that GND, GD and BD extracts at high concentrations (especially, after prolonged incubation) led to significant (p≤0.05) decrease in endogenous GSH levels and GST antioxidant activity. It is possible that the observed effects may be due to the concentration of the polyphenol utilized since, high doses of some phenolic compounds may be pro-oxidant and negatively affect cell growth and viability. Recently, Weisburg et al.41 explained that studies utilizing human cells in vitro to evaluate the anticancer properties of dietary plant-derived polyphenols have showed that polyphenols by acting as pro-oxidants, generate sufficient levels of ROS to induce oxidative stress thus, leading to apoptosis and mitochondrial membrane damage of carcinoma cells. The decline in endogenous antioxidant activity i.e., GSH and GST is according to Valentovic et al.42 indicative of oxidative stress prior to cytotoxicity and the loss of membrane integrity. Hence, the decrease in endogenous antioxidants observed in cells treated with high concentrations of GD, GND and BD extracts might correlate with the higher anti proliferative and cytotoxicity index observed.
In a study by Franco and Cidlowski43 aimed to evaluate the role of GSH in the regulation of apoptosis, the researchers implied that GSH depletion is a common feature of apoptotic cell death. As a major antioxidant, GSH participates in the disposal of potentially harmful electrophiles and by this means, protects cells from damaging effects of free radicals and ROS44,45. Therefore, its depletion renders cell particularly vulnerable to oxidative stress. On the other hand, some studies utilizing high concentrations of plant extracts have reported increased levels in GSH35 as observed in the current study when HepG2 cells were treated with BND extracts. These results may suggest an attempt by the cells to recover from the aggression to which they were submitted46. Yet again, Ortega et al.47 emphasized that GSH synthesis could be up-regulated during oxidative stress by phenolic compounds and oxidative stressors that is if their levels do not compromise cell viability, (as observed in cells treated with low concentrations of GND, GD and BD extracts). According to the researchers provides the paradox that is associated with the protective mechanisms of GSH against oxidative stress.
The current data indicated that cells treated with high concentrations of GND, GD and BD extracts decreased GST activity compared to cells treated with low concentrations. Rao and Shaha48 pointed out that inhibition of GST activity could lead to an increase in the formation of lipid peroxidation such as GSTs may serve as a useful parameter of products of oxidative stress. It has been suggested that GST catalyzes the conjugation of GSH to yield hydrophobic molecules that are less reactive than the parental compound and participates in reactions that destroy hydrogen peroxide and organic peroxides49,50. Therefore, it is possible that the depletion of GSH may have been a result of the inhibition of GST activity.
Morphological determinations of basil and ginger extracts-treated cells demonstrated that the extracts induced cell shrinkage and chromatin condensation. These observations suggest that in vitro digested basil and ginger extracts induced apoptosis in HepG2 cells. The results also raises the possibility that in vitro digested extracts obtained from basil and ginger possesses distinct bioactivity.
CONCLUSION
The current study was undertaken to determine the effects of in vitro digestion on the bioactivity of basil and ginger. This study indicated that digested and non-digested extracts of basil and ginger induced cytotoxicity and reduced cell viability in HepG2 cells. Although, the data indicated some variations in bioefficacy of digested basil and ginger extracts with regard to cytotoxicity, this may be a result of differences in the putative active components. While, bioefficacy of herbs and spices are reported studies investigating their beneficial effects post digestion are scarce. This study may provide baseline information which may be beneficial for consumers, nutritionists and food manufactures since, basil and ginger are widely utilized herbs and spices. For future studies, it is planned to identify and determine the concentrations of these bioactive compounds in hopes that these compounds with different bioactive profiles could explain the varied responses seen in this study. Basil and ginger may have an enhanced bioaccessibility after in vitro digestion but compounds contributing to their bioactivity require further investigation.
SIGNIFICANCE STATEMENTS
| • | Polyphenols and antioxidant activity of in vitro digested and non-digested basil and ginger extracts were evaluated |
| • | In vitro digestion reduced the levels of total phenols and antioxidant activity |
| • | Cytotoxicity, anti-proliferative properties and effect on antioxidant enzymes were evaluated |
| • | Study may provide baseline information which may be beneficial for consumers, nutritionists and food manufactures |
ACKNOWLEDGMENT
This study was funded by the USDA capacity Building grant No. 11343134, Alabama A and M University Agriculture Research Station, Normal AL 35762.
