
Stella O. Olubodun
Department of Medical Biochemistry, School of Basic Medical Sciences, University of Benin, Nigeria
George E. Eriyamremu
Department of Biochemistry, Faculty of Life Sciences, University of Benin, Nigeria
Journal of Biological Sciences
Year: 2015 | Volume: 15 | Issue: 3 | Page No.: 150-155







ABSTRACT
Bean seedlings were used to study carbohydrates (glucose and starch) and Glucose-6-Phosphate Dehydrogenase (G6PD) activity in bean roots exposed to different crude oil fractions [Whole Crude (WC), Water Soluble Fraction (WSF) and Water Insoluble Fraction (WIF)] at different percentages of soil contamination. Equal amount of seedlings in each group were harvested after 7, 14 or 21 days post germination and used for analysis. Descriptive statistics and analysis of variance were employed to statistically analyze data that were obtained. The study reveals higher glucose and Glucose-6-Phosphate Dehydrogenase (G6PD) activity in the WSF than in whole crude oil (WC) and WIF during 21 DPG when compared with control. Greater decrease in starch was observed in WSF than in WC and WIF at 21 DPG when compared with control. The decline of growth observed in the reduction in the number of leaves, root length and plant height observed in WC, WSF and WIF when compared with control may be related to the carbohydrates and glucose-6-phosphate dehydrogenase activity in bean roots.
PDF Abstract XML References Citation
Received: July 28, 2015;
Accepted: September 15, 2015;
Published: October 15, 2015
How to cite this article
Stella O. Olubodun and George E. Eriyamremu, 2015. Glucose-6-Phosphate Dehydrogenase and Carbohydrate in Bean (Vigna unguiculata) Exposed to Crude Oil. Journal of Biological Sciences, 15: 150-155.
DOI: 10.3923/jbs.2015.150.155
URL: https://scialert.net/abstract/?doi=jbs.2015.150.155
DOI: 10.3923/jbs.2015.150.155
URL: https://scialert.net/abstract/?doi=jbs.2015.150.155
INTRODUCTION
One major factor apart from plant nutrient that affects plant growth and productivity is environmental stressors. Environmental stressors may include but not limited to, salt stress, drought stress, heavy metal stress and crude oil stress (Liu et al., 2007; Chugh et al., 2011; Wuana et al., 2013; Olubodun and Eriyamremu, 2013).
Crude oil stress has been shown to affects almost all the physiological and biochemical functions of a plant (Ekpo et al., 2012; Olubodun and Eriyamremu, 2013). Crude oil-induced damage is due mostly to osmotic stress (Ekpo et al., 2012), reduction of mineral element (Odjegba and Sadiq, 2002), reduced availability of mineral elements (Schwab and Banks, 1999) and oxidative stress (Olubodun and Eriyamremu, 2013) as a result of over production of Reactive Oxygen Species (ROS).
Reactive Oxygen Species (ROS) is used to describe highly reactive and partially reduced-oxygen forms which include oxidants or oxygen-radicals, such as superoxide anion (O2·-), hydroxyl radical (OH·), perhydroxyl radical (HO2·) and hydrogen peroxide (H2O2) (Desikan et al., 2005).
Even though ROS are produced during normal metabolic processes, a high concentration of ROS brought about as a result of environmental stressors (drought, crude oil spill), can lead to non-controlled oxidation in cells (oxidative stress) (Desikan et al., 2005; Olubodun and Eriyamremu, 2013). Oxidative stress is an imbalance between the production of ROS and antioxidant enzyme and non-enzyme defense system. Reactive Oxygen Species attack (ROS-attack) can cause damage to cellular macromolecules, such as proteins (protein oxidation), membrane lipids (lipid peroxidation), carbohydrates and DNA (Mittler et al., 2004; Gill and Tuteja, 2010; Olubodun and Eriyamremu, 2013). An increase in oxidative stress due to over production of ROS or decrease in enzymatic and non-enzymatic defense mechanisms may ultimately leads to cell death (Desikan et al., 2005; Gill and Tuteja, 2010). In plants, the links between ROS production and photosynthetic metabolism has led to the selection of antioxidative defence mechanisms, which place all aerobic cells in a state of pro-oxidant or antioxidant balance that prevents cellular damage. Also, the control of the levels of endogenous ROS has opened the evolutionary possibility of using ROS as signalling and effector molecules in growth, development, stress responses and biotic interactions (Mittler et al., 2004; Couee et al., 2006).
