
E.T.S. Putra
Faculty of Agriculture, Universitas Gadjah Mada, Bulaksumur, 55281 Yogyakarta, Indonesia
Issukindarsyah
Faculty of Agriculture, Universitas Gadjah Mada, Bulaksumur, 55281 Yogyakarta, Indonesia
Taryono
Faculty of Agriculture, Universitas Gadjah Mada, Bulaksumur, 55281 Yogyakarta, Indonesia
B.H. Purwanto
Faculty of Agriculture, Universitas Gadjah Mada, Bulaksumur, 55281 Yogyakarta, Indonesia
D. Indradewa
Faculty of Agriculture, Universitas Gadjah Mada, Bulaksumur, 55281 Yogyakarta, Indonesia
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 5 | Page No.: 155-166







ABSTRACT
Objective: The objectives of the study were to determine (1) The level of biochemical resistance to drought stress on oil palm using boron (B) and silicon (Si) applications and (2) The mechanism of B and Si actions in the induction of biochemical resistance of oil palm seedlings to drought stress. Materials and Methods: Field trial was arranged in the Randomized Complete Block Design (RCBD) factorial using three blocks as replications. The first factor was the dose of B, namely 0.00, 0.17, 0.44, 0.87, 1.31 and 1.75 g per seedling. The second factor was the dose of Si, namely 0.00, 1.15, 2.31, 3.46 and 4.69 g per seedling. The observations were done on the contents of proline and total phenolic compound, total antioxidant, H2O2 activities, dry weight, trunk height and diameter. The data were analysed using analysis of variance (ANOVA) and the means were separated using Duncan’s Multiple Range Test (DMRT) at 5% level. Meanwhile, the optimum dose of B and Si were determined using regression analysis. Results: The accumulations of proline in leaf tissue reach a maximum at 0.81 g of B per seedling. The application of 1.75 g of B per seedling was significantly able to maximize the rate of total phenolic compound accumulation, total antioxidant activity and inhibit H2O2 activity. On the other hand, the application doses of Si that able to maximize the rate of proline accumulation, total phenolic compound and total antioxidant activity were 1.15, 0.73 and 2.82 g per seedling, respectively. Hydrogen peroxide activity begins to compress when the application dose of Si was more than 4.29 g per seedling. Conclusion: The application of B and Si were able to induce biochemical resistance of the oil palm seedlings to drought stress through the accumulations of proline and total phenolic compound, increase of the total antioxidant activity and inhibition of H2O2 activity.
PDF Abstract XML References Citation
Received: June 02, 2016;
Accepted: June 07, 2016;
Published: June 15, 2016
How to cite this article
E.T.S. Putra, Issukindarsyah, Taryono, B.H. Purwanto and D. Indradewa, 2016. Induction of Biochemical Resistance of Oil Palm Seedlings to
Drought Stress Using Boron and Silicon Applications. Journal of Biological Sciences, 16: 155-166.
DOI: 10.3923/jbs.2016.155.166
URL: https://scialert.net/abstract/?doi=jbs.2016.155.166
DOI: 10.3923/jbs.2016.155.166
URL: https://scialert.net/abstract/?doi=jbs.2016.155.166
NTRODUCTION
Oil palm’s response to drought is generally negative. Previous studies provide information that the drought in the area of oil palm was decreasing the yield of fresh fruit bunches up to 26.30%1-3. This information suggests that oil palm is a commodity that is quite sensitive to drought, so it is quite urgent to be solved4. The next few years, drought becomes more frequent with heavy intensity due to the phenomenon of global climate change5, particularly in the area of oil palm plantations. Agronomic approaches can be taken to reduce the sensitivity of oil palm to drought. The approach is intended to stimulate biochemical character changes in the oil palm tissue so that the biochemical resistance of oil palm to drought stress can be realized.
Boron (B) is an essential nutrient that is needed by plants to grow normally6, especially when the plants are exposed to drought stress and oxidative damage threatened. Application of B to the plant that is exposed to drought is able to reduce the level of oxidative damage because B stimulates the synthesis of several types of antioxidants that has a role to eliminate the negative effects of Reactive Oxygen Species (ROS). Synthesis of proline and phenolic compounds in plant tissues which exposed to drought also stimulated by the adequacy of B. Proline is a simple organic compound group of proteins that are used by plants in the process of osmotic adjustment to induce the biochemical resistance mechanism’s oil palm to drought, while phenol is the result of secondary metabolites that can act as non-enzymatic antioxidants7,8. Non-enzymatic antioxidants in the form of phenolic compounds have a role in the inhibition of the synthesis and activity of ROS.
Silicon (Si) is not an essential nutrient for plants, but it gives positive influence on the growth and development of plants, especially plants exposed to drought9,10. Some studies indicate that Si is able to avoid the crop from damage when it grown under the environmental that exposed by drought. These conditions indicate that the application of Si improves drought resistance11,12. Mechanisms associated with the role of Si for resistance to drought are an increase in the content of several types of solutes in the cytoplasm in the form of glucose, sucrose, raffinose, total soluble protein and phenol compounds. This mechanism is often referred to as osmotic adjustment due to decreased of cell osmotic potential to compatible with the soil solution osmotic potential, osmotic potential decrease, however, is not to endanger the life of the cell13,14. Associated with growth hormone, the application of Si is able to increase the accumulation of auxin, gibberellins and cytokinins in the tissue, meanwhile, abscisic acid content is decreasing. It is also a positive effect for the improvement of drought resistance15. The ROS synthesis also encountered in plants that receiving Si application, so that oxidative damage to plants in drought conditions can be avoided. In addition, the application of Si to plants is able to increase the activity of enzymatic antioxidants such as superoxide dismutase (SOD), catalase (CAT), guaiacol peroxidase (GPX), glutathione peroxidase (GSH-Px), ascorbate peroxidase (APX), Glutathione Reductase (GR), dehydroascorbate reductase (DHAR) and monodehydroascorbate reductase (MDHAR). The increase of antioxidant activity capable to decrease oxidative damage caused by ROS, thus increasing drought resistance13,14,16.
