
Raviteja Pagor
Faculty of Biotechnology, Jeju National University, Jeju, 690-756, Korea
Md. Shafiqul Islam Sikdar
Department of Agronomy, Hajee Mohammad Danesh Science and Technology University, Dinajpur, 5200, Bangladesh
Eun-Sung Moon
Faculty of Biotechnology, Jeju National University, Jeju, 690-756, Korea
Jung-Sup Kim
Faculty of Biotechnology, Jeju National University, Jeju, 690-756, Korea
Journal of Biological Sciences
Year: 2015 | Volume: 15 | Issue: 1 | Page No.: 33-39







ABSTRACT
3-deoxy-D-arabino-heptulosonate 7-phosphate synthase (DAHPS) is an entry enzyme of the shikimate pathway which is responsible for primary carbohydrate metabolism with the biosynthesis of most aromatic amino acids in microorganisms and plants. Enzymes of the pathway are drawing attention in the recent years because they are recognized as potential targets for design of antimicrobial drugs and herbicides. Analysis of sequences from rice revealed that the OsDAHPS showed a full-length open reading frame consisting of 537 amino acids, which encoded for a protein of approximately 59.0 kDa. The predicted amino acid sequence of OsDAHPS is highly homologous to those of DAHPS enzymes from many plants. The OsDAHPS expression in a aroH mutant of Escherichia coli showed that the gene was functionally capable of complementing the mutant. These results showed that the OsDAHPS encoded for a protein in 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase in rice.
PDF Abstract XML References Citation
Received: April 07, 2015;
Accepted: July 15, 2015;
Published: August 27, 2015
How to cite this article
Raviteja Pagor, Md. Shafiqul Islam Sikdar, Eun-Sung Moon and Jung-Sup Kim, 2015. Characterization of a Gene Encoding 3-Deoxy-D-Arabino-Heptulosonate 7-Phosphate Synthase from Rice. Journal of Biological Sciences, 15: 33-39.
DOI: 10.3923/jbs.2015.33.39
URL: https://scialert.net/abstract/?doi=jbs.2015.33.39
DOI: 10.3923/jbs.2015.33.39
URL: https://scialert.net/abstract/?doi=jbs.2015.33.39
INTRODUCTION
The shikimate pathway is consist of the several enzymes that catalyze the first seven reactions in the synthesis of a diverse set of important aromatic compounds in multiple organisms converting primary carbon metabolites into chorismate. Chorismate is a common precursor for the synthesis of a number of aromatic metabolites, vitamin K, folic acid, ubiquinone and other secondary metabolites including the three aromatic amino acids (AAA): phenylalanine (Phe), tyrosine (Tyr) and tryptophan (Trp) (Bentley and Haslam, 1990; Roberts et al., 1998; Ma et al., 2012; Light and Anderson, 2013). In plants, the shikimate and AAA biosynthetic pathways provide the substrates for numerous secondary metabolites, such as alkaloids, flavonoids, lignin, coumarins, indole derivatives, para-amino benzoic acid and other phenolic compounds (Gilchrist and Kosuge, 1980). The pathway is a bridge between primary and secondary metabolism. Therefore, it is being looked upon as an attractive target for the development of herbicides and antimicrobial agents against a plethora of diseases (Bentley and Haslam, 1990). The shikimate pathway is necessary for it’s an important target for the development of antibiotics and herbicides such as glyphosate in plants and microorganisms. Glyphosate is a commercially important herbicide that inhibit to 5-enolpyruvylshikimate-3-phosphate synthase, which catalyzes the sixth step in the pathway (Steinrucken and Amrhein, 1980).
The first reaction in the shikimate pathway that convert phosphoenolpyruvate (PEP) and erythrose-4-phosphate (E4P) to chorismate links between the primary and specialized metabolism leading to aromatic amino acids (Fig. 1), provides an unusually fertile ground to examine the role of quaternary structure in enzyme function and regulation.
