
Kourosh Delavar
Department of Biology, Ashtian Branch, Islamic Azad University, Ashtian, Iran
Shekoofeh Enteshari
Department of Biology, Payame Noor University, Islamic Republic of Iran
Shahnaz Gagoonani
Department of Biology, Payame Noor University, Islamic Republic of Iran
Saeed Kamali
Department of Biology, Ashtian Branch, Islamic Azad University, Ashtian, Iran
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 4 | Page No.: 128-135







ABSTRACT
Background: Silicon has beneficial effects on the plant growth by alleviating of biotic and abiotic stresses. The aimed of the present experiment was the study of the effect of silicon nutrition on the Borago officinalis under aluminum stress condition. Materials and Methods: Interaction of four concentrations of silicon (0, 0.2, 1 and 2 mM) and three concentrations of aluminums (0, 20 and 60 mM) on the biomass and mineral (Ca, K and P) content of Borago officinalis were investigated under hydroponic condition. The experiment was based on a randomized block design with tree replicates. Results: Results of present study clearly demonstrated the alleviation role of Si in borage under Al stress conditions. Also, the results demonstrated the absorption of both aluminum and silicon by Borago officinalis, so that the highest contents of silicon and the aluminum was observed in the Si 0.2 mM and Al 60 mM treatments, respectively. The results showed that the presence of silicon or aluminum in the nutrient solution significantly increased the calcium (Ca), phosphorous (P) and potassium (K) content of plants, especially in the shoots. However, combination of Si with Al significantly decreased the content of all mentioned elements in the borage. On the other hand, Al and Si have an antagonistic interaction, because silicon almost in all treatments reduced the Al content of plant and vice versa. Conclusion: The results of present study suggested that Al and Si have an antagonistic effect on the mineral content of plants and probably silicon by decreasing of Al absorption in borage, reduced its toxicity.
PDF Abstract XML References Citation
Received: February 12, 2016;
Accepted: March 06, 2016;
Published: April 15, 2016
How to cite this article
Kourosh Delavar, Shekoofeh Enteshari, Shahnaz Gagoonani and Saeed Kamali, 2016. Effect of Silicon on the Mineral Content of Borage (Borago officinalis L.) under Aluminum Stress. Journal of Biological Sciences, 16: 128-135.
DOI: 10.3923/jbs.2016.128.135
URL: https://scialert.net/abstract/?doi=jbs.2016.128.135
DOI: 10.3923/jbs.2016.128.135
URL: https://scialert.net/abstract/?doi=jbs.2016.128.135
INTRODUCTION
Since the beginning of the 19th century silicon has been found in significant concentrations in plants1. Silicon, after oxygen is the most abundant element both in the earth’s crust and soil2. Most soils contain about 100-500 μM of silicon as monosalisilic acid3, so any plants that are grown in soil can contain Si in their tissues. Although silicon is not considered as an essential element for plants, it plays an important role in the growth and development of many plant species, especially grasses, such as rice, sugarcane and some cyperaceous2,4,5. In addition, there are several reports about the beneficial effects of silicon in plants under biotic and abiotic stress such as disease, insects, drought, flood, frost, salinity, heavy metals toxicity and nutritional stress4,6. Also, there are several reports about the alleviation roles of silicon on various type of stress in different plants such as barley (Hordeum vulgare)7, corn (Zea mays)5 and sorghum (Sorghum bicolor)8. Silicon impact the plant in various ways including stimulation of plant antioxidant systems, create complex with heavy metals and precipitated them, inactivation of toxic metals in plant growth medium, impact on the absorptions process and compartmentation of heavy metals within the plant cell1.
On the other hand, aluminum acts as a limiting factor for the growth of plants in acidic soils and its toxic effects on plants has been demonstrated9. Absorption of Al in both apoplastic and symplastic paths is very fast10 and therefore it can affect a variety of intercellular and intracellular regions of the cell11. Aluminum ions (Al3+) can bind to the oxygen of macromolecules such as polysaccharides, proteins and nucleic acids and prevents the dividing and developing of cells12. Aluminum ions in acidic environments are more active and therefore have more toxic effects on plant cells13.