The study on some previously paragraphs provide an overview of the potential of B and Si applications as an inducer agent of biochemical resistance of oil palm to drought. Nevertheless, accurate information related to this was still can not be found. Mechanism of action of B and Si as the inducer of biochemical resistance of oil palm to drought also remain unclear. Therefore, the study aimed to determine (1) The level of biochemical resistance to drought stress on oil palm using B and Si applications and (2) The mechanism of B and Si actions in the induction of biochemical resistance of oil palm seedlings to drought stress.
MATERIALS AND METHODS
Planting materials used in the study were 5 months old of oil palm seedlings, ready to be moved from pre-nursery to the main-nursery. Seeds used were uniform in terms of the size of the seeds, healthy and came from the same progeny. Seedlings planted in the polybags, 40×40 cm in size were filled with top soil surface layer. Seeds were arranged at a spacing of 90×90×90 cm with equilateral triangles pattern.
Experiment was arranged in a Randomized Complete Block Design (RCBD) factorial with three blocks as replications. The first factor was the dose of boron (B) and the second factor was the dose of silicon (Si). In this study, the boric acid (H3BO3) was used as a source of B and sodium silicate (Na2SiO3) as a source of Si. The first factor consisted of 6 dose levels, namely 0.00, 0.17, 0.44, 0.87, 1.31 and 1.75 g per seedling while the second factor consisted of 5 levels, namely 0.00, 1.15, 2.31, 3.46 and 4.69 g per seedling. Application of B and Si were done before drought stress treatment until 4 months after transplanting, once a month with the same dose every month depending on the treatment. The applications of B and Si were done by sowing in a circular array about 10 cm from the base of the stem and then covered with soil.
The plants were also fertilized using NPK fertilizer (15:15:15), KCl and kieserit. The NPK fertilizer doses at 1st, 2nd, 3rd and 4th months after transplanting were 24.00, 28.00, 32.00 and 40.00 g per seedling, respectively. The KCl doses at 1st, 2nd, 3rd and 4th months after transplanting were 2.50, 2.83, 3.25 and 4.17 g per seedling, respectively, while the application dose of kieserit at 1st, 2nd, 3rd and 4th months after transplanting were 10.81, 12.69, 13.07 and 3.85 g per seedling, respectively. In addition, the activities of weeding and pest control were done in accordance with field conditions. Before the oil palm seedlings get drought stress treatment, during the first until 4 months after transplanting, seedlings watered regularly every day as much as 1-3 L per polybag.
Testing the level of resistance of oil palm seedlings to drought stress was done as soon as the plants were maintained in ideal conditions for 5 months. Drought stress treatment was started at the beginning of the 6 month after transplanting. During the drought stress treatment, no watering was done until the moisture content of the soil at seedling media reached permanent wilting point. To anticipate the possibility of rain during the test, the seedlings being tested were placed in a plastic house. The oil palm seedlings with the age of 14 months after transplanting, with the planting medium was combine of soil:sand:compost (1:1:1), reached permanent wilting point after 18 days of drought stress treated with soil moisture content of approximately 10%.
Proline compounds analyzed from the leaf tissue, 5 and 6 months after transplanting or before and after drought stress treatment. Samples were in the form of leaves as much as 0.5 g. The leaves were pounded with mortar then put in 10 mL sulfosalisilat acid solution 3%. The solution filtered using Whatman 2 filter paper. A total of 2 mL filtrate was treated with 2 mL ninhydrin acid and 2 mL of glacial acetic acid in a test tube at 100°C for 1 h. The solution was then inserted into a beaker containing ice, added 4 mL of toluene and shaken with a stirrer 15-20 sec. Red toluene containing proline at the top aspirated with a pipette and the solution absorbance read using spectronic 21D at a wavelength of 520 nm17.
Analysis of total phenolic was done after the oil palm seedlings treated with drought treatment. Total phenolic was analyzed using spectrophotometric method. The samples were dry leaves, roasted for 48 h at 40°C. Samples were subsequently smoothed using a leaf grinder and sieved using a 60 mesh size of sieve. A total of 50 mg of sample was introduced into a test tube, added with 0.4 mL of Folin-Ciocalteu and left for 5-8 min. Then, the solution was added with 4 mL of Na2CO3 7% and filtered using Whatman filter paper. The solution was put into the measuring flask and it was added with 10 mL aquabidestilata (total volume of the solution reach up to 10 mL after added with aquabidestilata). After 2 h, the absorbance was read at a wavelength of 765 nm using UV-Vis spectrophotometer (UV-1700 pharma spec). As a blank was used aquabidestilata and Folin-Ciocalteu reagent. Total phenolic expressed as gram equivalents gallic acid per 100 g of dry weight of subfraction (% w/w EAG)18.
The antioxidant activity was measured after the oil palm seedlings treated with drought stress, using DPPH method (2,2-diphenyl-1-picrylydrazyl). A total of 50 mg of oil palm leaves powder that have been smoothed using a grinder coupled with 1 mL of DPPH 0.4 mM and 3.95 mL of ethanol. The mixture was homogenized using vortex and left for 30 min. After that, the absorbance of the solution was measured at a wavelength of 517 nm against the blank (consisting of 50 μL ethanol extract and 4.95 mL of ethanol). The amount of antioxidant activity was determined using the equation18:
Total antioxidant was determined based on IC50 values, namely the concentration of the extract that provides 50% of antioxidant activity when compared to control through the linear line18.
A total of 0.5 g of fresh leaves put into a test tube and homogenized with adding 5 mL of 0.1% (w/v) trichloroacetic acid (TCA). A total of 0.5 mL supernatants put into a test tube and added with 0.5 mL of phosphate buffer 100 mM and 2 mL of potassium iodida (KI) (1 M KI w/v in double-distilled water) and then left for 60 min in the dark room. The content of H2O2 was subsequently determined using spectronic UV-Vis 1700 Shimadzu at a wavelength of 390 nm. As the blank was TCA 1% without supernatant of oil palm leaves. The amount of H2O2 was calculated using a standard curve of known concentration19 of H2O2.