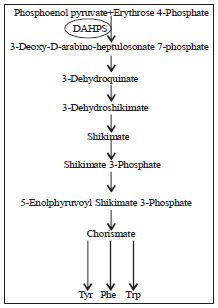 | |
| Fig. 1: | Schematic diagram of the shikimate pathway in plants |
The first committed enzyme of the shikimate pathway, the aroH gene encoding enzyme is 3-deoxy-D-arabino-heptulosonate-7- phosphate synthase (DAHPS; EC 4.1.2.15), which catalyses the first committed step in the biosynthesis of aromatic compounds (Fig. 1) in bacteria, fungi, plants and some lower eukaryotes the conversion of PEP and E4P to 3-deoxy-D-arabino-heptulosonate 7-phosphate and the phosphate group of PEP, the first reaction in the process (Schoner and Herrmann, 1976; Herrmann, 1995; Nazmi et al., 2014). As the first enzyme in the pathway DAHPS is located at a central gateway, where pathway input can be competently controlled in response to modify in the cellular concentration of pathway outputs. Evolutionary processes have led to at least four different mechanisms of conferring DAHPSs allosterically responsive to downstream reaction products (Light et al., 2012). The activities of DAHPS enzyme is generally regulated by inhibition with end-product amino acids Tyr, Phe, Trp, or other intermediates of the pathway (Cross et al., 2013).
Although, the regulation of the synthesis of AAAs from chorismate has been studied extensively in plants (Yamada et al., 2008; Tzin et al., 2009; Maeda et al., 2011). The most studied enzyme of the shikimate pathway is enolpyruvylshikimate 3-phosphate synthase (EPSPS), catalyzing the formation of enolpyruvylshikimate 3-phosphate, because of its association with resistance to the herbicide glyphosate (Singer and McDaniel, 1985; Smart et al., 1985; Klee et al., 1987; Duke and Powles, 2008; Vivancos et al., 2011).
The cDNA encoding DAHPS has been identified from some micro-organism and plants including tobacco (Wang et al., 1991), Arabidopsis thaliana (Keith et al., 1991), potato (Dyer et al., 1990; Zhao and Herrmann, 1992), tomato (Gorlach et al., 1993; Tzin et al., 2012) and Populus trichocarpa (Tuskan et al., 2006), grapes (Zhang et al., 2011), Thermotoga maritima (Cross and Parker, 2013), Corynebacterium glutamicum (Li et al., 2013); Neisseria meningitidis (Cross et al., 2013) and Hyperthermophile pyrococcus (Nazmi et al., 2014). To date, there is no studies elucidation and manipulation of metabolic regulatory in the conversion of primary metabolism into aromatic metabolites of rice genes of shikimate pathway. Hence, it is necessary to study the shikimate pathway in rice so, as to better uncover the formation and regulation mechanism of diverse flavonoids in rice crop. To elucidate the impact of the shikimate pathway on the production of specialized metabolites in rice, this work expressed a bacterial Phe-feedback-insensitive aroH under a fruit-specific promoter in transgenic plants. The manipulation of the rice gene that encoding DAHPS enzyme controlling the conversion of primary to specialized metabolism could be an attractive tool for improving rice aroma and flavor qualities. Here, we report the analysis and characterization of a gene for the DAHPS enzyme from rice (Oryza sativa), an important crop plant.
MATERIALS AND METHODS
Mutant strains and plasmids: The mutant strains which were used in this study are ME8637 and JW0001. The genotypes of the mutant strains are ME8637 (aroH367, thi-1, argE3, proA2, galK2, lacY1, tsx-29) and JW0001 (thrA). Both mutant strains are obtained from National Bio-Resource Project (NBRP) Japan. The EST clone carrying the expected gene encoding for DAHP synthase, having the GenBank accession number AK069968 and clone number is J023038D13 that was collected from Rice Genome Resource Center (RGRC) Japan.
DNA sequence analysis: The sequence analysis of rice EST clone (J023038D13), which was derived from rice cDNA library (Osato et al., 2002) from developing seeds prepared in pBluescript that was done by using various bioinformatics tools. The DNA sequencing and sequence analysis were described previously (Sikdar and Kim, 2010). The nucleotide and amino acid sequences were compared with various sequences present in GenBank and were analyzed by using Nucleotide-BLAST (Wheeler et al., 2003) and CLUSTAL X a multiple sequence alignment program (Thompson et al., 1994). Motifs regions were searched by the Genome Net computation Service at Kyoto University (http://www.genome. ad.jp) and the phylogenic tree analysis was performed by MEGA 4.1 neighbor_joining program (Kumar et al., 2008).
