
E. Knopf
Center of Life and Food Sciences Weihenstephan, Technische Universit�t M�nchen (TUM), Microbiology/Ecophysiology of Plants, Hans-Carl-von Carlowitz-Platz 2, 85354, Germany
H. Blaschke
Center of Life and Food Sciences Weihenstephan, Technische Universit�t M�nchen (TUM), Microbiology/Ecophysiology of Plants, Hans-Carl-von Carlowitz-Platz 2, 85354, Germany
J.C. Munch
Institute for Soil Ecology, Helmholtz Zentrum M�nchen, Neuherberg, 85764, Germany
G. Rambold
Mycology University of Bayreuth, Universit�tsstrasse 30, Bayreuth, 95447, Germany
A. Murage
Department of Agricultural Enonomics ICIPE, Kenya, Box 34 Mbita/Homabay/Agricultural Research Institute, Private Bag, Njoro, Kenya
P. Kirika
Department of Botany, National Museums of Kenya, P.O. Box 45166, Nairobi, 00100, Kenya
S. Okaka
College of Biological and Physical Sciences, University of Nairobi, P.O. Box 30197, Nairobi, 00100, Kenya
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 1-2 | Page No.: 12-21







ABSTRACT
Depending on pressure on land, mineral turnover may not be able accommodate increased plant nutritional needs. Arbuscular Mycorrhizal Fungi (AMF) and symbiotic microbial consortia such as diazotrophs may improve and sustain plant production in Lake Victoria Basin (LVB), however soil factor may be bottlenecks to successful application. A biotest was conducted on the three rhizospheric soil types representative of the LVB. Plants established in vert+hist soils showed a rather superior germination rate of 62.5%, compared to loamy Oxisols at 33% and Alfisols with 58%. Moreover , vert+hist soils sustained a very high mycorrhizal colonization (>90% full cortex arbuscle occupancy) compared to its archetypes. Since vert+hist soils registered higher K content in soil chemical analysis at 42 per K (K2O5-CAL m) mg/100 g content compared to oxisols (11) and alfisols (33). Independent experiment with K+ salt and cultured inoculum AMF using pure vermiculite set under strict growth conditions revealed K influence on plant performance at symbiosis. The results revealed that although a major component of the organic N and P cycles in symbiosis influence plant growth, the missing loop K+ could be adjusted by adding a little K+ salt in eco-sensitive low-input agriculture.
PDF Abstract XML References Citation
Received: October 02, 2015;
Accepted: December 09, 2015;
Published: January 15, 2016
How to cite this article
E. Knopf, H. Blaschke, J.C. Munch, G. Rambold, A. Murage, P. Kirika and S. Okaka, 2016. Impacts of Soil on Arbuscular Mycorrhizal Fungi: Growth Responses of Moringa spp., Plants Sampled from Lake Victoria Basin. Journal of Biological Sciences, 16: 12-21.
DOI: 10.3923/jbs.2016.12.21
URL: https://scialert.net/abstract/?doi=jbs.2016.12.21
DOI: 10.3923/jbs.2016.12.21
URL: https://scialert.net/abstract/?doi=jbs.2016.12.21
INTRODUCTION
Soil is a biogeochemical transformation field, food bank to rhizospheric life with regulation functions important to terrestrial gas exchange activities. Approximately 17 elements occur in the soils viz., macro/micronutrients (N, P, K, Ca, Mg and S/Fe, Zn, Mn, B, Cu, Mo, Ni, Al, Na and Cl) and gases (CO2, O2 and H2O) in higher plants (Marschner, 1995). Other elements such as Cu, Cr, Pb, Cd are not so useful to plants but rather exacerbate heavy metal accumulation and contamination in soils. The dynamics, distribution and availability of N and P are ascribed to the biogeochemical cycles in the soil matrix. The process is known to release chelating compounds such as siderophores (metabolic byproducts of fungus) and acidifying molecules by plants that help in the in mineralization processes. However, when mineral turnover does not meet soil nutrient deficit, fertilizer in-put is required to achieve the so-called "critical" nutrient level (Pilon-Smits et al., 2009; White and Brown, 2010) for efficient plant growth. Mineral deficiency stunts growth and excess in soils is toxic to plants, soil fungi population and biodiversity.
Nitrogen mostly occurs in ammonium (NH4+) and nitrate (NO3‾) forms, bonded in Isotopes N15 at 0.3663% and N14 at ≥99.6337%, respectively in the soils. Levels below these indicate a depleted state, while above the mark, indicates N enrichment in the soil. Nitrogen affects the absorption and distribution of practically all other elements and works in cohesion with other macronutrients effectively in promoting plant growth. Phosphorus (P2O5) promotes growth and stimulates development of floral and fruit buds. Phosphorus deficiency affects organogenesis and biomass partitioning in favour of the root system, affecting root:shoot ratio (Poorter and Nagel, 2000) in plants. Phosphor starved indicates a reduction of plant growth and yield (Snyder and Stewart, 2003) and in excess, toxic (Romera et al., 1992). Potassium activates plant metabolism in protein synthesis, photosynthetic band-mediating osmoregulation in stomatal movements and tropisms, effecting deeper and larger root systems through cell development (Marschner, 1995; Silva, 2004).