The Oxidative Pentose Phosphate Pathway (OPPP) is the main pathway of production of NADPH for biosynthesis (Hutchings et al., 2005) and for redox balance of plant cells (Scheibe, 2004).
Glucose-6-Phosphate Dehydrogenase (G6PD; EC 1.1.1.49) is a homotetramer with subunits of 50-60 kDa and is the rate-limiting enzyme of the PPP. Glucose-6-phosphate dehydrogenase is required for the antioxidant defense system as it produces the cell’s principal reductant, NADPH (Zhang et al., 2000; Wakao et al., 2008; Cardi et al., 2011). Report has shown that G6PD is the principal source of NADPH utilized in redox regulation. Many studies have shown that G6PD and the OPPP increase under oxidative stress conditions (Zhang et al., 2000; Liu et al., 2007; Wakao et al., 2008; Cardi et al., 2011; Dal Santo et al., 2012). Other researches in different cell types has also shown that slight changes in G6PD activity have significant effects on cell growth and cell death (Tian et al., 1999; Zhang et al., 2000). The activity of G6PD is therefore required for adequate defense against oxidative stress and prevention of cell damage or cell death.
Studies have shown that carbohydrates (soluble sugars) is not only involved in, or related to, ROS-producing metabolic pathways but can also feed NADPH-producing metabolic pathways (OPPP), which can contribute to ROS scavenging. In yeasts and mammalian cells, high glucose has been associated with toxicity and pathogenesis through increased production of ROS by auto-oxidation of glucose and glucose metabolism (Couee et al., 2006). By contrast, feeding glucose into the OPPP can enhance NADPH production, which is a major cofactor of ROS scavenging pathways such as ascorbate-glutathione cycles. In some mammalian cell types, glucose has been shown to enhance cellular defences against cytotoxicity of hydrogen peroxide (Couee et al., 2006). However, another report suggested that high glucose predisposes cells to oxidative damage because of inadequate activation of the OPPP after exposure to an oxidant (Zhang et al., 2000). The study is thus aimed at investigating the effect of different crude oil fractions on the carbohydrates and activity of G6PD in the roots of bean seedlings.
MATERIALS AND METHODS
Study location: The ex situ study was carried out at the University of Benin, Benin City, Edo State, Nigeria. The region is made up of flat plains with sandy loam soil characterized as coastal plain sand.
| Table 1: | Concentration of crude oil contamination in soil |
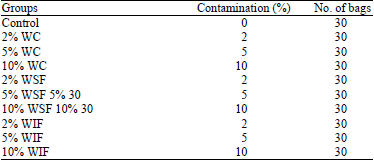 | |
| WC: Whole crude, WSF: water soluble fraction, WIF: Water insoluble fraction | |
The region experiences relatively constant temperature and humidity throughout the year with annual rainfall of about 2673.8 mm.
Experimental design: Sandy loam soil of pH 6.15 was weighed into 120 polythene bags such that each bag contained 500 g soil and grouped as shown in Table 1.
Plant materials: Bean (Vigna unguiculata) seeds of variety ITA 189-288 were bought from a local market in Benin City, Nigeria. Viability of the seeds was assessed using floatation method. The seeds were placed in a beaker containing tap water and stirred. The viable seeds represent the seeds that did not float.
Crude oil and fractionation: Crude oil (Bonny light, oAPI (American Petroleum Institute) gravity = 37) was obtained from Warri Refinery and Petrochemical Company, Delta State, Nigeria. A portion of the crude oil was fractionated by the method of Anderson et al. (1974) into Water Soluble Fraction (WSF) and Water Insoluble Fraction (WIF). For the fractionation, a 1:2 dilution of 200 mL of crude oil was put in a 1 L conical flask and constantly stirred with a magnetic stirrer for 48 h. The WSF then separated from the WIF in a separating funnel.