Some studies demonstrated the alleviative effect of silicon on the aluminum toxicity13. Several mechanisms have been proposed for this effect. The first mechanism is the interaction of silicon and aluminum in soil and production of subcloids, which decreased phytotoxic aluminum concentration in the soil 13. Also, Kidd et al.14 indicated that silicon may stimulate phenolic exudation by roots and chelated Al and thus Al absorption reduced by the corn roots. Finally some researchers explained that Al can be detoxified by plant mechanisms either by forming hydroxyl aluminum silicates in the root apoplast15,16 or by a sequestration in phytoliths17.
In the present study, the effects of different concentrations of silicon (0, 0.2, 1 and 2 mM) on the biomass and mineral content of borage (Borago officinalis L.), under weak (Allow = 20 mM) and strong (Alhigh = 60 mM) aluminum stress conditions were investigated.
MATERIALS AND METHODS
Borage (Borago officinalis) seeds were obtained from Isfahan Agricultural Research Center in Iran. The seeds were surface sterilized with 5% (w/v) sodium hypochlorite for 15 min, rinsed 4 times with distilled water and then sown in plastic pots (2 kg) containing perlite and kept in greenhouse. The temperature of greenhouse was 26±2°C in day, 18±2°C in night and relative humidity was between 60-80%. Plants irrigated with half-strength long Ashton nutrient solution (pH 5.5)18 and distilled water respectively for 3 weeks. Plants divided in 4 main groups (control, silicon treatment, aluminum treatment and silicon with aluminum treatment). For silicon treatment plants irrigated with silicon that added on long Ashton solution with 4 concentrations (0, 0.2, 1 and 2 mM as Na2SiO3.5H2O). After two weeks, for aluminum treatments, 3 concentrations of AlCl3 [0, 20 mM (Allow) and 60 mM (Alhigh)] were applied at pH 4. Control plants only were irrigated with long Ashton solution. After two week plants were harvested and the fresh weight was measured directly after harvesting and dry weight was measured by drying the plants at 70°C for 48 h to give a constant weight.
The ion content of plants was determined in dry matter by using nitric-perchloric acid (1%) digestion19 and analyzed with inductively coupled plasma optical emission spectrometry (ICP- Optima 7300dv). This experiment was based on a randomized block design with tree replicates. Data from quantitative parameters were analyzed using ANOVA (SAS version 8.1) and means were compared by Duncan’s multiple range tests at the 5% level.
RESULTS
Growth parameters: Results of present study indicated that Si in all applied concentrations significantly increased both fresh and dried weight of plants, however, Al only in 60 mM concentration (Alhigh) significantly decreased biomass of borage compared with control groups (Table 1). On the other hands, the application of Si under Al stress, almost in all treatments, significantly increased fresh and dried weight of borage compared with alone Al treated plants (Table 1).
Aluminum: The results showed that the aluminum content of the shoots, in the high concentration of Al (Alhigh) treatments, significantly increased (8 times) compared with the control groups. On the other hand, the use of different concentrations of silicon in the presence of low concentration of aluminum (Allow) had no effect on the shoot’s Al content, but significantly decreased the Al content of plant under Alhigh treatments.
| Table 1: | Effect of different concentrations of Si and Al on the fresh (FW) and dried (DW) weight of borage |
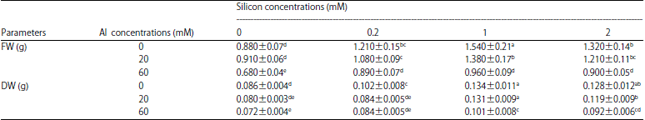 | |
| Different letters indicate significant differences by LSD (p<0.05) | |
In roots, the results showed that the Al content of plants increased in both Alhigh and Allow treatments and added of silicon significantly decreased the Al content of the roots (Fig. 1a, b).