Approximately 63% of total N from anthropogenic sources recurring in radio-/photo and biological organic N in mineral N in NO3‾ and NH4+ forms are important to plants, but may impact the environment negatively (Dobermann, 2005). The gaseous state of N (NOx, NH3 and N2O) are chemically active in the troposphere with greenhouse effect (Galloway et al., 2008), that could exacerbate acid rains, global warming and other negative environmental impacts. Co-inoculation studies with Plant Growth Promoting Rhizobacter (PGPR) and rhizobia have shown increased plant nodulation and N fixation (Figueiredo et al., 2008; Knopf et al., 2014) which may reduce chemical application in vulnerable freshwater ecosystems like Lake Victoria Basin (LVB).
Arbuscular Mycorrhiza Fungi (AMF) glomeromycetes are obligate biotrophs forming arbuscles which transfer phosphorus to the root system in exchange of photosynthates in P deficient soils in symbiosis. Benefits of AMF are several. Johnson et al. (2004) reported that mycorrhizal fungi assist in modification of functional abilities of soil microflora, mitigating ecological stresses by raising soil pH at acidity, increasing soil moisture levels and regulating temperature. Auge et al. (2001) recorded symbiotic fungal ability to fight pathogens. Hildebrandt et al. (1999) and Medina and Azcon (2010) showed that these fungi might remediate soils via soil enzyme activities, by biodegrading organic compounds passing through soil profile (Schutz et al., 2010), bind Ni, thereby influencing heavy metal uptake by plants, while the sequestered elements in mycorrhizal fungi for example, may reflect a polluted environmental condition, acting as a bio-indicator, an important tool in agriculture (Orlowska et al., 2011). Expansion in agriculture, deforestation, soil degradation, erosion and several other factors have led to current challenges in Lake Victoria basin as depicted by microbial diversity loss. Over 30 million people resides within Lake Victoria, distributed at 297, 97 and 635 persons per kilometer square, respectively (Odada et al., 2009). This has led to deforestation, expansion in agriculture, soil degradation and erosion, interfering with biodiversity. Deforestation increases desertification and ecosystem degradation, exposing the soils to direct solar radiation and higher soil temperature which may alter soil biology and reduce microbial activities (De Deyn et al., 2011). Likewise, sporulation and reproduction of AMF are directly affected by seasonal dynamics, confirmed by large seasonal fluctuations in number of spores of fungi (Giovannetti, 1985).
Moringa (class Moringacea; genera Rosidae) is a monotypic tropical forest species; M. stenopetala is endemic to Kenya, Ethiopia and Somalia while M. oleifera is indigenous to the sub-Himalayan tracts of north-west India, Asia. Moringa is a fast-growing deciduous tropical species that can sustain environment in simplicity when established either at mixed-/or agroforestry levels. Moringa grows well in most tropical climates ranging from arid/semi arid regions to moist/humid, monsoon types. Ideal average annual rainfall reported so far is between 250 and 3000 mm. Moringa grows in altitudes from 600-1200 m but can survive at 2000 m above sea level and outlive light frosts (Reyes, 2006). Investigations show growing temperature of Moringacea range from 25-40°C, performing well in nearly all soil types, with pH range between 5-9, but prefers slightly acidic soils.
Moringa is beneficial in areas of industrial oil production, medicine and domestic use such as multivitamins of high nutritional values, minerals (De Saint-Sauyeur, 1991; Ayssiwede et al., 2011) and in soil sustainability. The plant growth hormones present in Moringa can increase yields by 25-30% for nearly any crop (Foidl et al., 2001). The pterygospermin compound in Moringa enhances plant growth (Foidl et al., 2001; Ayssiwede et al., 2011). The lignocellulose enzymes in plant are useful chemical filters in soil remediation. Optimal use of Moringa and mycorrhizal fungi may be excellent partners at improving soil health status and enhancing plant growth in most eco-sensitive biomes.
MATERIALS AND METHODS
Soil sampling, treatment and storage: The rhizospheric soils were sampled from three regions representative of Lake Victoria Basin (LVB), Kenya with altitudinal stretch of 800, 1300 and 1800 m above sea level and standard/control substrate of German origin "Biotopf®/Pikiererde." The vert-/+histosols were cored from 00°42'00"N and 37°22'00"E location within a rice growing district. Clay alfisols were sampled from a commercial sugar plantation area, at 00°05'04.77"S and 35°07'57.51"E location, 1300 m above sea level. The third soil type (acidic-red oxisol) was collected from humid highlands of Kisii district with high rainfall, approximately 1800 m above sea-level. The fields are frequently tilled using multiple techniques. Soil coring points were in the arable lands mostly used for annual crops.
| Table 1: | Chemical characteristics of native soil types sampled from Lake Victoria basin soils and standard substrate* |
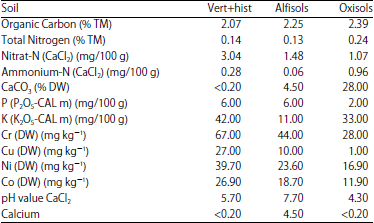 | |
*Standard soil with pH (CaCl2):5, 5, KCl in g L–1 1, 1 solutes 150 mg L–1 N, (CaCl2) 150 mg L–1 P2O5, phosphate (CAL): 210 mg L–1 K2, potassium oxide (CAL) organic substance: 85%, DW: Dry weight | |
The rhizospheric soils were air-dried in the shade for 14 days, organic matter removed, pulverized at 2 mm size and analysed using VDLUFA MB I, D2.1 (Fingerprobe) A 5.1.1 (pH-Wert) and A 6.2.1.1 CAL-Methods, version 2002 (Table 1).
