
M. E. Abu-Ziada
Department of Botany, Faculty of Science, Mansoura University, Egypt
G.A. El Sherbeny
Department of Botany, Faculty of Science, Mansoura University, Egypt
H. M. Al-Jubawy
College of Education, Thi Qar University, Iraq
Journal of Biological Sciences
Year: 2015 | Volume: 15 | Issue: 1 | Page No.: 25-32







ABSTRACT
This study focused on ecological characteristics and bioactive metabolic products of Stellaria pallida (Dumort.) Piré. Twenty eight vascular plants related to 16 families were recorded in S. pallida community type. Poaceae, Barassicaceae and Asteraceae were the most important families. Chenopodium murale, Rumex dentatus and Euphorbia helioscopia were the common associates. Therophytes were the most frequent life-form (85.7%). The floristic composition of this community was strongly influenced by the Mediterranean elements (15 species). S. pallida preferred wet, loamy sand textured, non-saline, fertile and neutral-slightly alkaline soils. The aerial fraction of this Caryophyllaceous weed showed high nutritive value of 276.17 cal. /100 g. The polyphenols and flavonoids contents have been detected with mean values of 1.95 and 5.85%, respectively. The yield of volatile oils of the flowers was 143 mg/100 g. Twenty compounds were identified in the volatile oils of Stellaria pallida flowers.
PDF Abstract XML References Citation
Received: February 09, 2015;
Accepted: April 24, 2015;
Published: August 27, 2015
How to cite this article
M. E. Abu-Ziada, G.A. El Sherbeny and H. M. Al-Jubawy, 2015. Autecology and Bioactive Metabolites of Stellaria pallida Growing in North East Nile Delta, Egypt. Journal of Biological Sciences, 15: 25-32.
DOI: 10.3923/jbs.2015.25.32
URL: https://scialert.net/abstract/?doi=jbs.2015.25.32
DOI: 10.3923/jbs.2015.25.32
URL: https://scialert.net/abstract/?doi=jbs.2015.25.32
INTRODUCTION
Stellaria pallida is a delicate herbaceous winter-spring active weed. It is native to Europe with Mediterranean-continental type of geographic range (Wittig, 2004). The genus Stellaria is common in the Northern hemisphere (Chinnappa and Morton, 1984). Taxonomically, Stellaria belongs to family Caryophyllaceae, subfamily Alsinoideae and tribe Alsineae (Bittrich, 1993). The members of this subfamily are distinguished by their free sepals and exstipulate leaves (Charter and Heywood, 1993). Members of the family Caryophyllaceae are best known to contain many bioactive metabolic constituents (Arora and Sharma, 2012; Sindhu and Manorama, 2012; Mamadalieva et al., 2010).
In Egypt, the genus Stellaria is represented by two species namely; S. media (L.) Vill. and S. pallida (Dumort.) piré. The comparative diagnostic features of the two species are: Flowers are smaller and have no petals in S. pallida. Seeds are smaller, thinner and paler in colour in S. pallida as compared with those of S. media. Stellaria media has been used as traditional Chinese medicine for over 200 years; it has been used as an anti-viral agent (Ma et al., 2012; Kokate, 2003). The second species, S. pallida is well known as an invasive weed in cultivated fields (Shan et al., 2010). This plant is reported to be useful in inflammation of the digestive, renal, respiratory and reproductive tracts. It is also possesses diuretic, expectorant and anti-asthmatic properties (Kirtikar and Basu, 2006; Kitanov, 1992). Due to the high nutritional value, it is used as a leaf vegetable, often raw in salads (Kirtikar and Basu, 2006).
The present study is an attempt to throw further light on morphology, anatomy and autecology of S. pallida in addition to evaluation of the primary metabolites, polyphenols, flavonoids and volatile oils of this plant.
MATERIALS AND METHODS
Morphological description: Fresh specimens were described according to Foster and Gifford (1974) and LAWG (1999).
Anatomical investigation: Plant materials were collected and immediately immersed in fixative solution of formalin-acetic acid-alcohol (10:5:85 v/v). Thin cross-sections of stem, leaf and root were made as described by Jensen (1962) and Peacock and Bradbury (1973). The sections were examined using light microscope and were photographed.
Ecological characteristics
Vegetation analysis: Twenty stands dominated by S. pallida were studied in detail including: Total coverage, a list of species, phenological aspect of growth and cover-abundance estimate of each species. Identification and nomenclature of the species were followed Tackholm (1974) and Boulos (1995). Techniques of floristic analysis based on the methods given in Kent and Coker (1992), Muller-Dombois and Ellenberg (1974) and Poole (1974).