Calcium: The results indicated that the calcium content of the shoots in Alhigh treatments significantly increased compared with both control and Allow treatment. Similarly, Si increased the Ca content of shoots compared with control group. However, application of Si under Alhigh stress condition significantly decreased the Ca content of plants compared with alone Alhigh treated plants (Fig. 1c). A Simllar result was observed in the roots, but in Allow treatments, silicon commonly increases the content of calcium (Fig. 1d).
Potassium: Potassium content of the shoot increased in both Alhigh and all silicon treatments. However, the application of Si 1 and 2 mM decreased the P content of shoots under Al stress condition. Although in plants that treated with silicon 0.2 and 2 Mm shoot’s potassium content was increased under Allow stress conditions (Fig. 2a). Potassium content of roots significantly reduced in plants that treated with silicon 2 mM or Alhigh. Silicon, in both Allow and Alhigh concentration had no significant effect on the potassium content of roots (Fig. 2b).
Phosphorus: Phosphorus content of shoot significantly increased in plants that treated with 1 and 2 mM of silicon or Alhigh. In all other treatments P content had no significant changes (Fig. 2c). On the contrary, different concentrations of silicon and aluminum had no effect on P content in roots (data not shown).
Silicon: The maximum contents of silicon in the shoots of borage was observed in Si 0.2 mM treatments, whether or not presence of aluminum. Apparently aluminum could not prevent the Si absorption by borage in Si 0.2 mM treatments. However, aluminum decreased the silicon content of shoots in Si 1 and 2 mM treatments compared with the control groups (Fig. 2d). The highest contents of silicon in the roots of borage were observed in the Si 0.2 mM treatments, but unlike the shoots, aluminum significantly decreased the silicon contents of roots in mentioned treatments. On the other hand, Al had no effects on the Si content of roots in the other concentrations of silicon (i.e., 1 and 2 mM), in the presence or absence of Al (Fig. 2e).
DISCUSSION
Results of present study clearly demonstrated the alleviation role of Si in borage under Al stress conditions, because the application of Si significantly improved the biomass of borage under Al stress compared with the alone Al treated plants. Alleviative effects of Si on the Al stress also have been previously documented on maize15, soybean13 and rice20. Stimulation of antioxidant systems, complex formation or co-precipitation with Si and immobilization of toxic metal ions in growth media and within the plant have been suggested as mechanisms for Si-mediated alleviation of Al and other heavy metal stresses in higher plants13.
Results showed that the borage can absorbed and transferred silicon to the shoot, but the total amount of silicon in the roots and shoots, at the best case were less than about 0.05 and 0.16% in dry weight, respectively. According to the Marschner21 classification, borage can be classified as Si excluding plants. However, it should be noted that this amount of Si have been absorbed during a 4 week period. Therefore if the plant was exposed to silicon for a longer period, it may have been accumulated the higher amounts of Si.
In the case of aluminum, results demonstrated that a high amount of Al can be absorbed by the roots of borage, especially at the Alhigh, but the little amounts of it can be transferred to the shoots (Fig. 3). Some studies showed that Al rapidly absorbed in the apoplastic and symplastic pathway4,8, therefore, absorbs a large amount of Al, particularly in Alhigh is a common process. Mossor-Pietraszewska12 examining about 400 plant species from 45 different families and determined that the accumulation of Al in plants depends largely on their genetic background.