REFERENCES
- Shobana, S. and K.A. Naidu, 2000. Antioxidant activity of selected Indian spices. Prostaglandins Leukot. Essent. Fatty Acids, 62: 107-110.
CrossRefPubMedDirect Link - Jungbauer, A. and S. Medjakovic, 2012. Anti-inflammatory properties of culinary herbs and spices that ameliorate the effects of metabolic syndrome. Maturitas, 71: 227-239.
CrossRefDirect Link - Dzoyem, J.P., V. Kuete, L.J. McGaw and J.N. Eloff, 2014. The 15-lipoxygenase inhibitory, antioxidant, antimycobacterial activity and cytotoxicity of fourteen ethnomedicinally used African spices and culinary herbs. J. Ethnopharmacol., 156: 1-8.
CrossRefDirect Link - Hinneburg, I., H.J.D. Dorman and R. Hiltunen, 2006. Antioxidant activities of extracts from selected culinary herbs and spices. Food Chem., 97: 122-129.
CrossRefDirect Link - Wojdylo, A., J. Oszmianski and R. Czemerys, 2007. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem., 105: 940-949.
CrossRefDirect Link - Del Rio, D., A. Rodriguez-Mateos, J.P.E. Spencer, M. Tognolini, G. Borges and A. Crozier, 2013. Dietary (Poly)phenolics in human health: Structures, bioavailability and evidence of protective effects against chronic diseases. Antioxid. Rredox Signal., 18: 1818-1892.
CrossRefDirect Link - Vallverdu-Queralt, A., J. Regueiro, M. Martinez-Huelamo, J.F.R. Alvarenga, L.N. Leal and R.M. Lamuela-Raventos, 2014. A comprehensive study on the phenolic profile of widely used culinary herbs and spices: Rosemary, thyme, oregano, cinnamon, cumin and bay. Food Chem., 154: 299-307.
CrossRefDirect Link - Ghanjaoui, M.E., M.L. Cervera, M. El Rhazi and M. de la Guardia, 2011. Validated fast procedure for trace element determination in basil powder. Food Chem., 125: 1309-1313.
CrossRefDirect Link - Sacchetti, G., S. Maietti, M. Muzzoli, M. Scaglianti, S. Manfredini, M. Radice and R. Bruni, 2005. Comparative evaluation of 11 essential oils of different origin as functional antioxidants, antiradicals and antimicrobials in foods. Food Chem., 91: 621-632.
CrossRefDirect Link - Tapsell, L.C., I. Hemphill, L. Cobiac, C.S. Patch and D.R. Sullivan et al., 2006. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust., 185: S4-S24.
PubMedDirect Link - Zhao, X., Z. Yang, G. Gai and Y. Yang, 2009. Effect of superfine grinding on properties of ginger powder. J. Food Eng., 91: 217-222.
CrossRefDirect Link - Mesomo, M.C., A. de Paula Scheer, E. Perez, P.M. Ndiaye and M.L. Corazza, 2012. Ginger (Zingiber officinale R.) extracts obtained using supercritical CO2 and compressed propane: Kinetics and antioxidant activity evaluation. Supercritical Fluids, 71: 102-109.
CrossRefDirect Link - Ali, B.H., G. Blunden, M.O. Tanira and A. Nemmar, 2008. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol., 46: 409-420.
CrossRefPubMedDirect Link - Butt, M.S. and M.T. Sultan, 2011. Ginger and its health claims: Molecular aspects. Crit. Rev. Food Sci. Nutr., 51: 383-393.
CrossRefDirect Link - Wang, D., B.A. Williams, M.G. Ferruzzi and B.R. D'Arcy, 2013. Microbial metabolites, but not other phenolics derived from grape seed phenolic extract, are transported through differentiated Caco-2 cell monolayers. Food Chem., 138: 1564-1573.