Plant dry weight was measured at 5 and 6 months after transplanting or before and after drought treatment. Dry weight of the plant was separated into several sections, namely roots, stems and leaves. Dry weight obtained by drying the samples using oven at 65°C for 48 h, until the dry weight was constant. The next, samples were weighed using a digital balance.
Height and diameter of oil palm seedling stem were observed once a week, on the beginning of the 1st week after transplanting. The instruments which used were the ruler and digital callipers. Plant height was measured from the base of the stem above the soil surface to the tip of the highest leaf. Meanwhile, the trunk diameter measured at a height of ±2 cm above the soil surface.
The data were analysed using the analysis of variance (ANOVA) at 5% level and followed by Duncan’s Multiple Range Test (DMRT) if there were significant differences among the treatments. The optimum doses of B and Si that were able to induce biochemical resistance of oil palm seedlings to drought stress were determined using regression analysis. Meanwhile, the relationship patterns between the parameters were determined using correlation analysis. All the analysis were performed using the General Linear Model Procedure (PROC GLM)20.
RESULTS
The doses of B applications had quadratic regression relationships with the concentrations of proline in the leaf tissues of oil palm seedlings, before or after drought stress period (Table 1). In the drought stress treatment period, the increase of B applications doses up to 1.12 g per seedling were always followed by the increase of the synthesis of proline in the leaf tissues of oil palm seedlings, so that the concentrations of proline become higher (Fig. 1). However, increasing the doses of B applications after exceeding the 1.12 g per seedling was able to inhibit the rate of proline synthesis in the leaf tissues of oil palm seedlings. So that, the lower concentrations of proline, the higher doses of B (Fig. 1). The doses of B applications that higher than 1.12 g per seedling inhibited the rate of proline synthesis due to the toxicity of B. The toxicity of B was indicated by the appearance of chlorosis and necrosis of leaf tissue which was followed by drying the leaves. Drying leaves that was caused B toxicity occurred due to oil palm seedlings experiencing oxidative stress.
The synthesis rate of proline in leaf tissue increases after oil palm seedlings exposed to drought stress (Fig. 1). The concentration of proline in the leaf tissue of oil palm seedlings after drought stress period was higher than the period before drought stress treatment. The soil’s moisture content in planting medium was drastically decreased due to the impact of drought stress treatment. The condition was responded by oil palm seedlings trough the increase of proline synthesis rate.
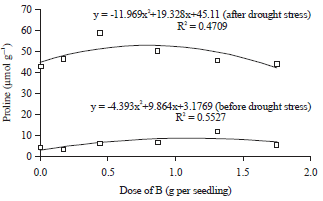 | |
| Fig. 1: | Regression between dose of B and proline |
| Table 1: | Regression between application dose of B and proline |
 | |
| *Regression | |
This mechanism was one of the strategies developed of oil palm seedlings to defend themselves from drought stress through osmotic adjustment mechanism. In such situations, proline compounds act as osmoregulator that prevent the cell organelles from damage due to drought stress. At the period after drought stress, the increase of application B up to 0.81 g per seedling was always followed by an increase proline synthesis rate in the leaf tissues of oil palm seedlings. Increasing doses of B applications after exceeding 0.81 g per seedling could inhibit the proline synthesis rate due to the toxicity of B. The toxicities of B damage the cell membranes and remodeled proline compounds that had been synthesized and disrupted the activities of its synthesis.
The increase in doses of B applications before reaching the optimal dose could increase the proline synthesis rate, before and after the drought period (Fig. 1). The dose application of B which closed to the optimal dose could stimulate the synthesis of glutamate. Glutamate is a member of amino acid that acts as a precursor of proline. One of the functions of B in the metabolic processes of plants is closely related to the metabolism of nitrogen, particularly amino acids. The rate of synthesis of proline compounds may be indicated by the presence of glutamate. The increase of glutamate concentration in the cytoplasm of leaf cell was followed by an increase in the synthesis of proline, thus proline concentration becomes higher.
Sillicon application did not have regression relationship with the concentration of proline in the leaf tissue’s oil palm seedlings (Table 2). This is consistent with some previous research which indicates that the plant response to Si application is not significant when it’s exposed to the ideal environmental conditions.
 | |
| Fig. 2: | Regression between Si application dose and proline, after drought stress |
 | |
| Fig. 3: | Regression between application dose of B and total phenolic compounds, under drought stress |
 | |
| Fig. 4: | Regression between application dose of B and total antioxidant activity, under drought stress |
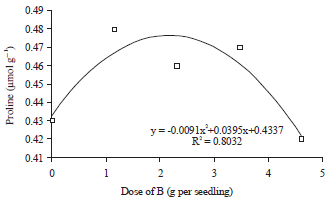
In the period after drought stress, however, doses of Si application has quadratic regression relationship with proline concentration (Fig. 2). The increase of Si applications doses up to 1.15 g per seedling were always followed by the increase in the concentration of proline in the leaf tissue of oil palm seedlings after drought stress.
| Table 2: | Regression between application dose of Si and proline |
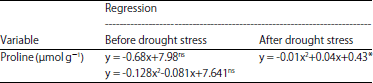 | |
| *Regression, ns: No regression | |
| Table 3: | Regressions between application dose of B with total phenolic compounds, total antioxidant and H2O2 activities, under drought stress condition |
 | |
| *Regression | |
The increase of Si applications doses higher than 1.15 g per seedling inhibited the rate of proline synthesis so that its concentrations decrease in leaf tissues of oil palm seedlings after drought stress.