Habitat conditions: One soil sample was collected from each stand (depth 0-20 cm), air dried and sieved through a 2 mm sieve to remove gravel and debris. The procedures followed in estimating their variables were according to the methods recommended by Carter and Gregorich (2008), Pansu and Gautheyrou (2006), Margesin and Schinner (2005), Baruah and Barthakur (1997) and Klute (1986).
Phytochemical analysis
Quantitative estimation of the primary metabolites: Fine powdered aerial parts of S. pallida were used to determine the mean values of moisture content, total ash, acid-insoluble ash, crude fibers, total lipids, protein, total carbohydrates and soluble sugars were determined as described by Van Handel (1968), Sadasivam and Manickam (2008) and AOAC (2000). The nutrient value was calculated as cited from Indrayan et al. (2005).
Qualitative estimation of the secondary metabolites: The phytochemical screening of sterols, terpenes, flavonoids, phenols, tannins, saponins and alkaloids was carried out using the procedures described by Harborne (1973), Sofowara (1993) and Trease and Evans (1989).
Quantitative estimation of polyphenols and flavonoids: The polyphenols content was determined using the method described by Sadasivam and Manickam (2008). The flavonoids were extracted and subjected to column chromatograph according to Boham and Kocipai-Abyazan (1994).
Volatile oils investigation: Qualitative and quantitative analysis of the volatile oils of S. pallida flowers was performed using Gas Chromatography (GC) and Gas Chromatography Coupled with Mass Spectrometry (GC-MS) (Adams, 2007).
RESULTS AND DISCUSSION
Morphology: Stellaria pallida (Fig. 1) is an annual herbaceous weed, decumbent or ascending, 10-20 cm to a maximum 40 cm long, much-branched and yellowish-green in colour. Leaves are cauline, pale-green, exstipulate, margin entire and apex acuminate. Leaves are pinnately veined with actinodromous primary vein category and reticulodromous secondaries. Veins spacing smoothly increasing towards the base. Flowers in late winter and self-pollinated because the flowers never open widely (cleistogamous).
 | |
| Fig. 1: | Close up view of S. pallida plant showing flowers in terminal cymes |
Flowers are borne in terminal 3-6 flowered cymes. Sepals are 5 free, persistent, hairy and lanceolate. Petals are absent. Stamens are 3 with grey anthers, longitudinally dehiscent and inserted on annular disc. Ovary is superior with many ovules. The fruit is simple dry dehiscent capsule, dehiscing by valves, straw coloured and ovoid (2-3 mm). The seeds are pale brown, reniform, 7-11 seeds/fruit and possess phanerocotylar germination. Stellaria pallada possesses survival strategies through seasonality and seeds dimorphism. The findings of the present morphological study are relatively correlated to the earlier reports (Chinnappa and Morton, 1984; Harbaugh et al., 2010; Miller and West, 2012).
Anatomy: The stem anatomy of S. pallida is presented in (Fig. 2a). It is a typical herbaceous dicot stem with normal secondary thickening and produced a small amount of secondary tissues. The epidermis is a single layer of quadrangular cells. Adjacent to the epidermis, the cortex starts with two layers of collenchymas then a wide zone composed of 4-9 layers of parenchymatous cells. The outer few layers of the cortical cells are assimilatory in nature due to the presence of chloroplasts. The vascular bundles were eleven and situated in a ring. The pith in the central part and comprised a hollow space.
The leaf cross-section (Fig. 2b) showed that, it is of the bifacial type and some spongy cells possess druses crystals of Ca-oxalate. The lower epidermal cells were larger than the upper epidermal cells. The midrib showed arc shaped vascular bundle.
The root cross-section (Fig. 2c) is circular. Beneath the epidermis there is 3-5 layered cortex of parenchyma cells, followed by multilayered periderm. The epidermal and outer cortical cells become fractured and disintegrated due to increasing pressure from the developing secondary tissues.
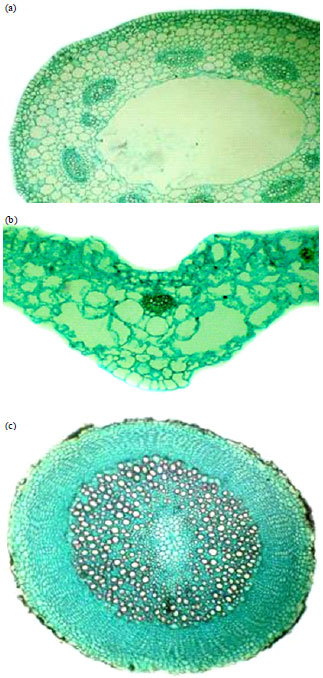 | |
| Fig. 2(a-c): | Cross section in (a) Stem of S. pallida, (b) Leaf of S. pallida and (c) Root of S. pallida |
A complete cylinder of secondary xylem is formed under the cambium ring with xylem vessels are wide arranged irregularly. The primary phloem strands are forced outward and digested. The pith is narrow and formed of parenchymatous ovoid cells. The obtained results are in accordance with those of Fahn (1982), Arora and Sharma (2012) and Abu Ziada et al. (2013).