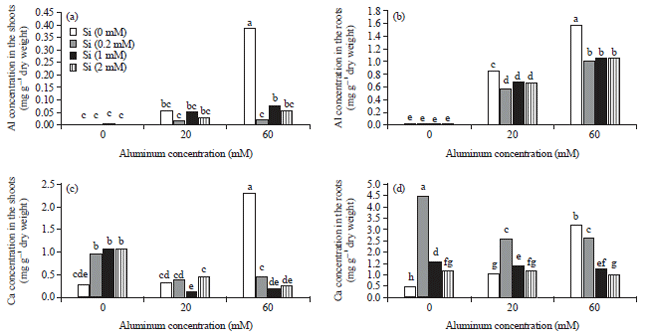 | |
| Fig. 1(a-d): | Effect of different concentrations of silicon and aluminum, on the aluminum content of (a) Shoots, (b) Roots, calcium content of the (c) Shoots and (d) Roots in borage. Different letters indicate significant differences by LSD (p<0.05) |
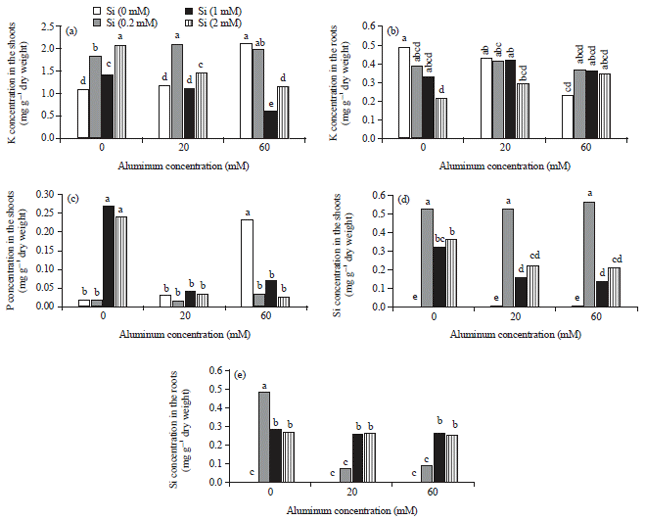 | |
| Fig. 2(a-e): | Effect of different concentrations of silicon and aluminum, on the potassium content of the (a) Shoots, (b) Roots, the phosphorous content of the (c) Shoots and the silicon content of the (d) Shoots and (e) Roots in borage. Different letters indicate significant differences by LSD (p<0.05) |
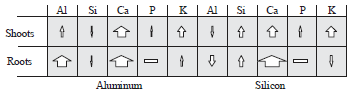 | |
| Fig. 3: | Effect of the silicon and aluminum on the mineral content of shoots and roots in borage. The width of each arrow represents the amounts of increase or decrease of the desired elements compared to control and calculated based on the average content of each elements in Si (0.2, 1 and 2 mM) or aluminum treatments (20 and 60 mM) plants, increase: |
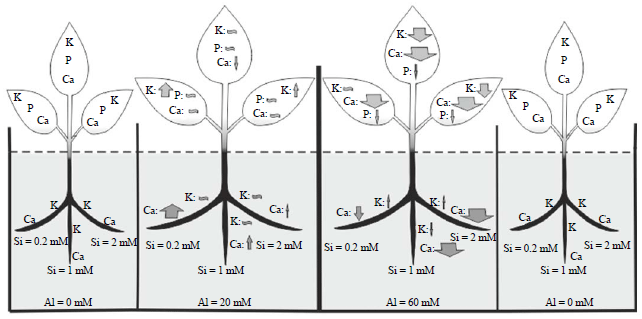 | |
| Fig. 4: | Effect of the interaction between aluminum and silicon on the minerals (phosphorus, calcium and potassium) content of the shoots and roots in borage. The width of each arrow represents the amounts of increase or decrease of the desired elements compared to control, increase: |
On the other hand, Al and Si have an antagonistic interaction, because silicon almost in all treatments (especially at Alhigh, in shoots) reduced the Al content of plant and vice versa (Fig. 3). It may be due to formation of the inactive aluminum hydroxy silicate complex (HAS) that most likely is detoxificated aluminum. The HAS complex formation in apoplast causes the aluminum translocation in a non-toxic form without decreasing of total amount of it15,16,22. But since the formation of this complex is very small, researchers are looking for another way to justify the decreasing of aluminum22. Corrales6 suggested that esterification of cell wall components by silicon, decreased the aluminum binding to the cell wall. While Kidd et al.14 have suggested that Si-induced exudation of phenolics compounds by the roots apex could be potential mechanism in the amelioration of Al toxicity by Si. Phenolic compounds with the chelating aluminum ions, reducing its absorption by the roots. Therefore, results of present study suggested that silicon decreased Al toxicity in borage with reduction its absorption and translocation in plant.