Soil treatment: Control soil contained only sandy loam soil. The soil in the bags contaminated with Whole Crude (WC), Water Soluble Fraction (WSF) and Water Insoluble Fractions (WIF) were mixed thoroughly in their respective polythene bags containing 500 g sandy loam soil with the aid of a plastic spade. Soil of 500 g was treated with 10, 25 and 50 mL of crude oil to obtain 2, 5 and 10% v/w crude oil contamination.
Planting of seeds and seedling germination studies: The seeds were planted by a modified version of Vavrek and Campbell (2002). Three viable bean seeds were sown in 500 g sandy loam soil with a depth of about 1-2 cm. The time and number of seeds that spouted from each bag were noted and the percentage seedling germination in each treatment was calculated using the equation:
Equal amounts of seeds that germinated were harvested at 7, 14 and 21 day. Growth parameters were assessed after each harvest.
Biochemical assays: Glucose, starch and glucose-6-phosphate dehydrogenase were also assessed in the roots of the bean plant after each harvest.
Enzyme extraction: Glucose-6-phosphate dehydrogenase enzyme extraction was done on ice using a modified method described by Esposito et al. (2001). Glucose-6-phosphate dehydrogenase was extracted by homogenizing (0.5 g) of the roots in buffer A (50 mM Tris-Phosphate, pH 8.0, 10 mM MgCl2, 10% glycerol, 4 mM EDTA, 15% l M NADPH), supplemented with 1 L mL–1 Plant Protease Inhibitor. All homogenates were centrifuged at 5,000x g for 20 min and the supernatant defined as crude extract was used to assay for the enzyme, G6PD. (modified Esposito et al., 2001). All experiments were performed in triplicate.
Enzymes activity: The activities of Glucose-6-Phosphate Dehydrogenase (G6PD) were determined with the Randox test kit (Randox Laboratories Ltd., 55 Diamond Road, Crumlin, Co. Antrim, United Kingdom, BT29 4QY). All experiments were performed in three replicate.
Enzyme activity was determined by measuring the rate of absorbance change at 340 nm, due to the reduction of NADP+ to NADPH when the plant extract was incubated with Glucose-6-Phosphate (G6P). Activity of glucose 6 phosphate dehydrogenase was assigned by measuring the amount of NADP reduced per minute per mg protein.
Calculation: The G6PD activity (mU/sample per mL) = 33650×Δ A 340 nm min–1.
Determination of starch and glucose: Starch determination was carried out after enzymatic hydrolysis into glucose. Starch and glucose were estimated by the RANDOX test kit (Randox Laboratories Ltd., 55 Diamond Road, Crumlin, Co. Antrim, United Kingdom, BT29 4QY), according to the manufacturer’s instructions. Bio-molecular activities, based on enzyme-linked formation of NADPH, were expressed as a specific activity measured at 570 nm. The corresponding concentration was determined against hydrogen peroxide standard curve prepared by using hydrogen peroxide solution. The amount of sugar was expressed as mg g–1 fresh weight and all experiments were performed in triplicate.
Statistical analysis: The result of the study was expressed as Mean±standard Error of Mean (SEM). Analysis of variance was used to test for differences in the groups while Duncan’s multiple comparisons test was used to determine significant differences between means. The Instat-Graphpad software, San Diego, California, USA, was used for this analysis. A p<0.05 was considered statistically significant.
RESULTS
The percentage germination of the bean seeds for each of the fractions used decreased as the percentage contamination increased. The seeds grown on 10% Water Insoluble Fraction (WIF) contaminated soil exhibited the lowest germination rate of 27% while the highest germination rate was observed in the soil contaminated with 2% water soluble fraction (Table 2). The Leave Number (LN) of bean seeds grown in the different treatment groups did not significantly differ from control after 7 Days Post Germination (DPG). However, the LN in the test groups significantly reduced (p<0.05) after 14 and 21 DPG, with 10% WIF showing the greatest reduction. Plant Height (PH) and Root Length (RL) reduced for the various fractions used in a nearly percentage contamination dependent manner. For all the growth parameters assessed, 2% Whole Crude (WC) and Water Soluble Fraction (WSF) appeared to exert the least negative effects, whereas 10% WIF exerted the most negative effects.