The chromosomes count revealed that, the chromosome number of S. pallida was found to be 2n = 22. This agrees with the results reported by Ghaffari and Kelich (2006).
Autecology
Vegetation: Table 1 gives the floristic composition of twenty stands representing Stellaria pallida community type. It is obvious that S. pallida consistently the dominant, present in all the stands and has the highest cover-abundance estimate. This community has only (22 dicots and 5 monocots) belong to 16 families. Poaceae, Brassicaceae and Asteraceae are the most important. Chenopodium murale is the most abundant associate species (p = 100%). Rumex dentatus, Euphorbia helioscopia and Capsella bursa-pastorisare common, have presence estimates of 95% each. Beside the above mentioned associates, nine species have presence estimates of 30-80% in addition to other 13 with presence estimates 5-25%.
Regarding the life-span, the recorded species are classified into 23 annuals and 5 perennials. The dominance of the annuals could be attributed to the fact that, annuals have high reproductive capacity; ecological, morphological and genetic plasticity under high levels of environmental disturbance (Sadek, 2008).
According to the life-forms, these species are grouped under three types: therophytes (24 species = 85.7%). Geophytes (2 species = 7.1%) and hemicryptophytes.
The life-forms provide information which may help in assessing the response of vegetation to variations in certain environmental factors. The life-form spectra are important physiognomic attributes which have been widely used by ecologists in vegetation studies. The present study showed that therophytes are the most frequent type, followed by geophytes and hemicryptophytes. This trend is similar to spectra reported in the Egyptian weed flora (Mussa, 2002; Al-Sodany, 1992; El-Fahar, 1989).
Dealing with chorotypes (Table 2), the floristic components of this community belong to 7 categories: Mediterranean taxa (15 species), Irano-Turanian taxa (13 species), Cosmopolitan and Euro-Sirberian taxa (9 species each) and other three categories are poorly represented. From the floristic point of view, Egypt is meeting point of floristic elements belonging to at least four phytogeographical regions, namely; the African Sudano-Zambesian, the Asiatic Irano-Turanian, the Afro-Asiatic Saharo-Sindian and the Euro-Afro-Asiatic Mediterranean (El-Hadidi, 1993). It is clear that in Stellaria community type, the Mediterranean taxa are represented by a relatively high percentage of plant species (53.5%). The other floristic elements are representing by varying numbers of species, reflecting their different capability to penetrate the region. This can be attributed to the influence of human action and the history of agriculture.
Habitat conditions: The data analysis of twenty profiles representing the soil conditions associated with S. pallida are showed in Table 3. The soil was loamy sand in texture with the dominance of fine sand fraction. The percentage of silt was relatively high with mean value of 25.85%. The clay particles ranged between 1.92 and 11.46%. The soil moisture content attained high values and varied from 12.3-22.6%. The soil pore-space ranged between 38.8 and 60.2%. The water holding capacity was also high (42.0-59.7%).