The applications of Si with the doses higher than 1.15 g per seedling changed the orientation of the amino acid usage, previously used to synthesize glutamate into chlorophyll change to glutamate into enzymes. Glutamate synthesis from the amino acids was not a priority at the time of Si application dose >1.15 g per seedling so that the concentration of proline in leaf tissue decreased. The mechanism of resistance of oil palm seedlings to drought stress changed from the osmotic adjustment mechanisms into strengthening the capacity of photosynthesis through higher chlorophyll and enzyme synthesis in Si application dose >1.15 g per seedling.
On the environmental conditions which were not ideal, for example during drought stress, Si helped plants to avoid oxidative damage on the leaf tissue. Leaf cells especially cell membrane was maintained by Si. It was not damaged during drought stress exposure. The cell membrane was maintained to provide a greater opportunity for the cell to synthesize higher proline compound in the cytoplasm of leaf cell, thus came the osmotic adjustment mechanism. Based on these opinions, the role of Si in the synthesis of proline was indirect through the cell conditioning, so the cells became healthy although exposed to drought stress. The cells had the opportunity to synthesize more proline.
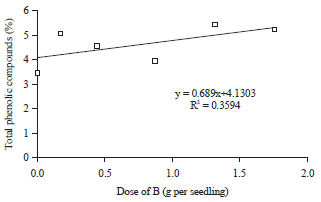
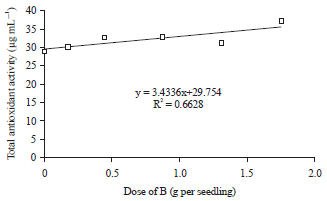
There were linear regression relationships between the dose of B application with concentrations of phenolic compounds, total antioxidant and H2O2 activities in the period of after drought stress treatment (Table 3). The increase of doses B application up to 1.75 g per seedling were always followed by the increase of phenolic compounds concentration (Fig. 3) and total antioxidant activity (Fig. 4), whereas, the H2O2 activity was gradually decreased (Fig. 5) due to the increase of doses B application that applied to the oil palm seedlings after drought stress treatment.
 | |
| Fig. 5: | Regression between application dose of B and H2O2 activity, under drought stress |
 | |
| Fig. 6: | Regression between application dose of Si and total phenolic compounds, under drought stress condition |
| Table 4: | Regressions between application dose of Si with antioxidant activity, total phenolic compounds and H2O2 activity, after drought stress treatment |
 | |
| *Regression | |
These conditions indicated that B was directly involved in the process of total antioxidant and phenolic compounds metabolism. Boron played an important role in the biosynthesis of phenolic compounds and total antioxidant through its function in hormonal regulation of plant tissue. The sufficiency of B in tissues of oil palm seedlings exposed to drought stress stimulated the hormonal activity that lead to the synthesis of phenolic compounds and total antioxidant activity.
Drought stress conditions stimulated the synthesis of free radicals that caused oxidative damage to plant tissues, especially photosynthesis organ. The free radical compounds in the form of Reactive Oxygen Species (ROS), which one of them was H2O2. Hydrogen peroxide was one of the most widely found ROS in plant tissues when exposed to drought stress and stimulated photosynthesis organ to damage. On the other hand, the increase in the synthesis of H2O2 in plant tissue instinctively followed by a rise in the rate of synthesis and activities of total antioxidant and phenolic compounds. Total antioxidant and phenolic compounds were organic compounds which important to suppress the synthesis and inhibit the activity of H2O2, so that oxidative damage of photosynthetic organs on oil palm seedlings could be inhibited. The increase in application doses of B were always followed by the increase in the synthesis of phenolic compounds and total antioxidant activity (Fig. 3, 4). The increase of synthesis rate of phenolic compounds and total antioxidant activity at higher application doses of B would lead to inhibite the synthesis and activity of H2O2 (Fig. 5). Synthesis and activity of H2O2 were also influenced by the concentration of proline in the leaf tissue. Higher concentrations of proline compounds in leaf tissue as a result of the application doses of B (Fig. 1) were able to inhibit the synthesis and activity of H2O2 (Fig. 5). Therefore, the relationship between the doses of B application to the synthesis and activity of H2O2 were not direct but through the activities of proline compounds, total antioxidant and phenolic compounds.
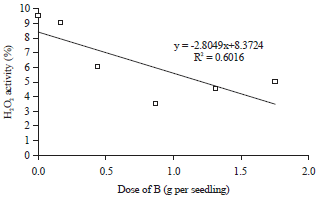
Results of the statistical analysis presented in Table 4 provided information that the doses of Si applications had quadratic regression relationship with the concentration of phenolic compounds, total antioxidant and H2O2 activities. Phenol is an organic compound that is resulted of secondary metabolism that plays an important role as a detoxifying agent of free radical compounds, especially in plants exposed to drought stress. The concentration of phenolic compounds reached a maximum at 0.73 g Si per seedling. The increase of Si applications doses up to 0.73 g per seedling were always followed by the increase in the concentration of phenolic compounds in the leaf tissue of oil palm seedlings which exposed to drought stress (Fig. 6). However, the increase in the doses of Si applications that were higher than 0.73 g per seedling had lower concentrations of phenolic compounds in leaf tissue of oil palm seedlings (Fig. 6). The synthesis rate of phenolic compounds experienced barriers on Si application doses higher than 0.73 g per seedling.