Seed preparation and planting: Seeds from Kenya were acquired from Orongo village within co-ordinates (UTM) 699, 285.37 S and 9,986,154.13 E; 34°45'/0°00': 0°00'/35°00'; 34°45'/0 15': 0°15'/35°00' and Kenya Forestry Research Institute (KEFRI), Maseno within LVB. The first set of germplasm was developed from moringa seeds grown in soils of vert+histosol origin. The second set had acidic loam oxisol background. The seeds were dehulled and the nuts were soaked in sterile water for three days at room temperature to accelerate germination with frequent water changes to avoid fungal contamination. The seeds were planted in vermiculite initially and transplanted into different soil types according to treatments. These blocks were set on a greenhouse bench with mean temperature of 20°C under ambient light supply. These were left to develop for two weeks before inoculation.
Inoculum development: Cultured AMF propagules, Glomus hoi, of University of York origin, registered at BEG, (BEG 104), updated as "Revised sequences from UY110 (accession numbers KC182044 and KC182045) and BEG104 (KC182048-KC182048) cluster within the G. mocrocarpum clade". Glomus mosseae, (currently classified as Funelliformis mosseae, BEG 68, Glomus intraradices (renamed Rhizophagus irregularis) BEG 31, in cocktail form was used to develop an AMF inoculum bank using Plantago major from open sources.
Plantago seeds were left to germinate in vermiculite for about 36 h. The germinated seedlings were transplanted into 5×5×5 cm holes of 650 mL plastic containers filled with autoclaved sand of 0.5-1 mm size, collected from Kronthaler Kieswerk, Freising, Germany. The sand was washed several times and repeatedly in sterile water. Using 2 g of inoculum, a total of 15 pots were established for inoculant banks. These were irrigated daily using Hoagland and Arnon (1950) nutrient solution in 1/10 strength.
Biotest and inoculation of moringa: A biotest was done using 65 mL multi-pots with native soils. Seeds of wild Plantago major were cleaned, dried and planted in 17×24×5 replicated five times. The seeds were sown directly into the native soil. The blocks were left to establish in a greenhouse. After 5 days of germination, images of Plantago major were taken to assess germination rates and plant performance. The colonized roots were harvested after two months of growth for mycorrhizal assays while, the rest served as inoculants.
K probes: A total of 195 rootrainers® of volume 55 cm3 were arranged in trays and divided into three blocks of 65 plantlets each. These were filled with vermiculite growth substrate. Moringa seedlings were directly planted in the substrate and left to germinate under a temperature of 23°C and a humidity of 70%. The seedlings were inoculated with AMF and NFB. After two weeks of germination K+treatment was applied. The seedlings were irrigated with 2 mL of KCl salt solution for further two weeks.
Moringa seedling establishment in native soils and inoculation: Flexible rootrainers® were used for seedling germination using vermiculite or standard (Biotopferde®) as growth media before transplant. Moringa seeds were planted in 55 cm3 sized rootrainers in three blocks with 65 plantlets in each block using vermiculite for pregermination. After two weeks of growth, the seedlings were transplanted into the native soils, divided into four blocks according to soil types and transferred into greenhouse. The seedlings were inoculated with AMF and NFB in rhizobia.
Block 1→AMF inoculum, block 2→AMF+ NFB, block 3→control, without inoculum. In all experiments Plantago major culture and chick-pea rhizobia (Rhizobium sp.) of Cicer arietum legume were used to generate inoculum banks for Moringa inoculation.
Irrigation was done via water pipe sprinkler. Soil moisture content was assessed using time domain reflectometry (TDR; TEKTRONIX sensor, 1502 C/Tektronix Oregon, United States). A 12 cm head with two pins was installed in each root compartment for soil moisture content reading.
Harvest: Moringa plants were fully harvested at the end of the study period although in situ data collection of Basal Stem Diameter (BSD), plant height and fine roots (for mycorrhizal assay quantification; 2-6 months intervals) was done at intervals of two months over the period 2010-2011. In all experiments, measurements on plant height, BSD and biomass were taken. Basal stem diameter and plant height were taken at two weeks intervals for a minimum of two months in each case.
Identification and quantification of mycorrhiza: The roots of Moringa established in different soils were harvested and washed carefully in running water. Root segments were soaked in potassium hydroxide (KOH) 10% at room temperatures (18-22°C). Potassium hydroxide solution was removed and HCl applied for 1-2 h to depigment the dark colored root samples using 3 mL of NH4OH to 30 mL of 10% H2O2 and 567 mL of tap water (Brundrett, 2009). The cleared roots were incubated in ink and vinegar according to Vierheilig et al. (1998) for 12–24 h at room temperature. The stained segments were mounted on slides for light microscopy using Leitz Aristoplan® microscope (Wetzlar, Germany) connected to KAPPA® digital camera. Morphological criteria (Brundrett, 2009) were used to define anatomical key mycorrhizal features, using amount of cells and intercellular spaces in the cortex containing hyphae following, Trouvelot et al. (1986) for estimation.