| Table 1: | Floristic composition of twenty stands representing Stellaria pallida (Dumort.) Piré community type |
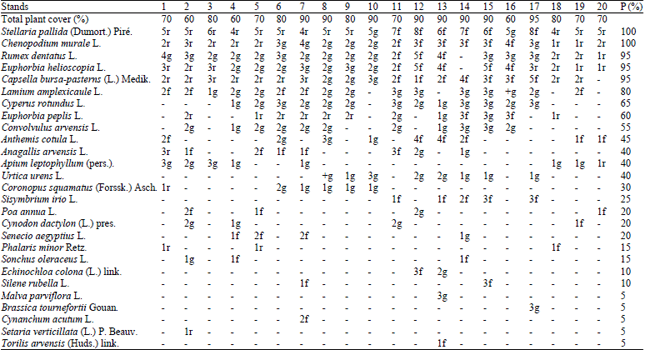 | |
| Domin scale (1-10) g = in foliage, f: Flowering and r: Fruiting, P: Presence (%) | |
| Table 2: | Life-span, life-forms and chorotypes of the floristic components of Stellaria pallida community type |
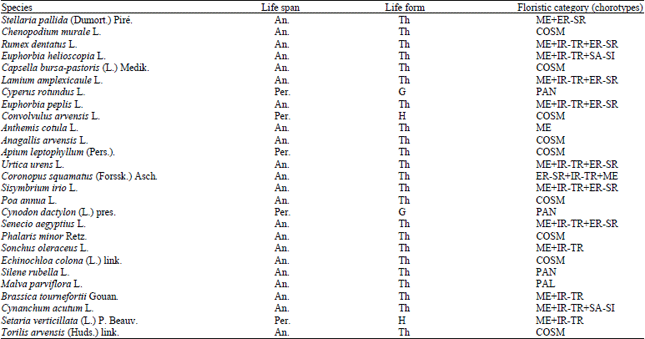 | |
| Life-span: An.: Annuals, Per.: Perennials. Life-form: Th: Therophytes, G: Geophytes, H: Hemicryptophytes. Floristic categories: ME: Mediterranean, ER-SR: Euro-Siberian, COSM: Cosmopolitan, IR-TR: Irano-Turanian, SA-SI: Saharo-Sindian, PAN: Pantropical, PAL: Palaeotropical (2 species = 7.1%) | |
The organic carbon content varied from 0.6-1.6%. CaCO3 content was low and fluctuated between 3.5 and 12.0%. The electric conductivity showed wide range of variation fluctuated between 174 and 374 μmohs cm‾1. Most of the soluble anions were sulphates (0.021-0.086%) and bicarbonates (0.031-0.091%).
| Table 3: | Analysis of soil samples collected from twenty representative stands of Stellaria pallida (Dumort.) Prié. Community inhabiting El Dakahlia Governorate (Jan-Feb, 2014) |
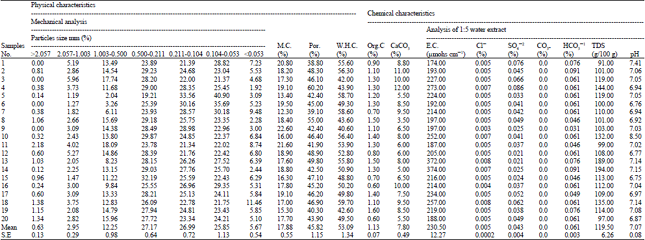 | |
| M.C: Moisture content, Por: Porosity, W.H.C: Water holding capacity, Org. C: Organic carbon, E.C: Electrical conductivity, TDS: Total dissolved salts1 | |
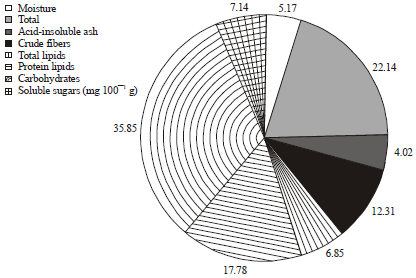 | |
| Fig. 3: | Mean values of the primary constituents of S. pallida aerial parts |
| Table 4: | Qualitative estimation of the secondary metabolites |
 | |
| Table 5: | Quantitative estimation of phenolic and flavonoids compounds |
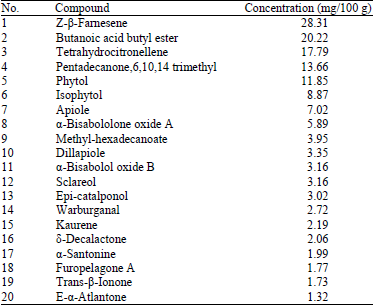
| Table 6: | Phytochemical composition of the volatile oil of Stellaria pallida fruits, determined by GC/MS analysis |
 | |
Chlorides were present in low amounts and varied from 0.003-0.008%. The soluble carbonate content was nil. The soil nature was neutral to slightly alkaline with pH value varied from 6.75-8.5. Consequently, it may be concluded that S. pallida flourished in moist loam sandy soil, non-saline, fertile and slightly-moderately alkaline and this plant is unable to survive drought conditions, thus thrive during the rainy seasons.
The observations, may be give an indication that this Caryophyllaceous herb is well-adapted and thriving in the Mediterranean type of climate which is characterized by a long hot dry season and a short cold mild one. Many species of community are short-lived annuals and restrict germination until mild temperature occur which coincides with water availability from late autumn rains. Temperature is thought to be the main factor affecting the occurrence of these weeds. Comparable plant communities have been described in the Nile Delta by Abu El-Shyokh (2013) and El-Shazly (2013).