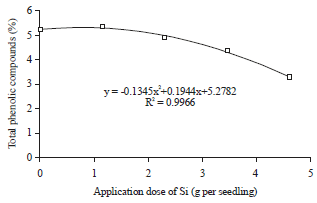
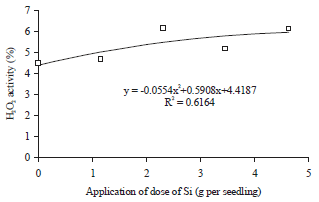
The application doses of Si also affected the total antioxidant activity in leaf tissue of oil palm seedlings at drought stress period. Total antioxidant activity reached a maximum at a dose of Si application by 2.82 g per seedling.
 | |
| Fig. 7: | Regression between application dose of Si and total antioxidant activity, under drought stress |
 | |
| Fig. 8: | Regression between application dose of Si and H2O2 activity, under drought stress |
| Table 5: | Tegressions between applications doses of B with the dry weight of stem, shoot, root, total and root shoot ratio |
 | |
| *Regression, ns: No regression | |
The increase of Si applications doses up to 2.82 g per seedling were always followed by the increase in the total antioxidant activity, while the increase in the Si applications doses after passing the 2.82 g per seedling inhibited the total antioxidant activity (Fig. 7). This condition had a relationship with the concentration of phenolic compounds in leaf tissue. Obviously, some types of active antioxidants in plant tissue are phenolic compounds, especially the non enzymatic antioxidant. The increase in the concentration of phenolic compounds as a result of Si application had possibility to increase the total antioxidant activity. The concentration of phenolic compounds in leaf tissue of oil palm seedlings began to show the downward trend in the application dose of Si >0.73 g per seedling, but the total antioxidant activity of new applications start to decrease at 2.82 g Si per seedling. The decrease in the concentration of phenolic compounds after application dose of Si was >0.73 g per seedling did not necessarily inhibit the total antioxidant activity. The application dose of Si in the range of 0.73-2.82 g per seedling, decreased the antioxidant activity of phenolic compounds. Types of other antioxidant that free from phenolic compounds constituent were still active on applications doses of Si in the range of 0.73-2.82 g per seedling.
Applications doses of Si had quadratic regression relationship with the activity of H2O2 (Fig. 8). The increase of Si applications doses up to 4.29 g per seedling were always followed by the increase in the activity of H2O2, so that the oxidative damage of leaf tissue of oil palm seedlings were increased. The applications doses of Si were >4.29 g per seedling, the increase in applications doses of Si were always followed by decrease in the activity of H2O2. These conditions suggested that the activity of H2O2 could be inhibited by the presence of Si in plant tissues. The applications doses of Si were <4.29 g per seedling were able to maintain the activity of H2O2 in the higher level even at that dose the synthesis of phenolic compounds and total antioxidant activity increase. Although, the concentration of phenolic compounds and total antioxidant activity increased at lower dose of Si but the increase of that materials were not sufficient to withstand the negative effects of H2O2 as indicated by the higher activity of H2O2. The effect of Si application on the inhibition of the activity of H2O2 was direct at higher doses of Si applications. The higher content of Si in the plant tissue of oil palm seedlings affected the biosynthetic process of H2O2, consequently H2O2 activity decreased in the applications doses of Si were >4.29 g per seedling.
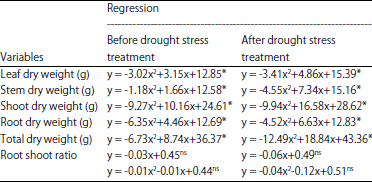
The dry weight of leaves, stems, roots and total oil palm seedlings had quadratic regression relationships with the dose of B application, but did not so with the ratio of the root and shoot (Table 5). Dry weight of leaves, stems, roots and total oil palm seedlings reached maximum at the dose of B applications in the range of 0.35-0.70 g per seedling, before the period of drought stress treatment. Increasing doses of B applications up to >0.70 g per seedling had lower dry weight of leaves, stems, roots and total oil palm seedlings, because the availability of B in the soil exceed the needs of oil palm seedlings. The canopy and root dry weight of wheat decreased in application dose of B less than 6 mg kg–1 soil21.
| Table 6: | Regressions between applications doses of Si with leaf, stem, shoot, root and total dry weight and also root shoot ratio |
 | |
| ns: No regression | |
| Table 7: | Regressions between B applications doses with trunk height and diameter of oil palm seedlings |
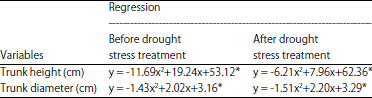 | |
| *Regression | |
| Table 8: | Regressions between applications doses of Si with trunk height and diameter of oil palm seedlings |
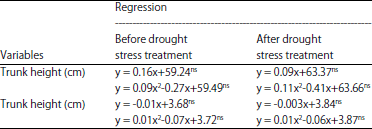 | |
| ns: No regression | |
In the period of drought stress treatment, dry weight of leaves, stems, roots and total oil palm seedlings reached the highest level at 0.71-0.83 g of B per seedling. Increasing doses of B application to >0.83 g per seedling was followed by the decrease of dry weight of leaves, stems, roots and totals due to disruption of plant metabolic activity and tissue damage due to B toxicity.
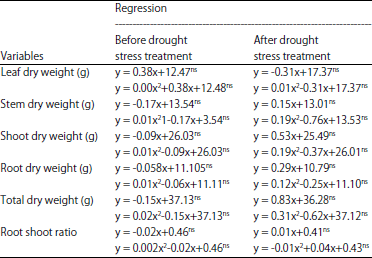
Based on the results of the regression analysis, there was no regression between the dose of Si applications with the dry weight of leaves, stems, roots, total and the ratio of the root shoot of oil palm seedlings, in the period of before and after drought stress treatment (Table 6). The dry weight of all parts of oil palm seedlings was similar to the varying doses of Si up to 4.61 g per seedlings. However, the relationship between the doses of Si application and dry weight of plant parts still could not be determined in this study at the doses of Si application >4.61 g per seedling.
Applications doses of B had quadratic regressions relationships with trunk height and diameter of oil palm seedlings, before and after drought stress period (Table 7). Before the drought stress period, trunk height and diameter of oil palm seedlings reached the maximum at 0.82 and 0.50 g of B per seedlings. The B applications were >0.82 g per seedling caused delays in the increase of trunk height, while trunk diameter of oil palm seedlings began to decline at >0.50 g of B per seedling. In the period of after drought stress condition, trunk height and diameter of oil palm seedlings reached maximum at 0.64 g of B per seedling and 0.73 g of B per seedling. The increase of trunk height and diameter of oil palm seedlings started to inhibit at B applications doses >64 and 0.73 g per seedling, respectively.