The results showed that the silicon significantly increased the shoots phosphorus content, but had no effects on the roots phosphorus content (Fig. 3). The role of Si in the uptake of phosphorus is one of the earliest known effects of this element. When researchers studied the effect of silicon fertilizer on barley, they have found that silicon fertilizer can increase the phosphorus absorption in the soil, especially in phosphorus deficiency condition4. Also, Eneji et al.23 found a relationship between silicon and phosphorus absorption and released the positive effect of silicon.
Although the results showed that phosphorus content of shoots dramatically increases in Al stressed plants, however, application of Si under Al stress significantly decreased its content (Fig. 3 and 4). Aluminum and phosphorus interactions in plants are very complex and depend on the type of plant and its developmental phase24. The increase of P contents in borage under Al stress condition may be contributed to two types of interaction between aluminum and phosphorus: precipitation-adsorption reaction in apoplast and reactions with various organic compounds in the symplast25. In addition, the simultaneous presence of Si and Al in medium had no effect on the shoots phosphorus content. Apparently these elements were involved in interaction with each other and their effect on phosphorus was eliminated.
Both silicon and aluminum enhanced the translocation of potassium from shoots to roots, but silicon has a more prominent role (Fig. 4). Mali and Aery26 showed that even low concentrations of silicon can increase the potassium uptake from the medium (soil or hydroponic culture) due to activation of H+-ATPaes pumps. Silicon, under low concentration of aluminum (Allow), slightly increased the K content of the shoots, but in the high concentration of aluminum (Alhigh) significantly decreased its content (Fig. 4). Zsoldos et al.27 indicated a significant relationship between aluminum concentration and potassium absorption and transportation in plants shoots and Berger et al.28 also concluded that aluminum reduced potassium uptake in plants. According to the results of present study, silicon had no effect on the potassium content of roots under aluminum stress. These results are in contrary with Liang et al.29 results in barley under salt stress. They indicated that silicon by increasing H+-ATPase activity in membrane, increased the uptake and transport of K+ from roots to aerial parts of the plant. This difference may be due to the different effects of salinity and aluminum, specifically on the concentrations of K+ and Na+ ions. Therefore, as Liang et al.29 showed silicon increased the uptake and transport of K+ from roots to shoots and reduced the uptake and transport of sodium in plant.
The most obvious effect of silicon and aluminum in present study were about the changes in the content of Ca ions in borage. Both Si and Al, almost in all concentrations, significantly increased total calcium content of plants (Fig. 3). These results are inconsistent with the Lindberg and Griffiths30 and Minocha et al.31 findings. They showed that aluminum can reduce the calcium content of plant. Also Lindberg and Strid32 indicated that aluminum at 50 μM, decreased the calcium concentration in the cytoplasm of plant cells. Apparently these effects may be contributed to the blocking of calcium channels1 by Al. Sierra et al.33 also reported that aluminum decreased the calcium content in both shoots and roots. Therefore, results of this study about the effect of aluminum on Ca content of borage are inconsistent with the findings of other investigations and needs further study.
However, the results of present study about the silicon content of plant are consistent with the results of other investigations. Savvas et al.34 showed that the use of silicon in a hydroponic system, increased the calcium content of the leaves. Also Mali and Aery26 reported that applying of silicon fertilizer, improved the absorption of calcium in the wheat and cowpea. On the other hand, the results showed that Si under aluminum stress conditions significantly decreased the calcium content of plants, especially in the shoot and counteracts the effects of aluminum (Fig. 4).
CONCLUSION
Results of present study clearly demonstrated the alleviation role of Si in borage under Al stress conditions. The results also showed that the silicon and aluminum have similar effects on the increases of mineral contents of plant, especially in the shoots. However, Al and Si have an antagonistic effect over each other and each of the two can decreased another contents in the plants. On the contrary, the effect of silicon on mineral contents of shoots under aluminum stress was different from the effect of both alone Si and Al treatments, because the mineral content of plants decreased in combination of Si and Al treatments. The results suggested that the antagonistic effects of Si and Al can countervailed their effects on the mineral contents of plant.