Statistics: Statistical tests were done using GenStat 9th Edition for windows (VSN) International Ltd., UK). Linear regression model was applied in a two-way analysis of variance (ANOVA) repeated measures with standardized response fitted value residual leverage. The predictions have been formed only for those combinations of factor levels for which means can be estimated without involving aliased parameters. Predictions from regression model based on soil and inoculum effects on basal stem diameter, height increments was applied for consistency, to ascertain if there were significant differences between soils and treatments. Differences were considered significant if p<0.05.
RESULTS
Soil response: A higher plant germination (Σplant/pot-1) rate, ranging between 4.5-29.5% in vert+hist soils compared to those established in loamy oxisols and clay alfisols was observed. This was in harmony with dry weight biomass records, showing higher values in vert+hist, while the standard soil "Biotopferde® showed ≥90% rate in germination. Two-way ANOVA test on soil factor and inoculum influence displayed significant differences on BSD and height increment of both moringa species at p<0.001. The analysis indicated that growth rates and vigor of moringa plants depended much on soil chemical properties. The AMF inoculated moringa seedlings also displayed vigor and faster growth compared to non-inoculated ones in both species, exhibiting higher Basal Stem Diameter (BSD) and plant heights (Fig. 1).
Faster growth and plant vigor was demonstrated in the beginning of growth period on the two Moringa spp. Leafy biomass were maintained longer in AMF+NFB than in single AMF inoculated samples and the non-inoculated blocks. In all blocks, apart from the depletion phase with the outcrop of other weedy competitors, inoculated Moringa seedlings had relatively larger mean BSD and greater heights. However standard soils had minimal colonization apart from runner hyphae and <1% arbuscle formation at the end of the experiment on moringa species.
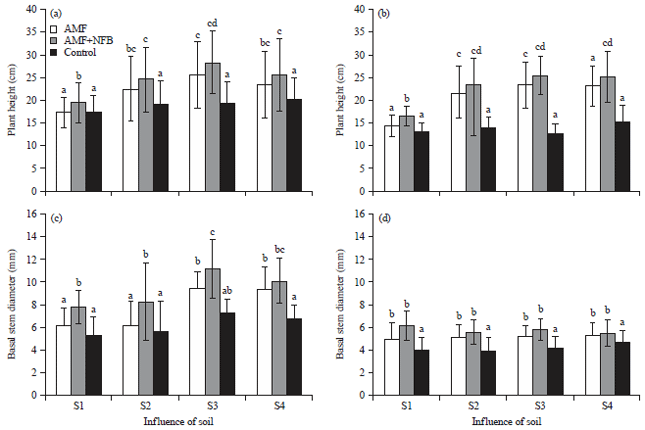 | |
| Fig. 1(a-d): | Influence of soil (S1-S4) on basal stem diameter and height of Moringa stenopetala and Moringa olefera seedings, grwon under ambient light in greenhouse. Soil type, S1: Loamy oxisols, S2: Clay alfisols, S3: Vertisols.histosols and S4: Standard soil, "Biotoferde®" Treatment: Arbuscular Mycorrhizal Fungi (AMF) and Nitrogen Fixing Bacteria (NFB) in hickpea rhizobia. Box range 25-86th and 50th percentile median, whiskers indicate highest and lowest values (Significant α = 0.05) |
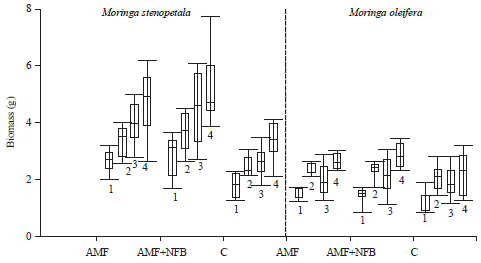 | |
| Fig. 2: | Soil influence on Moringa stenopetala and Moringa oleifera seedlings growth established under ambietn light in greenhouse. Soil types, 1 Oxisols, 2: Alfisols, 3: Vert; histosols) and 4: As a standard sunstrate "Biotopferde®". Treatments: Arbuscular mycorrhizal fungi (AMF+nitrogen fixing bacteria in chickpea rhizobia. Box range 25-75th and 50th percentile median. Whiskers indicate highest and lowest values (Significant α = 0.05) |
Samples established in vert+hist soils or standard soils survived this phase better than those in other native soils, which had mostly defoliated shoots. Non-inoculated samples desiccated with no regeneration. Considering total dry weight biomass distribution (Fig. 2), treatments influenced growth of M. stenopetala unlike M. oleifera.