The application dose of Si did not have regression relationship with trunk height and diameter of oil palm seedlings, before and after the oil palm seedlings exposed to drought stress (Table 8). Trunk height and diameter of oil palm seedlings were equal among the applications doses of Si, up to 4.61 g of Si per seedling. However, at the applications doses of Si >4.61 g per seedling the pattern of relationships between Si applications doses with trunk height and diameter of oil palm seedlings could not be determined.
DISCUSSION
After drought stress treatment, B applications up to 1.75 g per seedling could increase the antioxidant activity and phenol content, while proline levels increased up to 0.81 g of B per seedling. Decreased levels of proline in the doses of B applications >0.80 g per seedling associated with impaired metabolic activities due to the effects of B toxicity. The increase of proline, phenols and antioxidants in plants were experience to drought could increase the osmotic potential of cells and suppress the activity of free radical compounds22-25. The application of B significantly affect to the concentration of H2O2 in plant tissue26. The H2O2 activity declined consistently in line with the increase in dose application of B up to 1.75 g per seedling. The decrease in H2O2 concentration was related to the presence of phenols, antioxidants and proline, because theoretically these three organic compounds have the ability to detoxify27 H2O2. Nevertheless, no relationship among the proline, phenols and antioxidants were indicated by the absence of correlation in these three variables. Under the drought conditions, proline played a role as a regulator while the phenol and antioxidants were acts as antioxidant components.
Decreased activity of free radical compounds such as H2O2 in plant tissue when exposed to drought stress significantly inhibited the rate of oxidative damage in plants28. Plant organ that was first saved by decreased activity of H2O2 was the inhibition of the aging rate due to the maintenance of leaf chlorophyll content at high level. The main indicators of decrease in the rate of oxidative damage in plant tissues were photosynthetic organs of the plant, especially the leaves. The leaves were still green for long period because of the high content of chlorophyll29,30. As a result, the photosynthetic capacity of the leaf organs still well preserved even when the plants were exposed to drought stress. Indicators of preservation of leaf photosynthetic capacity were the high production of assimilate31-36.
Application of B on oil palm seedlings significantly stimulated the activity of antioxidant that reduces the negative effect of H2O2. These conditions affected the photosynthetic activity directly with the positive effect, because the greenish leaves of oil palm seedlings were maintained for quite long period under drought stress condition. High photosynthetic activity significantly increased the production of assimilates. Assimilates produced subsequently used for plant growth, illustrated by changes in the physical volume in the form of trunk height and diameter. Trunk height and diameter of oil palm seedlings after drought could improve through the applications of B in the range of 0.64 and 0.73 g per seedling. Doses applications of B greater than 0.64-0.73 g per seedling tends to hamper the development of trunk height and diameter of oil palm seedlings, associated with the poison effect of B on oil palm seedlings, because in the toxicity level, B actually interfere the metabolism activity of the plant cell37-40.
Performance of plant morphology which was indicated by the height and diameter of trunk affected the dry weight of oil palm seedlings. Plants with larger physical volume has a greater dry weight as well41-44. Trunk height and diameter were significantly affected by the dose of B application, therefore the dry weight of the plant must also be influenced by the presence of these elements. Plant dry weight increased up to the applications of 0.83 g of B per seedling. There was positive correlation between trunk height and diameter with total dry weight (R2 = 0.67 and R2 = 0.62), so that the increase in trunk height and diameter of oil palm seedlings were always followed by increase in total plant dry weight34,45.
The application of Si to oil palm seedlings has positive influence to the activities of proline, antioxidant and phenol46-48. Levels of proline, phenol and antioxidant activities in plant tissue reach maximum at 1.15, 0.73 and 2.82 g of Si per seedling, respectively. The increase of proline, antioxidants and total phenolic activities in the presence of Si application on a particular dose was indirect.
Oil palm seedlings which were not applied with Si had thinner cell wall and also did not have layer of silica gel on the outer surface of the leaf epidermis, compared with seedlings applied with Si. These conditions result in the decreased of tissue water content of oil palm seedlings without the application of Si49,50. The tissue water content decreased dramatically in the period of drought stress. This major was cause by lower total phenolic, proline and antioxidant activities in oil palm seedlings which were not applied with Si. Proline, phenols and antioxidants were organic compounds that were responsive to drought stress when synthesized51.
The H2O2 activity was increased if the drought stress period lasts longer52-54. The applications of Si up to 4.61 g per seedling tended to increase the activity of H2O2. The increase was associated with decreased levels of phenol, proline and antioxidant activities in Si applications doses of >0.73, >1.15 and >2.82 g per seedling, respectively. There were negative correlations between proline, phenol and antioxidant activities with H2O2 concentration (R2 = -0.62, -0.54 and -0.51). The increased in proline, phenol and antioxidant activities in plant tissues were consistently followed by decrease in the activity of H2O2. The decrease of H2O2 activity was caused by proline, phenols and antioxidants activities in oil palm seedlings applied with Si. The Si application could help the oil palm seedlings to avoid severe damage during drought stress periods.
The increase of oil palm seedlings resistance levels to drought stress were indicated by increase of proline, total phenolic compounds and antioxidant activities and also decrease in the activity of H2O2. However, the increase in the level of resistance was not expressed on dry weight and organ size variables of oil palm seedlings. The Si applications doses up to 4.61 g per seedling have not been able to provide the increase in size of organ or organ dry weight significantly on oil palm seedlings exposed to drought stress.
CONCLUSION
Biochemical resistance level of oil palm seedlings to drought stress may be induced by the applications of B and Si. The application of B was able to induce biochemical resistance significantly on oil palm seedlings to drought stress through the mechanism of accumulations of proline, phenolic compounds and antioxidant activities and also the increase in inhibitory activity of H2O2. On the other hand, the application of Si was able to induce biochemical resistance of oil palm seedlings to drought stress through the mechanism of accumulation of proline and phenolic compounds and also the increase of antioxidant activity and inhibitory H2O2 activity.