| Table 2: | Effect of different fractions of crude oil contamination on percentage germination and growth parameters of bean seeds |
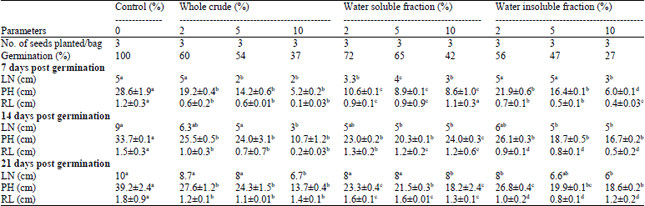 | |
| PH: Plant height, RL: Root length, LN: Number of leaves, values are means of 3-5 determinations±SEM. Means carrying different notations are statistically different at p<0.05 using Instat graph pad | |
| Table 3: | Effects of different concentrations of crude oil and its fractions on starch content in the roots of bean (mg g–1) |
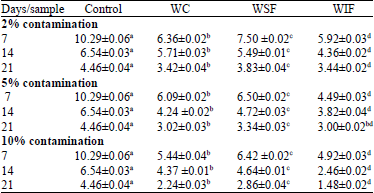 | |
| Values are mean of three (n = 3) replicates±SEM, WC: Whole crude, WSF: Water soluble fraction of crude oil, WIF: Water insoluble fraction of crude oil, starch contents = mg g–1 wet tissue, Means of the same row carrying different notations are statistically different at p<0.05 using Instat graphpad | |
| Table 4: | Effects of different concentrations of crude oil and its fractions on glucose content in the roots of bean (mg g–1) |
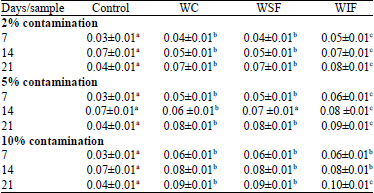 | |
| Values are mean of three (n = 3) replicates±SEM, WC: Whole crude, WSF: Water soluble fraction of crude oil, WIF: Water insoluble fraction of crude oil, glucose contents = mg g–1 wet tissue, Means of the same row carrying different notations are statistically different at p<0.05 using Instat graphpad | |
The results of the effects of different concentration of crude oil and its fractions on starch and glucose contents of bean roots are presented in Table 3 and 4. The results indicated reduction in starch contents and increase in glucose content in the crude oil exposed plant when compared with control.
The results of Glucose-6-Phosphate Dehydrogenase (G6PD) activity are presented in Table 5. The results showed that the activity of G6PD in the bean roots exposed to crude oil was significantly increased when compared with control, the WSF having the greatest increase.
DISCUSSION
Crude oil has been reported to be increasingly deleterious to soil, animals and plants (Onuh et al., 2008; Olubodun and Eriyamremu, 2013). This study revealed that percentage seed germination and the growth parameters of the bean seedlings were adversely affected by crude oil contamination and these negative effects increased as the percentage contamination of the soil increased. Various studies have also reported decreased crop production and greater damage to crops with increasing concentration of crude oil spillage.
| Table 5: | Effects of different concentrations of crude oil and its fractions on Glucose-6-phosphate dehydrogenase activities in bean roots (µmole of NADPH utilized min–1 mg–1 protein) |
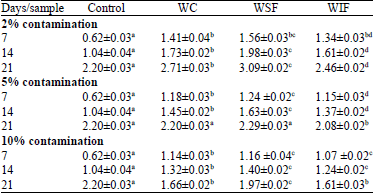 | |
| Values are mean of three (n = 3) replicates±SEM, WC: Whole crude, WSF: Water soluble fraction of crude oil, WIF: Water insoluble fraction of crude oil, glucose contents = mg g–1 wet tissue, Means of the same row carrying different notations are statistically different at p<0.05 using Instat graphpad | |
Several researchers reported delayed germination and reduction in crop growth in different contaminated soils and plant species (Alves et al., 2012; Odjegba, 2013; Olubodun and Eriyamremu, 2013; Omosun et al., 2008). Growth reduction of plants as a result of crude oil contamination in the soil have been attributed to insufficient aeration and limited water necessary for plant growth (Njoku et al., 2008; Odjegba, 2013; Olubodun and Eriyamremu, 2013). Since the water contains dissolved nutrients, the reduction in root length observed in the study, may have limited the quantity of nutrients the plant is able to absorb. Also, the reduction in leaf number may reduce the rate of photosynthesis which invariables may have had the cumulative effect of reduction of plant growth. The water insoluble fractions, which may have further limited the availability of water soluble nutrients in the soil, could account for the greater reduction in growth parameters observed in bean seeds sown in soil contaminated with WIF especially at the highest concentration used (10%). This agrees with the observation of Olubodun and Eriyamremu (2013).