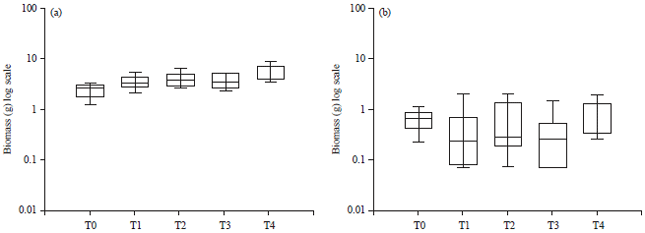 | |
| Fig. 3(a-b): | Boz plot of total dry weight boimass of (a) Moringa stenopetala and (b) Moringa olefera with treatment levels, from T1-T4; AMF, AMF+NFB, AMF+K, AMF+NFB+K and T0 as control. Box range 25-75th and 50th percentile median. Whiskers indicate highest and lowest values (Significatnt = α = 0.05) |
Co-inoculation (AMF+NFB) was again gave better results than single or non-inoculated seedlings with higher dry weight biomass records. Arbuscular mycorrhizal fungi+NFB combination likewise induced larger tubers compared to control samples. Leafy biomass did not respond to inoculum. Moringa stenopetala seedlings seemed to respond more favourably to inoculum compared to M. oleifera in all parameters. However, AMF and NFB combined treatments in both species gave better performance range.
From soil chemical analyses, vert+hist type of soil registered 42 per K (K2O5-CAL m) mg/100 g content as compared to oxisols (11) and alfisols (33). Therefore, K+ and mycorrhizal factor could be of significance in plant performance differences, warranting further K specific tests set under strict growth conditions of 70% humidity levels and temperature of 23°C using vermiculite as a substrate. AMF+K+NFB combination promoted larger BSD compared to other treatments in a row (Fig. 3). Moringa oleifera showed no improvement at K addition, in contrast to M. stenopetala. Improved BSD and height increments in M. stenopetala and M. oleifera revealed positive plant responses in the presence of K+ element in combined treatments. Most treatments caused slight improvements in height and BSD values in M. stenopetala, no significant differences were recorded on M. oleifera species. With K addition, a reasonably larger root diameter was evident.
Rootlets sampled from vert+histsols displayed full arbuscle occupation (>90%) in the root cortex in mycorrhizal quantification. The presence of morphological structures e.g., intracellular hyphal coils, intercellular hyphae, frequent vesicles, arbuscles, ornamented septate freelances in colonization networks were observed mostly from vert+hist types of soil. Highest fine root production was registered among samples established in standard soils. Contrarily Oxisols showed higher growth rates in root production although with very low mycorrhizal colonization. A higher root production was realized at inoculum in-put in standard samples but contrarily very limited colonization. A slightly higher mean diameter value was reported in the fine root calculations. The average fine root biomass (without mycorrhiza) of the fine rootlets inside the control subsets was lower than mycorrhized rootlets in the experiment. Vert+hist soils exhibited regeneration potential close to standard soil with diameter limits falling within the same range. Mean height and diameter increment variations were time-dependant. Negated growth experience at the second sampling season showed less inoculum response. In most cases mean differences were smaller across treatments; while dual (AMF+NFB) inoculation improved plant performance compared to single AMF inoculation.
DISCUSSION
Soil Organic Matter (SOM) is basically defined by stable, natural or inherent features related to soil forming factors and the loss of SOM results in a reduction in soil quality (Larson and Pierce, 1991; Carter, 2002). The soil material retains nutrients by effecting cation and anion-exchange, increasing water holding capacity, improving infiltration rates and enabling aggregate stability in soils (Jones et al., 2005; Barancikova et al., 2010). At decomposition they create benign habitats for micro-organisms, protists, fungi (Unterseher et al., 2008) facilitating biogeochemical cycles. With SOM loss from soils, increased microbial diversity loss consequently occurs. At this stage majority of soils require mineral restocking either though fertilizers or biological techniques such as AMF inoculum. Wagg et al. (2011) studied competitive interactions between Lolium multiflorum and Trifolium pratense in low and high diversity AMF treatments, their report indicated that the higher AMF diversity, the higher the experimental plant biomass. The standard (99.1% organic matter) soils in the experiments with moringa, however, revealed higher growth irrespective of inoculum strength. The native soils (vert+hist) only showed the Glomus tenuis genotypes and high population of the dark septate endophytes, but still ranking high among the rhizospheric soil types in biomass output. Glomus tenuis exists mostly in low phosphorous situations confirming our finding that although through removal of organic matter, the survival rate of species in the soils is still reliable in areas with extreme growth conditions. The impact of AMF inoculum on plant biomass varied with soil type dependent on species, revealing high soil impacts, based on fungal activities. However autochthony (Knopf et al., 2013) seemed to be a stronger variable in influencing positive growth of plants especially M. stenopetala indeginous to Lake Victoria, Kenya planted on its original soil habitat.
Gianinazzi and Schuepp (2002), Ouahmane et al. (2007) and Medina and Azcon (2010) have reported on the potential of mycorrhizal activities in improving degraded soils at aridity. In this study, plants established in the LVB rhizospheric soils with mycorrhizal abundance or inoculated experimental standard substrate on moringa had higher plant performance in comparison to non-inoculated ones, supporting these studies. Faster growth was confirmed to be dependant on soil microbial population, a possible consequence of increased nutrient bioavailability. Loamy oxisols originating from heavy agricultural background in arable lands within LVB had minimal degree of colonization. Medium and low agricultural tillage background soil categories exhibited more diverse mycorrhizal structure formations, indicating AMF richness. It has been reported that cultivated soils destroy AMF functions (Wright et al., 2007; Alguacil et al., 2008). Root cortex photomicrographs revealed high mycorrhization in vert+hist compared to clay alfisols and loam oxisols. This report supports Kengara et al. (2010), where by using the same soil, higher mineralization and degradation of hexachlorobenzene (HCB) by "a community of microbials" was achieved, mycorrhiza (Glomus tenuis) probably being the main driver to successful aerobic processes in their experiment. The AMF taxa mostly of Glomus tenuis identified in this study strongly confirms phosphorous deficiency conditions in tropical soils.