CrossRefDirect Link - Mandalari, G., C. Bisignano, A. Filocamo, S. Chessa and M. Saro et al., 2013. Bioaccessibility of pistachio polyphenols, xanthophylls and tocopherols during simulated human digestion. Nutrition, 29: 338-344.
CrossRefDirect Link - Gajula, D., M. Verghese, J. Boateng, L.T. Walker, L. Shackelford, S.R. Mentreddy and S. Cedric, 2009. Determination of total phenolics, flavonoids and antioxidant and chemopreventive potential of basil (Ocimum basilicum L. and Ocimum tenuiflorum L.). Int. J. Cancer Res., 5: 130-143.
CrossRefDirect Link - Kim, D.O., O.K. Chun, Y.J. Kim, H.Y. Moon and C.Y. Lee, 2003. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem., 51: 6509-6515.
CrossRefDirect Link - Montoro, P., A. Braca, C. Pizza and N. de Tommasi, 2005. Structure-antioxidant activity relationships of flavonoids isolated from different plant species. Food Chem., 92: 349-355.
CrossRefDirect Link - Benzie, I.F.F. and J.J. Strain, 1996. The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal. Biochem., 239: 70-76.
CrossRefPubMedDirect Link - Cao, G., H.M. Alessio and R.G. Cutler, 1993. Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med., 14: 303-311.
CrossRefDirect Link - Boateng, J. and M. Verghese, 2012. Protective effects of the phenolic extracts of fruits against oxidative stress in human lung cells. Int. J. Pharmacol., 8: 152-160.
CrossRefDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Carbonell-Capella, J.M., M. Buniowska, F.J. Barba, M.J. Esteve and A. Frigola, 2014. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compreh. Rev. Food Sci. Food Saf., 13: 155-171.
CrossRefDirect Link - Fernandez-Garcia, E., I. Carvajal-Lerida and A. Perez-Galvez, 2009. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res., 29: 751-760.
CrossRefDirect Link - Bouayed, J., L. Hoffmann and T. Bohn, 2011. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem., 128: 14-21.
CrossRefDirect Link - Tagliazucchi, D., E. Verzelloni, D. Bertolini and A. Conte, 2010. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem., 120: 599-606.
CrossRefDirect Link - Marchese, A., E. Coppo, A.P. Sobolev, D. Rossi, L. Mannina and M. Daglia, 2014. Influence of in vitro simulated gastroduodenal digestion on the antibacterial activity, metabolic profiling and polyphenols content of green tea (Camellia sinensis). Food Res. Int., 63: 182-191.
CrossRefDirect Link - Sancho, R.A.S., V. Pavan and G.M. Pastore, 2015. Effect of in vitro digestion on bioactive compounds and antioxidant activity of common bean seed coats. Food Res. Int., 76: 74-78.
CrossRefDirect Link - Campos-Vega, R., K. Vazquez-Sanchez, D. Lopez-Barrera, G. Loarca-Pina, S. Mendoza-Diaz and B.D. Oomah, 2015. Simulated gastrointestinal digestion and in vitro colonic fermentation of spent coffee (Coffea arabica L.): Bioaccessibility and intestinal permeability. Food Res. Int., 77: 156-161.
CrossRefDirect Link - Chen, G.L., S.G. Chen, Y.Q. Xie, F. Chen, Y.Y. Zhao, C.X. Luo and Y.Q. Gao, 2015. Total phenolic, flavonoid and antioxidant activity of 23 edible flowers subjected to in vitro digestion. J. Funct. Foods, 17: 243-259.
CrossRefDirect Link - Rodriguez-Roque, M.J., M.A. Rojas-Grau, P. Elez-Martinez and O. Martin-Belloso, 2013. Changes in vitamin C, phenolic and carotenoid profiles throughout in vitro gastrointestinal digestion of a blended fruit juice. J. Agric. Food Chem., 61: 1859-1867.
CrossRefDirect Link - Rice-Evans, C.A., N.J. Miller and G. Paganga, 1996. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Biol. Med., 20: 933-956.
CrossRefPubMedDirect Link - Offei-Oknye, R., J.L. Patterson, L.T. Walker, J. Boateng and M. Verghese, 2015. Chemopreventive potential of ginger on Hep2G cells. Int. J. Cancer Res., 11: 52-66.