ACKNOWLEDGMENTS
This study was funded by LPPM Universitas Gadjah Mada, under the Research University Grant Scheme (Project No. 085/Dir.Keu/KN/DIPA-UGM/2012). The authors would like to express their gratitude to Mr. Widodo and Mrs. Suprihatin Wijayanti, S.P., M.Sc. for their assistance in the study.
REFERENCES
- Henson, I.E. and M.H. Harun, 2007. Short-term responses of oil palm to an interrupted dry season in North Kedah, Malaysia. J. Oil Palm Res., 19: 364-372.
Direct Link - Legros, S., I. Mialet-Serra, J.P. Caliman, F.A. Siregar, A. Clement-Vidal and M. Dingkuhn, 2009. Phenology and growth adjustments of oil palm (Elaeis guineensis) to photoperiod and climate variability. Ann. Bot., 104: 1171-1182.
CrossRefDirect Link - Al-Amin, W. Leal, J.M. de la Trinxeria, A.H. Jaafar and Z. Abdul Ghani, 2011. Assessing the impacts of climate change in the Malaysian agriculture sector and its influences in investment decision. Middle-East J. Scient. Res., 7: 225-234.
Direct Link - Sun, C.X., H.X. Cao, H.B. Shao, X.T. Lei and Y. Xiao, 2011. Growth and physiological responses to water and nutrient stress in oil palm. Afr. J. Biotech., 10: 10465-10471.
Direct Link - Wang, L.F., 2014. Physiological and molecular responses to drought stress in rubber tree (Hevea brasiliensis Muell. Arg.). Plant Physiol. Biochem., 83: 243-249.
CrossRefDirect Link - Ahmed, S., L.H. Akhtar, S. Ahmad, N. Iqbal and M. Nasim, 2009. Cotton (Gossypium hirsutum L.) varieties responded differently to foliar applied boron in terms of quality and yield. Soil Environ., 28: 88-92.
Direct Link - Ananthi, K. and H. Vijayaraghavan, 2012. Soluble protein, nitrate reductase activity and yield responses in cotton genotypes under water stress. Insight Biochem., 2: 1-4.
CrossRefDirect Link - Abbas, S.R., S.D. Ahmad, S.M. Sabir and A.H. Shah, 2014. Detection of drought tolerant sugarcane genotypes (Saccharum officinarum) using lipid peroxidation, antioxidant activity, glycine-betaine and proline contents. J. Soil Sci. Plant Nutr., 14: 233-243.
CrossRefDirect Link - Almeida, G.D., D. Pratissoli, J.C. Zanuncio, V.B. Vicentini, A.M. Holtz and J.E. Serrao, 2009. Calcium silicate and organic mineral fertilizer increase the resistance of tomato plants to Frankliniella schultzei. Phytoparasition, 37: 225-230.
CrossRefDirect Link - Gorecki, R.S. and W. Danielski-Busch, 2009. Effect of silicate fertilizers on yielding of greenhouse cucumber (Cucumis sativus L.) in container cultivation. J. Elementol., 14: 71-78.
Direct Link - Henriet, C., L. Bodarwe, M. Dorel, X. Draye and B. Delvaux, 2008. Leaf silicon content in banana (Musa spp.) reveals the weathering stage of volcanic ash soils in Guadeloupe. Plant Soil, 313: 71-82.
CrossRefDirect Link - Henriet, C., X. Draye, I. Oppitz, R. Swennen and B. Delvaux, 2006. Effects, distribution and uptake of silicon in banana (Musa spp.) under controlled conditions. Plant Soil, 287: 359-374.
CrossRefDirect Link - Ahmad, S.T. and R. Haddad, 2011. Study of silicon effects on antioxidant enzyme activities and osmotic adjustment of wheat under drought stress. Czech J. Genet. Plant Breed., 47: 17-27.
Direct Link - Crusciol, C.A.C., A.L. Pulz, L.B. Lemos, R.P. Soratto and G.P.P. Lima, 2009. Effects of silicon and drought stress on tuber yield and leaf biochemical characteristics in potato. Crop Physiol. Metab., 49: 949-954.
CrossRefDirect Link - Sacala, E., 2009. Role of silicon in plant resistance to water stress. J. Elementol., 14: 619-630.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Alexieva, V., I. Sergiev, S. Mapell and E. Karanov, 2001. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ., 24: 1337-1344.
CrossRefDirect Link - Metwally, A., R. El-Shazoly and A. M. Hamada, 2012. Effect of boron on growth criteria of some wheat cultivars. J. Biol. Earth Sci., 2: B1-B9.
Direct Link - Celik, O. and C. Atak, 2012. The effect of salt stress on antioxidative enzymes and proline content of two Turkish tobacco varieties. Turk. J. Biol., 36: 339-356.
Direct Link - Cvikrova, M., L. Gemperlova, O. Martincova and R. Vankova, 2013. Effect of drought and combined drought and heat stress on polyamine metabolism in proline-over-producing tobacco plants. Plant Physiol. Biochem., 73: 7-15.
CrossRefDirect Link - Hameed, A., M. Goher and N. Iqbal, 2013. Drought induced programmed cell death and associated changes in antioxidants, proteases and lipid peroxidation in wheat leaves. Biol. Plant., 57: 370-374.
CrossRefDirect Link - Fahramand, M., M. Mahmoody, A. Keykha, M. Noori and K. Rigi, 2014. Influence of abiotic stress on proline, photosynthetic enzymes and growth. Int. Res. J. Applied Basic Sci., 8: 257-265.
Direct Link - Hajiboland, R. and F. Farhanghi, 2011. Effect of low boron supply in turnip plants under drought stress. Biologia Plantarum, 55: 775-778.
CrossRefDirect Link - Chakraborty, U. and B. Pradhan, 2012. Oxidative stress in five wheat varieties (Triticum aestivum L.) exposed to water stress and study of their antioxidant enzyme defense system, water stress responsive metabolites and H2O2 accumulation. Braz. J. Plant Physiol., 24: 117-130.