Sala et al. (2000) already reported that plant K+ uptake is optimal on warm, moist soils which are well aerated with slightly acidic to neutral pH aided by the presence on mycorrhizal communities. Indigenous moringa (M. stenopetala) contrarily showed improved growth in vert+hist (<2 mm airspace). Gathumbi et al. (2002) also documented on improved plant performance at P and K abundance in experiments with legumes. From moringa plant performance analysis, it is clear that high K+ and P bioavailability in vert+hist soils influenced heights and basal stem values consistent with this literature. Additionally, K+ is associated with plant nutrient translocation and concentration of other macronutrients in soil solution (Yanai et al., 1996; Ashley et al., 2006; Vinichuk et al., 2010). Increased biomass records of moringa plants in the independent block registered improved nutrient uptake occurred at K+ addition in growth chamber, consistent with superior performance in rhizospheric vert+hist soils. Other factors such as pH differences could be of added value, since most minerals are made available at neutral pH where P availability is maximal at 5.5-6.5 pH (Sala et al., 2000) qualifying vert+hist (pH 5.7) as an ideal media for plant mineral availability to seedlings. However, mycorrhizal abundance could likewise be responsible for the pH balancing in this experiment at biotest, facilitating efficient uptake condition for moringa plants. Although, application of K+ salt in soils may increase biomass yield, this is only useful in lower quantities. More than 2 mM as KCl salt applied on M. oleifera was found to be excess (Chaves et al., 2005) leading to K+ accumulation on moringa plant biomass. However soil compaction common in clay/vert+hist soils is likewise associated with higher volumetric water content which tends to facilitate K+ transport to the root surface, a factor that may cause a reduction in the root length but not necessarily result in increased K+ accumulation (Seiffert et al., 1995; Ashley et al., 2006). Moringa samples growing in vert+hist soils with higher K+ collected from Lake Victoria field research had much fleshier (approximately >3 mm) roots functioning as fine roots.
Although the studies of Requena et al. (2001), Barea et al. (2005) and Meng et al. (2015) report on improved symbiosis, superior plant performance at co-inoculation (AMF+NFB) and the need to increase rhizobia in the hosts, K+ factor is vital in evaluating the strength of microbial interactions in the rhizosphere. Co-inoculation was more effective in increasing growth of M. stenopetala and M. oleifera in this experiment compared to single treatments. Our arguments support the fact that a major component of the N and P cycles which influence plant nutrition as well as a majority of soil fungal structure formation is determined more by soil factors than the surrounding plant communities. Moreover plant performance could be determined more by soil factor than the facilitated symbiotic input of the N and P guilds, which neglect K+ deficit in most symbiosis models in the environment. Evidence shows that soil aggregate size influences P sorption and bioavailability (Misra et al., 1988; Linquist et al., 1997). Moringa being non-obligate species requires time to be effective in symbiosis. Fine root aliquots analysed, showed only some signs of single vesicle occupation apart from few runner hyphae presence, save from vert+hist soils. In every situation out-cropping weedy competitors from the soils, formed co-operation with AMF and dark septate endophytes, a case that was not reported on moringa seedlings. Nevertheless, during younger stages in plant growth, mineral depletion effects in soils are mostly dependent on plant development rather than an uptake function (De Freitas et al., 1997; Mantelin and Touraine, 2004). Molecular science may offer deeper understanding on taxon-specific behavior related to associations at different stages in growth and levels of stress. Investigations on effective symbiosis could be elaborated with competition among fungi and other microbial communities steering evolution of offensive and defensive mechanisms, including the production of secondary metabolites (Rambold et al., 2013) that could be useful in assessing degree of colonization related to soil heterogeneity. Molecular approaches such as functional gene chips may also provide sufficient feedback on community processes important in gaining insights into colonization patterns ascribed to prevalent microbial guilds (gene to gene interactions) driven by target metabolomes.
ACKNOWLEDGMENT
The authors in this study would like to thank the Technische Universitaet Muenchen (TUM), Weihenstephan (TUM Graduate School) for field sampling financial support and the Soil Ecology department at Helmholtzzentrum, Muenchen.
REFERENCES
- Johnson, D., P.J. Vandenkoornhuyse, J.R. Leake, L. Gilbert and RE. Booth et al., 2004. Plant communities affect arbuscular mycorrhizal fungal diversity and community composition in grassland microcosms. New Phytol., 161: 503-515.
CrossRefDirect Link - Kengara, O.F., K. Schramm, U. Doerfler, J.C. Munch and B. Henkelmann et al., 2010. Degradation capacity of a 1,2,4-trichlorobenzene mineralizing microbial community for traces of organochlorine pesticides. Sci. Total Environ., 408: 3359-3366.
CrossRefDirect Link - Knopf, E., H. Blaschke and J.C. Munch, 2013. Improving moringa growth by using autochthonous and allochthonous arbuscular mycorrhizal fungi in lake Victoria basin. West Afr. J. Applied Ecol., 21: 47-57.