Polymerase chain reaction and recombinant construct: Particular primers were designed from the sequence of OsDAHPS information surrounding the translational start and stop codons of OsDAHPS to amplify the full-length Open Reading Frame (ORF) and to construct a recombinant DNA to express the gene product in E. coli. Polymerase Chain Reaction (PCR) was conducted in accordance with the method described by Sambrook and Russell (2001). The ORF region of OsDAHPS was amplified by own designed forward and reverse primers. The cDNA of OsDAHPS was amplified by using the following designed forward and reverse primers from the OsDAHPS sequence; OsDAHPS-F (5’-GCATGAGGATCCAATGGCGCTCGCCACCAA-3’) OsDAHPS-R (5’-GCATGAAAGCTTCTTCCCCCGTTTCA GCAGTTTA-3’) both the primers contain BamHI and HindIII restriction sites at 5’ end. The PCR reaction was conducted using a MY Cyler TM PCR system (BioRad, U.S.A) for 35 cycles with denaturation at 95°C for 10 min, annealing at 58°C for 1 min and extension 72°C for 1 min, with 10 μM primers. The PCR products were analyzed on 1% (w/v) agarose gel (data not shown) and then subcloned into the BamHI and HindIII site of pBluescript II plasmid to construct OsDAHPS. Restriction analysis was conducted to confirm the orientation of the contract.
Growth determination in E. coli mutant strains: The transformed E. coli mutant strains ME8637 harbouring pB:OsDAHPS, control plasmid and wild type (JW0001) harboring control plasmid. Single colony was inoculated in 1 mL of LB broth with Amp (50 μg mL–1) and incubated at 37°C for overnight with shaking. The 1 mL overnight cultured medium were grown in 100 mL of M9 minimal media containing 1 mM IPTG, 20% glucose, amp (50 μg mL–1) and 19 amino acids excluding Trp and the same medium was used with all the reagents kept constant. The bacterial growth of E. coli was monitored for every hour by measuring the optical density measurement with a UV-spectrophotometer (UV1101, Biochrom, England) at 595 nm (OD595).
RESULTS
Sequence analysis of OsDAHPS: The EST clone (clone ID: J023038D13) obtained from the RGRC was analyzed to determine the nucleotide sequence using the designed primers and bioinformatics tools. The results of data analysis with the cDNA sequence and complete rice genome indicated that an ORF was identical to that of the rice genomic region located on chromosome VII (Os07g42960) in rice sequence. The cDNA sequence (OsDAHPS) contained a full-length open reading frame consisting of 1614 bp and encoded for a protein with 537 amino acids and approximate molecular weight of 59.0 kDa. The expected isoelectric point of the protein was 8.266. The sequences of different species and organisms such as rice, Arabidopsis, potato, bacteria and fungi were analyzed. The most of the regions are highly conserved in plant species showing that they were sharing similar domains but in the case of bacteria and fungi they share very few conserved regions with plant species. A sequence comparison of the predicted amino acids for the OsDAHPS with the deduced sequences from maize (Zea mays), Arabidopsis (A. thaliana), Ricimus communi, Saccharomyces cerevisiae and E. coli evidenced a high degree of homology with identity values of 95, 97, 95, 19 and 17%, respectively (Fig. 2). Motif analysis of the amino acid sequences of DAHPS revealed that there is one motif found from 77-514 in the sequence OsDAHPS (Fig. 2). The OsDAHPS is highly conserved with other plant DAHPS and shows very less similarity with bacteria and fungi DAHPS. This evident shows that plant has two types DAHPS such as type II and type I. Type I DAHP synthases are commonly found in microorganisms. The phylogenetic tree derived from the related sequences showed that OsDAHPS is divergent and evolved from ancestor bacterial DAHPS using the MEGA 4.1 program (Kumar et al., 2008). The branching pattern and Numbers at the nodes indicate levels of bootstrap support based on neighbor_joining analyses of 1000 resampled data sets using the same program (Fig. 3).