ACKNOWLEDGMENT
The authors of this study thanks Dr. Khabiri, K., Dr Zattali, A. and Mr. Alimoradi, E., for their efforts.
REFERENCES
- Sommer, M., D. Kaczorek, Y. Kuzyakov and J. Breuer, 2006. Silicon pools and fluxes in soils and landscapes-a review. J. Plant Nutr. Soil Sci., 169: 310-329.
CrossRefDirect Link - Guntzer, F., C. Keller and J.D. Meunier, 2012. Benefits of plant silicon for crops: A review. Agron. Sustainable Dev., 32: 201-213.
CrossRefDirect Link - Liang, Y., J.W.C. Wong and L. Wei, 2005. Silicon-mediated enhancement of cadmium tolerance in maize (Zea mays L.) grown in cadmium contaminated soil. Chemosphere, 58: 475-483.
CrossRefDirect Link - Corrales, I., C. Poschenrieder and J. Barcelo, 1997. Influence of silicon pretreatment on aluminium toxicity in maize roots. Plant Soil, 190: 203-209.
CrossRefDirect Link - Hammond, K.E., D.E. Evans and M.J. Hodson, 1995. Aluminium/silicon interactions in barley (Hordeum vulgare L.) seedlings. Plant Soil, 173: 89-95.
CrossRefDirect Link - Galvez, L., R.B. Clark, L.M. Gourley and J.W. Maranville, 1987. Silicon interactions with manganese and aluminum toxicity in sorghum. J. Plant Nutr., 10: 1139-1147.
CrossRefDirect Link - Delhaize, E. and P.R. Ryan, 1995. Aluminum toxicity and tolerance in plants. Plant Physiol., 107: 315-321.
PubMedDirect Link - Vazquez, M.D., C. Poschenrieder, I. Corrales and J. Barcelo, 1999. Change in apoplastic aluminum during the initial growth response to aluminum by roots of a tolerant maize variety. Plant Physiol., 119: 435-444.
CrossRefDirect Link - Jones, D.L., L.V. Kochian and S. Gilroy, 1998. Aluminum induces a decrease in cytosolic calcium concentration in BY-2 tobacco cell cultures. Plant Physiol., 116: 81-89.
CrossRefDirect Link - Mossor-Pietraszewska, T., 2001. Effect of aluminium on plant growth and metabolism. Acta Biochimica Polonica, 48: 673-686.
PubMedDirect Link - Liang, Y., W. Sun, Y.G. Zhu and P. Christie, 2007. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut., 147: 422-428.
CrossRefPubMedDirect Link - Kidd, P.S., M. Llugany, C. Poschenrieder, B. Gunse, J. Barcelo, 2001. The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties of maize (Zea mays L.). J. Exp. Bot., 52: 1339-1352.
CrossRefDirect Link - Wang, Y., A. Stass and W.J. Horst, 2004. Apoplastic binding of aluminum is involved in silicon-induced amelioration of aluminum toxicity in maize. Plant Physiol., 136: 3762-3770.
CrossRefDirect Link - Ryder, M., F. Gerard, D.E. Evans and M.J. Hodson, 2003. The use of root growth and modelling data to investigate amelioration of aluminium toxicity by silicon in Picea abies seedlings. J. Inorg. Biochem., 97: 52-58.
CrossRefDirect Link - Hodson, M.J. and A.G. Sangster, 1993. The interaction between silicon and aluminium in Sorghum bicolor (L.) Moench: Growth analysis and X-ray microanalysis. Ann. Bot., 72: 389-400.
CrossRefDirect Link - Bastias, E.I., M.B. Gonzalez-Moro and C. Gonzalez-Murua, 2004. Zea mays L. amylacea from the Lluta Valley (Arica-Chile) tolerates salinity stress when high levels of boron are available. Plant Soil, 267: 73-84.