Direct Link - Knopf, E., H. Blaschke and J.C. Munch, 2014. Facilitating AMF activities in moringa species for reforestation purposes in lake Victoria basin, Kenya. Agric. Sci. Eng. Technol. Res., 2: 1-12.
Direct Link - Larson, W.E. and F.J. Pierce, 1991. Conservation and Enhancement of Soil Quality. In: Evaluation for Sustainable Land Management in the Developing World, Dumanski, J., E. Pushprajah, M. Latham and R. Myers (Eds.). Vol. 2, International Board for Soil Reseach and Management, Bankok, Thailand, pp: 175-203.
- Linquist, B.A., P.W. Singleton, R.S. Yost and K.G. Cassman, 1997. Aggregate size effects on the sorption and release of phosphorus in an Ultisol. Soil Sci. Soc. Am. J., 61: 160-166.
CrossRef - Mantelin, S. and B. Touraine, 2004. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake. J. Exp. Bot., 55: 27-34.
CrossRefDirect Link - Medina, A. and R. Azcon, 2010. Effectiveness of the application of arbuscular mycorrhiza fungi and organic amendments to improve soil quality and plant performance under stress conditions. J. Soil Sci. Plant Nutr., 10: 354-372.
CrossRefDirect Link - Meng, L., A. Zhang, F. Wang, X. Han, D. Wang and S. Li, 2015. Arbuscular mycorrhizal fungi and rhizobium facilitate nitrogen uptake and transfer in soybean/maize intercropping system. Front. Plant Sci., Vol. 6.
CrossRefDirect Link - Misra, R.K., A.M. Alston and A.R. Dexter, 1988. Root growth and phosphorus uptake in relation to the size and strength of soil aggregates. I. Experimental studies. Soil Tillage Res., 11: 103-116.
CrossRefDirect Link - Odada, E.O., W.O. Ochola and D.O. Olago, 2009. Drivers of ecosystem change and their impacts on human well-being in Lake Victoria basin. Afr. J. Ecol., 47: 46-54.
CrossRefDirect Link - Orlowska, E., W. Przybylowicz, D. Orlowski, K. Turnau and J. Mesjasz-Przybylowicz, 2011. The effect of mycorrhiza on the growth and elemental composition of Ni-hyperaccumulating plant Berkheya coddii roessler. Environ. Pollut., 159: 3730-3738.
CrossRefDirect Link - Ouahmane, L., J. Thioulouse, M. Hafidi, Y. Prin and M. Ducousso et al., 2007. Soil functional diversity and P solubilization from rock phosphate after inoculation with native or allochtonous arbuscular mycorrhizal fungi. For. Ecol. Manage., 241: 200-208.
CrossRefDirect Link - Pilon-Smits, E.A.H., C.F. Quinn, W. Tapken, M. Malagoli and M. Schiavon, 2009. Physiological functions of beneficial elements. Curr. Opin. Plant Biol., 12: 267-274.
CrossRefDirect Link - Poorter, H. and O. Nagel, 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant. Physiol., 27: 595-607.
Direct Link - Rambold, G., M. Stadler and D. Begerow, 2013. Mycology should be recognized as a field in biology at eye level with other major disciplines-a memorandum. Mycol. Progr., 12: 455-463.
CrossRefDirect Link - Requena, N., E. Perez-Solis, C. Azcon-Aguilar, P. Jeffries and J.M. Barea, 2001. Management of indigenous plant-microbe symbioses aids restoration of desertified ecosystems. Applied Environ. Microbiol., 67: 495-498.
CrossRefDirect Link - Romera, F.J., E. Alcantara and M.D. de la Guardia, 1992. Effects of bicarbonate, phosphate and high pH on the reducing capacity of Fe-deficient sunflower and cucumber plants. J. Plant Nutr., 15: 1519-1530.
CrossRefDirect Link - Schutz, K., E. Kandeler, P. Nagel, S. Scheu and L. Ruess, 2010. Functional microbial community response to nutrient pulses by artificial groundwater recharge practice in surface soils and subsoils. FEMS Microbiol. Ecol., 72: 445-455.
CrossRefDirect Link - Seiffert, S., J. Kaselowsky, A. Jungk and N. Claassen, 1995. Observed and calculated potassium uptake by maize as affected by soil water content and bulk density. Agron. J., 87: 1070-1077.
CrossRefDirect Link - Unterseher, M., M. Schnittler, C. Dormann and A. Sickert, 2008. Application of species richness estimators for the assessment of fungal diversity. FEMS Microbiol. Lett., 282: 205-213.
CrossRefDirect Link - Vierheilig, H., A.P. Coughlan, U. Wyss and Y. Piche, 1998. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Applied Environ. Microbiol., 64: 5004-5007.
Direct Link - Vinichuk, M., A.F.S. Taylor, K. Rosen and K.J. Johanson, 2010. Accumulation of potassium, rubidium and caesium (133Cs and 137Cs) in various fractions of soil and fungi in a Swedish forest. Sci. Total Environ., 408: 2543-2548.
CrossRefDirect Link - Wagg, C., J. Jansa, M. Stadler, B. Schmid and M.G. van der Heijden, 2011. Mycorrhizal fungal identity and diversity relaxes plant-plant competition. Ecology, 92: 1303-1313.