Phytochemical characteristics: The data in Fig. 3 showed the primary metabolic constituents and the nutritive value of S. pallida aerial parts. The mean values of: moisture content (5.17%), total ash (22.14%), crude fibers (12.31%), total lipids (6.85%), proteins (17.78%), carbohydrates (35.85%) and soluble sugars (7.14%), with a nutritive value was 276.17 cal./100 g.
Testing of the secondary metabolic products of S. pallida indicates the presence of sterols, terpenes, flavonoids, phenols, tannins and saponins while alkaloids were absent (Table 4). The absence of alkaloids in Stellaria pallida in the present study is in accordance with S. media (Arora and Sharma, 2012).
Polyphenols and flavonoids were detected in S. pallida with mean values of 1.95 and 5.85%, respectively (Table 5).
The yield of colourless volatile oils isolated from flowers of S. pallida was 143 mg/100 g dark-dried powder. Twenty components were identified (Table 6) and Z-B-Farnesene, Butanoic acid butyl ester, Tetrahydrocitronellene, Pentadecanone, Phytol and Apiole were the most important compounds. Balamurugan et al. (2013) reported that Caryophyllaceae is best known for the volatile oils common to many members and many biologically active oils have been isolated from various species.
From the aforementioned results, it may be concluded that S. pallida has relatively high content of lipids, proteins, soluble sugars and a reasonable nutritive value. Thus it may be useful as a leaf vegetable or as an animal feed. The medicinal value of this caryophyllaceous plant lies in the bioactive phytochemical constituents such as sterols, terpene, flavonoids and volatile oils.
REFERENCES
- Adams, R.P., 2007. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry. 4th Edn., Allured Publishing Co., Carol Stream, IL., USA, ISBN-13: 9781932633214, Pages: 804.
Direct Link - AOAC., 2000. Official Methods of Analysis of AOAC International. 17th Edn., Association of Official Analytical Chemists, Gaithersburg, Maryland.
Direct Link - Arora, D. and A. Sharma, 2012. Pharmacognostic and phytochemical studies of Stellaria media Linn. J. Pharm. Sci. Res., 4: 1819-1822.
Direct Link - Balamurugan, K., G. Sakthidevi and V.R. Mohan, 2013. Stimulatory effect of the ethanol extract of Melastoma malabathricum L. (Melastomataceae) leaf on the reproductive system of male albino rats. J. Applied Pharm. Sci., 3: 160-165.
Direct Link - Boulos, L., 1995. Flora of Egypt, Checklist. 1st Edn., Al-Hadara Publishing, Cairo, Egypt, Pages: 283.
Direct Link - Chinnappa, C.C. and J.K. Morton, 1984. Studies on the Stellaria longipes complex (caryophyllaceae)-biosystematics. Syst. Bot., 9: 60-73.
Direct Link - Ghaffari, S.M. and K. Kelich, 2006. New or rare chromosome counts of some angiosperm species from Iran II. Iran. J. Bot., 11: 81-86.
Direct Link - Van Handel, E., 1968. Direct microdetermination of sucrose. Anal. Biochem., 22: 280-283.
CrossRefDirect Link - Indrayan, A. K., S. Sharma, D. Durgapal, N. Kumar and M. Kumar, 2005. Determination of nutritive value and analysis of mineral elements for some medicinally valued plants from Uttaranchal. Curr. Sci., 89: 1252-1255.
Direct Link - Ma, L., J. Song, Y. Shi, C. Wang, B. Chen, D. Xie and X. Jia, 2012. Anti-Hepatitis B virus activity of chickweed [Stellaria media (L.) Vill.] extracts in HepG2. 2.15 cells. Molecules, 17: 8633-8646.
CrossRefDirect Link - Mamadalieva, N.Z., N.T. Ul'chenko, N.K. Yuldasheva, D.R. Egamberdieva, A.A. Zhanibekov, M.K. Dzhukharova and A.I. Glushenkova, 2010. Fatty-acid composition and antibacterial activity of CHCl3 extracts of three plants of the genus Silene. Chem. Nat. Comp., 46: 95-96.
CrossRefDirect Link - Shan, Y., J. Zhou, H.G. Zhao, X. Feng, Y. Dong and B. Xia, 2010. Amino-acid and mineral composition of Stellaria media. Chem. Nat. Comp., 46: 667-668.
CrossRefDirect Link - Sindhu, S. and S. Manorama, 2012. Antimicrobial activity of Polycarpaea corymbosa Lam. (Caryophyllaceae) against human pathogens. J. Chem. Pharm. Res., 4: 4014-4019.
Direct Link - Wittig, R., 2004. The origin and development of the urban flora of central Europe. Urban Ecosyst., 7: 323-339.
CrossRefDirect Link