Expression in E. coli and in vivo activity: The recombinant DNA, pB::OsDAHPS was constructed using the ORF of a PCR-amplified OsDAHPS fragment. After the transformation of E. coli with the recombinant DNA, OsDAHPS activity was observed in vivo in a medium containing IPTG and 19 amino acids, excluding Trp. Functional complementation was carried out using the DAHPS mutant of E. coli to prove the enzyme activity of the gene product of OsDAHPS. To evaluate the viability of E. coli cells by OsDAHPS activity, the OsDAHPS -expressing cells were cultured for overnight at 37°C with shaking and the diluted portion of culture was plated on agar medium containing the 19 amino acids except Trp with Amp (50 mg mL–1). The viable colonies greatly differed on the agar medium among the plasmids. The aroH mutant of E. coli with OsDAHPS could grow under conditions in which the mutant without OsDAHPS could not. This showed that the OsDAHPS was capable of functioning as a complement and evidenced functional DAHPS activity.
Expression of OsDAHPS can complement the aroH mutant of E. coli: A growth study of the pB::OsDAHPS activity was monitored to determine by functional complementing the aroH mutant E. coli strain. The pB::OsDAHPS construct was transformed into E. coli mutant aroH strain ME8637. The was used for the transformation in this experiment, the pB:OsDAHPS construct was transformed into ME8637 E. coli mutant strain. A control plasmid was also transformed into wild type (JW0001) and the aroH strain ME8637 to differentiate the activity of gene in minimal media without Trp. All of these transformed mutant strains were monitored via a growth assay for functional complementation in MM media with 1mM IPTG, AMP (50 μg mL–1) containing 19 amino acids excluding Trp. The E. coli strain harboring pB::OsDAHPS which was encoded by aroH showed they grew normally under tryptophan deficit condition in the MM media, this result indicated that the aroH strain was able to synthesize tryptophan, which grew normally in the medium and thus produced S-shaped growth curve.
 | |
| Fig. 2: | Multiple sequence alignment of DAHPS was analyzed by using Boxshade program. The amino acid sequences of DAHPS from different organisms that were obtained from NCBI database such as Oryza sativa (OsDAHPS; accession number: AK069968), Arabidopsis thaliana (AtDAHPS; accession number: NP_195077), Saccharomyces cerevisiae (ScDAHPS; accession number: NP_010320) and Escherichia coli (EcDAHPS; accession number:NP_415275), (In figure, Characters shows different color like: Completely Conserved Residues, Identical Residues, Similar Residues and Different Residues are green, cyan, magenta and white color, respectively) |
The wild type E. coli strain JW0001 harboring the control plasmid also grew normally and evidenced an S-shaped classical growth curve almost similar the pB::OsDAHPS which was encoded by aroH. The wild type train contain aroH as it mutant of thrB, so it was able to encode for DAHPS, therefore showed the activity in the medium by synthesizing the trp, as evident of S-shaped growth curve. The E. coli strain which was encoded by aroH harboring control plasmid was incubated in the same MM medium without Trp that evidenced dramatically retarded growth. In this case, the aroH mutant E. coli strains ME8637 could not synthesize Trp itself, that’s why was not able to grow normally. However, the same E. coli strains ME8637 containing pB::OsDAHPS grew well because the aroH mutants E. coli was able to synthesize Trp using DAHPS expressed by the pB::OsDAHPS plasmid (Fig. 4). This is a consequence of pB::OsDAHPS activity. From the above finding, it was concluded that OsDAHPS expression could be functionally complement the aroH mutants E. coli.
DISCUSSION
It was reported that there is the transit signal for chloroplasts in some plants including tobacco (Wang et al., 1991). The classical transit peptide characteristic with many hydroxyl amino acids such as Ser and Thr is also found in the N-terminal of OsDAHPS which might be active for the biosynthesis of aromatic amino acids and other secondary metabolites in chloroplasts.
There are three DAHPS isoenzymes which are regulated by Tyr, Phe, Trp, respectively, in E. coli (Kikuchi et al., 1997).