CrossRefDirect Link - Singh, V.P., D.K. Tripathi, D. Kumar and D.K. Chauhan, 2011. Influence of exogenous silicon addition on aluminium tolerance in rice seedlings. Biol. Trace Elem. Res., 144: 1260-1274.
CrossRefDirect Link - Cocker, K.M., D.E. Evans and M.J. Hodson, 1998. The amelioration of aluminium toxicity by silicon in wheat (Triticum aestivum L.): Malate exudation as evidence for an in planta mechanism. Planta, 204: 318-323.
CrossRefDirect Link - Eneji, E., S. Inanaga, S. Muranaka, J. Li, P. An, T. Hattori and W. Tsuji, 2005. Effect of calcium silicate on growth and dry matter yield of Chloris gayana and Sorghum sudanense under two soil water regimes. Grass Forage Sci., 60: 393-398.
CrossRefDirect Link - Amaizah, N.R., D. Cakmak, E. Saljnikov, G. Roglic, N. Kokovic and D. Manojlovic, 2013. Effect of waste Al-phosphate on soil and plant. Plant Soil Environ., 59: 130-135.
Direct Link - Clarkson, D.T., 1966. Effect of aluminum on the uptake and metabolism of phosphorus by barley seedlings. Plant Physiol., 41: 165-172.
CrossRefDirect Link - Mali, M. and N.C. Aery, 2008. Influence of silicon on growth, relative water contents and uptake of silicon, calcium and potassium in wheat grown in nutrient solution. J. Plant Nutr., 31: 1867-1876.
CrossRefDirect Link - Zsoldos, F., A. Vashegyi, L. Bona, A. Pecsvaradi and Z. Szegletes, 2000. Growth of and potassium transport in winter wheat and durum wheat as affected by various aluminum exposure times. J. Plant Nutr., 23: 913-926.
CrossRefDirect Link - Berger, T.W., C. Eagar, G.E. Likens and G. Stingeder, 2001. Effects of calcium and aluminum chloride additions on foliar and throughfall chemistry in sugar maples. For. Ecol. Manage., 149: 75-90.
CrossRefDirect Link - Liang, Y., W. Zhang, Q. Chen, Y. Liu and R. Ding, 2006. Effect of exogenous silicon (Si) on H+-ATPase activity, phospholipids and fluidity of plasma membrane in leaves of salt-stressed barley (Hordeum vulgare L.). Environ. Exp. Bot., 57: 212-219.
CrossRefDirect Link - Lindberg, S. and G. Griffiths, 1993. Aluminium effects on ATPase activity and lipid composition of plasma membranes in sugar beet roots. J. Exp. Bot., 44: 1543-1550.
CrossRefDirect Link - Minocha, R., S.C. Minocha, S.L. Long and W.C. Shortle, 1992. Effects of aluminum on DNA synthesis, cellular polyamines, polyamine biosynthetic enzymes and inorganic ions in cell suspension cultures of a woody plant, Catharanthus roseus. Physiologia Plantarum, 85: 417-424.
CrossRefDirect Link - Lindberg, S. and H. Strid, 1997. Aluminium induces rapid changes in cytosolic pH and free calcium and potassium concentrations in root protoplasts of wheat (Triticum aestivum). Physiologia Plantarum, 99: 405-414.
CrossRefDirect Link - Sierra, J., H. Ozier-Lafontaine, L. Dufour, A. Meunier, R. Bonhomme and C. Welcker, 2006. Nutrient and assimilate partitioning in two tropical maize cultivars in relation to their tolerance to soil acidity. Field Crops Res., 95: 234-249.
CrossRefDirect Link - Savvas, D., G. Manos, A. Kotsiras and S. Souvaliotis, 2002. Effects of silicon and nutrient-induced salinity on yield, flower quality and nutrient uptake of gerbera grown in a closed hydroponic system. J. Applied Bot., 76: 153-158.
Direct Link