PubMedDirect Link - White, P.J. and P.H. Brown, 2010. Plant nutrition for sustainable development and global health. Ann. Bot., 105: 1073-1080.
CrossRefDirect Link - Wright, S.F., V.S. Green and M.A. Cavigelli, 2007. Glomalin in aggregate size classes from three different farming systems. Soil Tillage Res., 94: 546-549.
CrossRefDirect Link - Yanai, J., D.J. Linehan, D. Robinson, I.M. Young, C.A. Hackett, K. Kyuma and T. Kosaki, 1996. Effects of inorganic nitrogen application on the dynamics of the soil solution composition in the root zone of maize. Plant Soil J., 180: 1-9.
CrossRefDirect Link - Trouvelot, A., J.L. Kough and V. Gianinazzi-Pearson, 1986. Mesure du Taux de Mycorhization d'un Systeme Radiculaire. Recherche de Methodes Destimation Ayant une Signification Fonctionnelle. In: Physiological and Genetical Aspects of Mycorrhizae, Gianinazzi-Pearson, V. and S. Gianinazzi (Eds). INRA., Dijon, France, pp: 217-221.
- Alguacil, M.M., E. Lumini, A. Roldan, J.R. Salinas-Garcia, P. Bonfante and V. Bianciotto, 2008. The impact of tillage practices on arbuscular mycorrhizal fungal diversity in subtropical crops. Ecol. Applic., 18: 527-536.
CrossRefDirect Link - Ashley, M.K., M. Grant and A. Grabov, 2006. Plant responses to potassium deficiencies: A role for potassium transport proteins. J. Exp. Bot., 57: 425-436.
CrossRefDirect Link - Auge, R.M., A.J.W. Stodola, J.E. Tims and A.M. Saxton, 2001. Moisture retention properties of a mycorrhizal soil. Plant Soil, 230: 87-97.
CrossRefDirect Link - Ayssiwede, S.B., J.C. Zanmenou, Y. Issa, M.B. Hane and A. Dieng et al., 2011. Nutrient composition of some unconventional and local feed resources available in senegal and recoverable in indigenous chickens or animal feeding. Pak. J. Nutr., 10: 707-717.
CrossRefDirect Link - Barancikova, G., J. Halas, M. Guttekova, J. Makovnikova, M. Novakova, R. Skalsky and Z. Tarasovicova, 2010. Application of RothC model to predict soil organic carbon stock on agricultural soils of Slovakia. Soil Water Res., 5: 1-9.
Direct Link - Barea, J.M., M.J. Pozo, R. Azcon and C. Azcon-Aguilar, 2005. Microbial co-operation in the rhizosphere. J. Exp. Bot., 56: 1761-1778.
CrossRefDirect Link - Brundrett, M.C., 2009. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil, 320: 37-77.
CrossRefDirect Link - Chaves, L.H.G., R.A. Viegas, A.C.F. de Vasconcelos and H. Vieira, 2005. Effect of potassium on moringa plants growth in nutriente solution. Revista Biologia Ciencias Terra, Vol. 5.
Direct Link - De Freitas, J.R., M.R. Banerjee and J.J. Germida, 1997. Phosphate-solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica napus L.). Biol. Fertil. Soils, 24: 358-364.
CrossRefDirect Link - De Deyn, G.B., H. Quirk, S. Oakley, N. Ostle and R.D. Bardgett, 2011. Rapid transfer of photosynthetic carbon through the plant-soil system in differently managed species-rich grasslands. Biogeosciences, 8: 1131-1139.
CrossRefDirect Link - Figueiredo, M.V.B., C.R. Martinez, H.A. Burity and C.P. Chanway, 2008. Plant growth-promoting rhizobacteria for improving nodulation and nitrogen fixation in the common bean (Phaseolus vulgaris L.). World Microbiol. Biotechnol., 24: 1187-1193.
CrossRefDirect Link - Galloway, J.N., A.R. Townsend, J.W. Erisman, M. Bekunda and G. Cai et al., 2008. Transformation of the nitrogen cycle: Recent trends, questions and potential solutions. Science, 320: 889-892.
CrossRefDirect Link - Gathumbi, S.M., G. Cadisch and K.E. Giller, 2002. 15N natural abundance as a tool for assessing N2-fixation of herbaceous, shrub and tree legumes in improved fallows. Soil Biol. Biochem., 34: 1059-1071.
Direct Link - Giovannetti, M., 1985. Seasonal variations of vesicular-arbuscular mycorrhizas and endogonaceous spores in a maritime sand dune. Trans. Br. Mycol. Soc., 84: 679-684.
CrossRefDirect Link - Hildebrandt, U., M. Kaldorf and H. Bothe, 1999. The zinc violet and its colonization by arbuscular mycorrhizal fungi. J. Plant Physiol., 154: 709-717.
Direct Link - Jones, C., C. McConnell, K. Coleman, P. Cox, P. Falloon, D. Jenkinson and D. Powlson, 2005. Global climate change and soil carbon stocks; predictions from two contrasting models for the turnover of organic carbon in soil. Global Change Biol., 11: 154-166.
CrossRefDirect Link - Carter, M.R., 2002. Soil quality for sustainable land management: Organic matter and aggregation interactions that maintain soil functions. Agron. J., 94: 38-47.
Direct Link