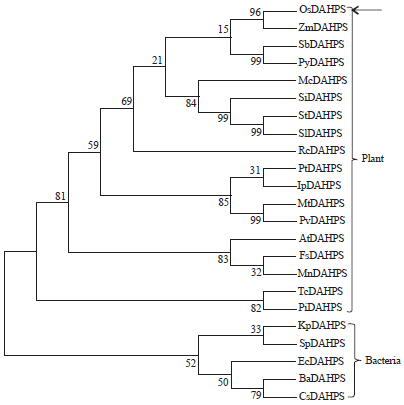 | |
| Fig. 3: | Phylogenetic tree: Phylogenetic analysis of OsTS related proteins using Clustal W and Mega 4.1 program. GenBank accession numbers of above protein sequences are as follows: AK069968 (OsDAHPS from Oryza Sativa), NM 001157394 (ZmDAHPS; Zea mays), XP_002463199 (SbDAHPS; Sorghum bicolor), AFL02467 (PyDAHPS; Petunia hybrid), CAA7509223 (McDAHPS; Morinda citrifolia), XP_004984156 (SiDAHPS; Setaria italic), NP_001275361 (StDAHPS; Solanum tuberosum), NP_001234418 (SlDAHPS; Solanum lycopersicum), XM_002513905 (RcDAHPS; Ricinus communis), XP_002307364 (PtDAHPS; Populus trichocarpa), AFD62807 (IpDAHPS; Ipomoea purpurea), XP_003615152 (MtDAHPS; Medicago truncatula), AGV54666 (PvDAHPS; Phaseolus vulgaris), NP_195077 (AtDAHPS; Arabidopsis thaliana), ABA54865 (FsDAHPS; Fagus sylvatica), EXB75364 (MnDAHPS; Morus notabilis), XP_007042911 (TcDAHPS; Theobroma cacao), XP_002908379 (PiDAHPS; Phytophthora infestans), YP_005955985 (KpDAHPS; Klebsiella pneumonia, YP_003877294 (SpDAHPS; Streptococcus pneumonia), NP_415275 (EcDAHPS; Escherichia coli), YP_005580695 (BaDAHPS; Bifidobacterium animalis) and YP_003543340 (CsDAHPS; Candidatus sulcia) |
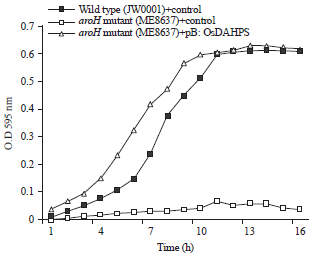 | |
| Fig. 4: | Growth curves of the aroH mutant strain ME8637 harboring pB::OsDAHPS. Bacterial cells were grown at 37°C in MM containing 19 amino acids excluding Trp. Growth was monitored via optical density measurements at 595 nm (OD595) |
It seems that there are more than four genes (Os07g0622200, Os03g0389700, Os08g0484500, Os10g0564400) in rice chromosome III, VII, VIII and X by searching in rice genome with the OsDAHPS sequence, even two homologous genes were reported in Arabidopsis, potato, tomato and grapes (Keith et al., 1991; Zhao and Herrmann, 1992; Gorlach et al., 1993; Zhang et al., 2011). These isoenzymes might be different in the sensitivity of feedback regulations by Tyr or other final products. Further research about these isoenzymes would be elucidated the overall regulation of shikimate pathway including the biosynthesis of aromatic amino acids and other secondary metabolites.
Current research elucidated that DAHPS is inhibited by Phe and would use a target for therapeutic agent against Neisseria meningitides (Cross et al., 2013).
Next step would be find out some important clues about the substrate specificity of the enzyme by purifying recombinant pB::OsDAHPS S in E. coli and physiological functions of this novel enzyme by screening T-DNA insertion mutants in which the OsDAHPS gene is knocked out in rice. Our reports about cloning and characterization of the cDNA encoding DAHPS from rice confirm the predictions and extend to motifs and expression in E. coli. The DAHPS gene could be used as a powerful tool for future application to improve the aromatic amino acids as well as others secondary metabolic compounds in rice plants.
ACKNOWLEDGMENTS
We wish to thank Rice Genome Resource Center (RGRC), National Institute of Agro biological Science (NIAS), Japan and National Bio-Resource Project (NBRP) Japan for providing an EST clone AK120254 and ME8637 mutant and wild type JW0001 of E. coli, respectively. This work was supported by the research grant of Jeju National University in 2012.
REFERENCES
- Bentley, R. and E. Haslam, 1990. The shikimate pathway-a metabolic tree with many branche. Crit. Rev. Biochem. Mol. Biol., 25: 307-384.
CrossRefDirect Link - Cross, P.J. and E.J. Parker, 2013. Allosteric inhibitor specificity of Thermotoga maritima 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase. FEBS Lett., 587: 3063-3068.