The reduction in starch contents observed when compared with control is in agreement with that of Dolferus et al. (2001), who demonstrated that reduction in oxygen supply lead to 18 fold reduction in adenosine triphosphate (ATP) production and plants had to compensated for this loss by accelerating sugar metabolism and glycolysis. The elevated glucose content in the crude oil exposed plant when compared with control also agrees with earlier reports (Dolferus et al., 2001). This is further confirmed by other reports that, total carbohydrates and proline content were, decreased with application of zinc in Salvia officinalis L. plants (Hendawy and Khalid, 2005; Cha-Um et al., 2009). The increased glucose contents observed in crude oil exposed root compares favourably with decrease in lipid and increase in glucose contents previously reported (Boopathy, 2000; Amora-Lazcano et al., 2010). This may indicate that starch was the main source of energy in the plant during crude oil exposure which may be related to the fact that starch can be converted into glucose under stress condition as starch (storage carbohydrate) are dissociated as primary metabolites in stress condition to overcome the energy demand by the plant for growth. The decrease in starch contents may be related with the toxicity of crude oil because starch helps the plant in giving energy at the time of emergency for survival. The results suggest that crude oil inhibited the growth of the plant possibly because of the decrease in starch contents while the high glucose contents may indicate high energy demand to overcome the stress of growing as well as crude oil toxicity. The results also suggest that the glycolytic pathway was active (high glucose 6 phosphate dehydrogenase activity) and that starch could be used by the growing plants to produce glucose as energy source to sustain the metabolic activities occurring in the plant.
When mitochondria respiration capacity decreases, secondary pentose phosphate pathway can be activated to compensate for the failure in energy supply (Smiri et al., 2009). Reducing power can be supplied through secondary NADPH-recycling dehydrogenase activities, such as G6PD (Smiri et al., 2009). Since glucose-6-phosphate dehydrogenase is the rate-limiting enzyme of OPPP and determines the level of NADPH, the metabolism of glucose through the OPPP can be controlled (Kletzien et al., 1994). Increased G-6-PD activity recorded in this study agrees with that of Liu et al. (2007). However, Liu et al. (2007) reported decreased in activity after certain levels of contamination. The increase in G-6-PD activity may have enhanced NADPH production in the roots of bean and may have resulted in the tolerance of the plant roots to crude oil stress. The increase of NADPH contents may have stimulated production of the nitrate reductase-dependent Nitric Oxide (NO) (Liu et al., 2007). Nitric oxide has been shown to act as a growth regulator which could induce seed germination, affect growth and development of plant tissue, increase iron homeostasis and accelerate plant cell. Nitric oxide was also suggested to be involved in resistant responses to various biotic and abiotic stressors senescence (Liu et al., 2007). The increase in G-6-PD activity may have increased NO production in the roots of the plants under crude oil stress. This may have further stimulated the increase in the activities of antioxidant enzymes observed in an earlier report (Olubodun and Eriyamremu, 2013). Enhanced antioxidant enzyme activities may have helped to scavenge the ROS produced due to crude oil exposure. Glucose-6-phosphate dehydrogenase activity may have played a key role in protecting the roots of bean against oxidative stress induced by crude oil exposure and establish tolerance of the roots to crude oil stress (Liu et al., 2007).