CrossRefDirect Link - Laloi, C., K. Apel and A. Danon, 2004. Reactive oxygen signalling: The latest news. Curr. Opin. Plant Biol., 7: 323-328.
CrossRefDirect Link - Putra, E.T.S., W. Zakaria, N.A.P. Abdullah and G.B. Saleh, 2012. Stomatal morphology, conductance and transpiration of Musa sp. cv. rastali in relation to magnesium, boron and silicon availability. Am. J. Plant Physiol., 7: 84-96.
CrossRefDirect Link - Putra, E.T.S., Issukindarsyah, Taryono and B.H. Purwanto, 2015. Physiological responses of oil palm seedlings to the drought stress using boron and silicon applications. J. Agron., 14: 49-61.
CrossRefDirect Link - Bellaloui, N., 2011. Effect of water stress and foliar boron application on seed protein, oil, fatty acids and nitrogen metabolism in soybean. Am. J. Plant Sci., 2: 692-701.
CrossRefDirect Link - Cao, H.X., C.X. Sun, H.B. Shao, and X.T. Lei, 2011. Effects of low temperature and drought on the physiological and growth changes in oil palm seedlings. Afr. J. Biotechnol., 10: 2630-2637.
Direct Link - Ashraf, M. and P.J.C. Harris, 2013. Photosynthesis under stressful environments: An overview. Photosynthetica, 51: 163-190.
CrossRefDirect Link - Cha-Um, S., N. Yamada, T. Takabe and C. Kirdmanee, 2013. Physiological features and growth characters of oil palm (Elaeis guineensis Jacq.) in response to reduced water-deficit and rewatering. Aust. J. Crop Sci., 7: 432-439.
Direct Link - Ancu, S., E. Chitu, F.C. Marin, I. Ancu and C. Plopa, 2014. Correlation of stomatal conductance with photosynthetic capacity of six walnut cultivars from the national assortment. South West J. Hortic. Biol. Environ., 5: 1-10.
Direct Link - Beig, A.V.G., S.H. Neamati, A. Tehranifar and H. Emami, 2014. Evaluation of chlorophyll fluorescence and biochemical traits of lettuce under drought stress and super absorbent or bentonite application. J. Stress Physiol. Biochem., 10: 301-315.
Direct Link - Hajiboland, R., F. Farhanghi and M. Aliasgharpour, 2012. Morphological and anatomical modifications in leaf, stem and roots of four plant species under boron deficiency conditions. Anales Biologia, 34: 15-29.
CrossRefDirect Link - Demiray, H. and A.E. Dereboylu, 2013. Effects of excess and deficient boron and niacin on the ultrastructure of root cells in Daucus carota cv. Nantes. Turk. J. Bot., 37: 160-166.
CrossRefDirect Link - Gupta, U. and H. Solanki, 2013. Impact of boron deficiency on plant growth. Int. J. Bioassays, 2: 1048-1050.
Direct Link - Putra, E.T.S., Issukindarsyah, Taryono, B.H. Purwanto and D. Indradewa, 2016. Role of boron and silicon in inducing mechanical resistance of oil palm seedlings to drought stress. J. Appl. Sci., 16: 242-251.
CrossRefDirect Link - Putra, E.T.S., W. Zakaria, N.A.P. Abdullah and G. Saleh, 2010. Cell ultrastructure and peel nutrient content of neck zone in six cultivars of Musa sp. fruit during ripening. Int. J. Bot., 6: 47-52.
CrossRefDirect Link - Putra, E.T.S., W. Zakaria, N.A.P. Abdullah and G. Saleh, 2010. Weak neck of Musa sp. cv. Rastali: A review on it's genetic, crop nutrition and post harvest. J. Agron., 9: 45-51.
CrossRefDirect Link - Chaukiyal, S.P. and P. Bhatia, 2014. Effect of water stress on nitrate reductase activity and growth parameters of some Dalbergia sissoo Roxb. clones under glass house condition. Octa J. Environ. Res., 2: 112-120.
Direct Link - Gong, H., X. Zhu, K. Chen, S. Wang and C. Zhang, 2005. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci., 169: 313-321.
CrossRefDirect Link - Bharwana, S.A., S. Ali, M.A. Farooq, N. Iqbal, F. Abbas and M.S.A. Ahmad, 2013. Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. J. Bioremed. Biodegrad., Vol. 4.
CrossRefDirect Link - Biglary, F., R. Haddad, R. Hosseini and A. Sotudehniya, 2011. Roles of silicon in improving oxidative stress resistance by increase of chlorophyll content and relative water content of rice (Oryza sativa L.) genotypes. Proceedings of the 5th International Conference on Silicon in Agriculture, September 13-18, 2011, Beijing, China, pp: 1-9.
- Dastan, S., A.G. Malidarreh and H.R. Mobasser, 2011. Effects of water stress and silicon application on agronomical indices, quantity yield and harvest index in rice (Oryza sativa L.). Proceedings of the 5th International Conference on Silicon in Agriculture, September 13-18, 2011, Beijing, China, pp: 30-31.
- Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Mafakheri, A., A. Siosemardeh, B. Bahramnejad, P.C. Struik and Y. Sohrabi, 2011. Effect of drought stress and subsequent recovery on protein, carbohydrate contents, catalase and peroxidase activities in three chickpea (Cicer arietinum) cultivars. Aust. J. Crop Sci., 5: 1255-1260.
Direct Link - Sekmen, A.H., R. Ozgur, B. Uzilday and I. Turkan, 2014. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot., 99: 141-149.
CrossRefDirect Link - Yang, L., J. Zhang, J. He, Y. Qin and D. Hua et al., 2014. ABA-mediated ROS in mitochondria regulate root meristem activity by controlling PLETHORA expression in arabidopsis. PLoS Genet, Vol. 10.
CrossRefDirect Link