CrossRefDirect Link - Padma, V., S.A.D. Christie and K.M. Ramkuma, 2007. Induction of apoptosis by ginger in HEp-2 cell line is mediated by reactive oxygen species. Basic Clin. Pharmacol. Toxicol., 100: 302-307.
CrossRefPubMedDirect Link - Wang, C.C., L.G. Chen, L.T. Lee and L.L. Yang, 2003. Effects of 6-gingerol, an antioxidant from ginger, on inducing apoptosis in human leukemic HL-60 cells. In vivo, 17: 641-645.
PubMedDirect Link - Lee, E. and Y.J. Surh, 1998. Induction of apoptosis in HL-60 cells by pungent vanilloids, [6]-gingerol and [6]-paradol. Cancer Lett., 134: 163-168.
CrossRefPubMedDirect Link - Lantto, T.A., M. Colucci, V. Zavadova, R. Hiltunen and A. Raasmaja, 2009. Cytotoxicity of curcumin, resveratrol and plant extracts from basil, juniper, laurel and parsley in SH-SY5Y and CV1-P cells. Food Chem., 117: 405-411.
CrossRefDirect Link - Shimizu, T., M.P. Torres, S. Chakraborty, J.J. Souchek and S. Rachagani et al., 2013. Holy Basil leaf extract decreases tumorigenicity and metastasis of aggressive human pancreatic cancer cells in vitro and in vivo: Potential role in therapy. Cancer Lett., 336: 270-280.
CrossRefDirect Link - Weisburg, J.H., A.G. Schuck, S.E. Reiss, B.J. Wolf, S.R. Fertel, H.L. Zuckerbraun and H. Babich, 2013. Ellagic acid, a dietary polyphenol, selectively cytotoxic to HSC-2 oral carcinoma cells. Anticancer Res., 33: 1829-1836.
PubMedDirect Link - Valentovic, M.A., J.G. Ball, J.M. Brown, M.V. Terneus and E. McQuade et al., 2014. Resveratrol attenuates cisplatin renal cortical cytotoxicity by modifying oxidative stress. Toxicol. In vitro, 28: 248-257.
CrossRefDirect Link - Franco, R. and J.A. Cidlowski, 2009. Apoptosis and glutathione: Beyond an antioxidant. Cell Death Differ., 16: 1303-1314.
CrossRefPubMedDirect Link - Du, C., Z. Gao, V.A. Venkatesha, A.L. Kalen and L. Chaudhuri et al., 2009. Mitochondrial ROS and radiation induced transformation in mouse embryonic fibroblasts. Cancer Biol. Therapy, 8: 1962-1971.
CrossRefDirect Link - Anderson, M.E., 1998. Glutathione: An overview of biosynthesis and modulation. Chem. Biol. Interac., 111-112: 1-14.
CrossRefDirect Link - Fernandes, F., C. Sousa, F. Ferreres, P. Valentao, F. Remiao, J.A. Pereira and P.B. Andrade, 2012. Kale extract increases glutathione levels in V79 cells, but does not protect them against acute toxicity induced by hydrogen peroxide. Molecules, 17: 5269-5288.
CrossRefDirect Link - Ortega, A.L., S. Mena and J.M. Estrela, 2011. Glutathione in cancer cell death. Cancers, 3: 1285-1310.
CrossRefDirect Link - Rao, A.V.S.K. and C. Shaha, 2000. Role of glutathione S-transferases in oxidative stress-induced male germ cell apoptosis. Free Radic. Biol. Med., 29: 1015-1027.
CrossRefPubMedDirect Link - Mari, M. and A.I. Cederbaum, 2001. Induction of catalase, alpha and microsomal glutathione S-transferase in CYP2E1 overexpressing HepG2 cells and protection against short-term oxidative stress. Hepatology, 33: 652-661.
CrossRefPubMedDirect Link - Garcia-Alonso, F.J., A. Guidarelli and M.J. Periago, 2007. Phenolic-rich juice prevents DNA single-strand breakage and cytotoxicity caused by tert-butylhydroperoxide in U937 cells: The role of iron chelation. J. Nutr. Biochem., 18: 457-466.
CrossRefDirect Link