CONCLUSION
This study has shown that the different fractions of crude oil contaminated soil significantly reduced several growth parameters of bean and induced oxidative stress in the bean root. We suggest that carbohydrate and glucose-6-phosphate dehydrogenase activity may have played a key role in protecting the roots of bean against oxidative stress induced by crude oil exposure and establish tolerance of bean roots to crude oil stress.
ACKNOWLEDGMENTS
We thank Mr. Kingsley Ikeke of the Department of Science Laboratory Technology, Edo State Institute of Technology and Management, Usen for helping to transport some of our samples for analysis. George E. Eriyamremu developed the concepts and designed the experiments. This work was carried out in collaboration between both authors. Both authors read and approved the manuscript.
REFERENCES
- Alves, G.A.R., B.G. dos Santos Filho, A.K. da Silva Lobato, D.K.Y. Tan and C.F. de Oliveira Neto et al., 2012. Water relations, nitrogen compounds and enzyme activities in leaf and root of young Yellow Lapacho (Tabebuia serratifolia) plants subjected to flooding. Plant Omics J., 5: 216-222.
Direct Link - Amora-Lazcano, E., L.A. Guerrero-Zuniga, A. Rodriguez-Tovar, A. Rodriguez-Dorantes and M.S.Vasquez-Murrieta, 2010. Rhizospheric Plant-Microbe Interactions that Enhance the Remediation of Contaminated Soils. In: Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology, Mendez-Vilas, A. (Ed.). Formatex Research Center, Badajoz, pp: 251-256.
- Boopathy, R., 2000. Factors limiting bioremediation technologies. Bioresour. Technol., 74: 63-67.
CrossRefDirect Link - Cardi, M., K. Chibani, D. Cafasso, N. Rouhier, J.P. Jacquot and S. Esposito, 2011. Abscisic acid effects on activity and expression of barley (Hordeum vulgare) plastidial glucose-6-phosphate dehydrogenase. J. Exp. Bot., 62: 4013-4023.
CrossRefDirect Link - Cha-Um, S., T. Trakulyingcharoen, P. Smitamana and C. Kirdmanee, 2009. two rice cultivars differing salt tolerant abilities in responses to iso-osmotic stress. Aust. J. Crop Sci., 3: 221-230.
Direct Link - Chugh, V., N. Kaur and A. Gupta, 2011. Evaluation of oxidative stress tolerance in maize (Zea mays L.) seedlings in response to drought. Indian J. Biochem. Biophys., 48: 47-53.
PubMedDirect Link - Couee, I., C. Sulmon, G. Gouesbet and A. El Amrani, 2006. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot., 57: 449-459.
CrossRefDirect Link - Dal Santo, S., H. Stampfl, J. Krasensky, S. Kempa and Y. Gibon et al., 2012. Stress-induced GSK3 regulates the redox stress response by phosphorylating glucose-6-phosphate dehydrogenase in Arabidopsis. Plant Cell, 24: 3380-3392.
CrossRefDirect Link - Dolferus, R., E.J. Klok, K. Ismond, C. Delessert and S. Wilson et al., 2001. Molecular basis of the anaerobic response in plants. IUBMB Life, 51: 79-82.
CrossRefDirect Link - Ekpo, I.A., R.B. Agbor, E.C. Okpako and E.B. Ekanem, 2012. Effect of crude oil polluted soil on germination and growth of soybean (Glycine max). Ann. Biol. Res., 3: 3049-3054.
Direct Link - Esposito, S., S. Carfagna, G. Massaro, V. Vona and V.D.M. Rigano, 2001. Glucose-6-phosphate dehydrogenase in barley roots: Kinetic properties and localisation of the isoforms. Planta, 212: 627-634.
CrossRefDirect Link - Gill, S.S. and N. Tuteja, 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem., 48: 909-930.
CrossRefDirect Link - Hendawy, S.F. and K.A. Khalid, 2005. Response of sage (Salvia officinalis L.) plants to zinc application under different salinity levels. J. Applied Sci. Res., 1: 147-155.
Direct Link - Hutchings, D., S. Rawsthorne and M.J. Emes, 2005. Fatty acid synthesis and the oxidative pentose phosphate pathway in developing embryos of oilseed rape (Brassica napus L.). J. Exp. Bot., 56: 577-585.