CrossRefDirect Link - Cross, P.J., A.L. Pietersma, T.M. Allison, S.M. Wilson‐Coutts, F.C. Cochrane and E.J. Parker, 2013. Neisseria meningitidis expresses a single 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase that is inhibited primarily by phenylalanine. Prot. Sci., 22: 1087-1099.
CrossRefDirect Link - Dyer, W.E., L.M. Weaver, J.M. Zhao, D.N. Kuhn, S.C. Weller and K.M. Herrmann, 1990. A cDNA encoding 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase from Solanum tuberosum L. J. Biol. Chem., 265: 1608-1614.
Direct Link - Duke, S.O. and S.B. Powles, 2008. Glyphosate: A once-in-a-century herbicide. Pest Manage. Sci., 64: 319-325.
CrossRefDirect Link - Gorlach, J., A. Beck, J.M. Henstrand, A.K. Handa, K.M. Herrmann, J. Schmid and N. Amrhein, 1993. Differential expression of tomato (Lycopersicon esculentum L.) genes encoding shikimate pathway isoenzymes. I. 3-Deoxy-D-arabino-heptulosonate 7-phosphate synthase. Plant Mol. Biol., 23: 697-706.
CrossRefDirect Link - Herrmann, K.M., 1995. The shikimate pathway: Early steps in the biosynthesis of aromatic compounds. Plant Cell, 7: 907-919.
CrossRefDirect Link - Keith, B., X.N. Dong, F.M. Ausubel and G.R. Fink, 1991. Differential induction of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase genes in Arabidopsis thaliana by wounding and pathogenic attack. Proc. Natl. Acad. Sci. USA., 88: 8821-8825.
Direct Link - Kikuchi, Y., K. Tsujimoto and O. Kurahashi, 1997. Mutational analysis of the feedback sites of phenylalanine-sensitive 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase of Escherichia coli. Applied Environ. Microbiol., 63: 761-762.
PubMedDirect Link - Klee, H.J., Y.M. Muskopf and C.S. Gasser, 1987. Cloning of an Arabidopsis thaliana gene encoding 5-enolpyruvylshikimate-3-phosphate synthase: Sequence analysis and manipulation to obtain glyphosate-tolerant plants. Mol. Gen. Genet., 210: 437-442.
CrossRefDirect Link - Kumar, S., M. Nei, J. Dudley and K. Tamura, 2008. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Briefings Bioinf., 9: 299-306.
CrossRefPubMedDirect Link - Li, P.P., D.F. Li, D. Liu, Y.M. Liu, C. Liu and S.J. Liu, 2013. Interaction between DAHP synthase and chorismate mutase endows new regulation on DAHP synthase activity in Corynebacterium glutamicum. Applied Microbiol. Biotechnol., 97: 10373-10380.
CrossRefDirect Link - Light, S.H. and W.F. Anderson, 2013. The diversity of allosteric controls at the gateway to aromatic amino acid biosynthesis. Protein Sci., 22: 395-404.
CrossRefDirect Link - Light, S.H., A.S. Halavaty, G. Minasov, L. Shuvalova and W.F. Anderson, 2012. Structural analysis of a 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase with an N-terminal chorismate mutase-like regulatory domain. Protein Sci., 21: 887-895.
CrossRefDirect Link - Ma, N., L. Wei, Y. Fan and Q. Hua, 2012. Heterologous expression and characterization of soluble recombinant 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase from Actinosynnema pretiosum ssp. auranticum ATCC31565 through co-expression with Chaperones in Escherichia coli. Protein Exp. Purification, 82: 263-269.
CrossRefDirect Link - Maeda, H., H. Yoo and N. Dudareva, 2011. Prephenate aminotransferase directs plant phenylalanine biosynthesis via arogenate. Nat. Chem. Biol., 7: 19-21.
CrossRefDirect Link - Nazmi, A.R., L.R. Schofield, R.C. Dobson, G.B. Jameson and E.J. Parker, 2014. Destabilization of the homotetrameric assembly of 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase from the hyperthermophile Pyrococcus furiosus enhances enzymatic activity. J. Mol. Biol., 426: 656-673.
CrossRefDirect Link - Osato, N., M. Itoh, H. Konno, S. Kondo and K. Shibata et al., 2002. A computer-based method of selecting clones for a full-length cDNA project: Simultaneous collection of negligibly redundant and variant cDNAs. Genome Res., 12: 1127-1134.
CrossRefDirect Link - Roberts, F., C.W. Roberts, J.J. Johnson, D.E. Kyle and T. Krell et al., 1998. Evidence for the shikimate pathway in apicomplexan parasites. Nature, 393: 801-805.
CrossRefDirect Link - Schoner, R. and K.M. Herrmann, 1976. 3-Deoxy-D-arabino-heptulosonate 7-phosphate synthase. Purification, properties and kinetics of the tyrosine-sensitive isoenzyme from Escherichia coli. J. Biol. Chem., 251: 5440-5447.
Direct Link - Sikdar, S.I. and J. Kim, 2010. Functional analysis of a gene encoding threonine synthase from rice. Afr. J. Biotechnol., 9: 1122-1129.
Direct Link - Singer, S.R. and C.N. McDaniel, 1985. Selection of glyphosate-tolerant tobacco calli and the expression of this tolerance in regenerated plants. Plant Physiol., 78: 411-416.
Direct Link - Smart, C.C., D. Johanning, G. Muller and N. Amrhein, 1985. Selective overproduction of 5-enol-pyruvylshikimic acid 3-phosphate synthase in a plant cell culture which tolerates high doses of the herbicide glyphosate. J. Biol. Chem., 260: 16338-16346.
Direct Link - Steinrucken, H.C. and N. Amrhein, 1980. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvylshikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun., 94: 1207-1212.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Tuskan, G.A., S. DiFazio, S. Jansson, J. Bohlmann, I. Grigoriev and U. Hellsten et al., 2006. The genome of black cottonwood, Populus trichocarpa (Torr. and Gray). Science, 313: 1596-1604.
CrossRefDirect Link - Tzin, V., S. Malitsky, M.M.B. Zvi, M. Bedair, L. Sumner, A. Aharoni and G. Galili, 2012. Expression of a bacterial feedback-insensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase of the shikimate pathway in Arabidopsis elucidates potential metabolic bottlenecks between primary and secondary metabolism. New Phytol., 194: 430-439.
CrossRefDirect Link - Tzin, V., S. Malitsky, A. Aharoni and G. Galili, 2009. Expression of a bacterial bi-functional chorismate mutase/prephenate dehydratase modulates primary and secondary metabolism associated with aromatic amino acids in Arabidopsis. Plant J., 60: 156-167.
CrossRefDirect Link - Vivancos, P.D., S.P. Driscoll, C.A. Bulman, L. Ying and K. Emami et al., 2011. Perturbations of amino acid metabolism associated with glyphosate-dependent inhibition of shikimic acid metabolism affect cellular redox homeostasis and alter the abundance of proteins involved in photosynthesis and photorespiration. Plant Physiol., 157: 256-268.
Direct Link - Wang, Y., K.M. Herrmann, S.C. Weller and P.B. Goldsbrough, 1991. Cloning and nucleotide sequence of a complementary DNA encoding 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase from tobacco. Plant Physiol., 97: 847-848.
PubMedDirect Link - Wheeler, D.L., D.M. Church, S. Federhen, A.E. Lash and T.L. Madden et al., 2003. Database resources of the national center for biotechnology. Nucleic Acids Res., 31: 28-33.
CrossRefDirect Link - Yamada, T., F. Matsuda, K. Kasai, S. Fukuoka and K. Kitamura et al., 2008. Mutation of a rice gene encoding a phenylalanine biosynthetic enzyme results in accumulation of phenylalanine and tryptophan. Plant Cell, 20: 1316-1329.
Direct Link - Zhang, Z.Z., X.X. Li, B.Q. Zhu, Y.Q. Wen, C.Q. Duan and Q.H. Pan, 2011. Molecular characterization and expression analysis on two isogenes encoding 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase in grapes. Mol. Biol. Rep., 38: 4739-4747.
CrossRefDirect Link - Zhao, J. and K.M. Herrmann, 1992. Cloning and sequencing of a second cDNA encoding 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase from Solanum tuberosum L. Plant Physiol., 100: 1075-1076.
PubMedDirect Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link