CrossRefDirect Link - Kletzien, R.F., P.K. Harris and L.A. Foellmi, 1994. Glucose-6-phosphate dehydrogenase: A housekeeping enzyme subject to tissue-specific regulation by hormones, nutrients and oxidant stress. FASEB J., 8: 174-181.
PubMedDirect Link - Liu, Y., R. Wu, Q. Wan, G. Xie and Y. Bi, 2007. Glucose-6-phosphate dehydrogenase plays a pivotal role in nitric oxide-involved defense against oxidative stress under salt stress in red kidney bean roots. Plant Cell Physiol., 48: 511-522.
CrossRefDirect Link - Mittler, R., S. Vanderauwera, M. Gollery and F. van Breusegem, 2004. Reactive oxygen gene network of plants. Trends Plant Sci., 9: 490-498.
CrossRefPubMedDirect Link - Njoku, K.L., M.O. Akinola and B.O. Oboh, 2008. Growth and performance of Glycine max L. (Merrill) grown in crude oil contaminated soil augmented with cow dung. Nat. Sci., 6: 48-56.
Direct Link - Odjegba, V.J., 2013. Responses of Rhizophora mangle to simulated crude oil pollution. Indian J. Innov. Dev., 2: 839-845.
Direct Link - Odjegba, V.J. and A.O. Sadiq, 2002. Effects of spent engine oil on the growth parameters, chlorophyll and protein levels of Amaranthus hybridus L. Environmentalist, 22: 23-28.
CrossRefDirect Link - Olubodun, S.O. and G.E. Eriyamremu, 2013. Effect of different crude oil fractions on growth and oxidative stress parameters of maize radicle. Int. J. Plant Soil Sci., 2: 144-154.
CrossRefDirect Link - Omosun, G., A.A. Markson and O. Mbanasor, 2008. Growth and anatomy of Amaranthus hybridus as affected by different crude oil concentrations. Am.-Eurasian J. Scient. Res., 32: 70-74.
Direct Link - Onuh, M.O., D.K. Madukwe and G.U. Ohia, 2008. Effects of poultry manure and cow dung on the physical and chemical properties of crude oil polluted soil. Sci. World J., 3: 45-50.
Direct Link - Scheibe, R., 2004. Malate valves to balance cellular energy supply. Physiologia Plantarum, 120: 21-26.
CrossRefDirect Link - Smiri, M., A. Chaoui and E. El Ferjani, 2009. Respiratory metabolism in the embryonic axis of germinating pea seed exposed to cadmium. J. Plant Physiol., 166: 259-269.
CrossRefDirect Link - Tian, W.N., L.D. Braunstein, K. Apse, J. Pang, M. Rose, X. Tian and R.C. Stanton, 1999. Importance of glucose-6-phosphate dehydrogenase activity in cell death. Am. J. Physiol.-Cell Physiol., 276: C1121-C1131.
Direct Link - Wakao, S., C. Andre and C. Benning, 2008. Functional analyses of cytosolic glucose-6-phosphate dehydrogenases and their contribution to seed oil accumulation in arabidopsis. Plant Physiol., 146: 277-288.
Direct Link - Wuana, R.A., P.A. Adie, J. Abah and M.A. Ejeh, 2013. Screening of pearl millet for phytoextraction potential in soil contaminated with cadmium and lead. Int. J. Sci. Technol., 2: 310-319.
Direct Link - Zhang, Z., K. Apse, J. Pang and R.C. Stanton, 2000. High glucose inhibits glucose-6-phosphate dehydrogenase via cAMP in aortic endothelial cells. J. Biol. Chem., 275: 40042-40047.
CrossRefPubMedDirect Link - Vavrek, M.C. and W.J. Campbell, 2002. Identification of plant traits that enhance biodegradation of oil. Proceedings of the 9th Annual International Petroleum Environmental Conference, Albuquerque, NM., October 22-25, 2002, The Integrated Petroleum Environmental Consortium, The University of Tulsa, OK.
- Anderson, J.W., J.M. Neff, B.A. Cox, H.E. Tatem and G.M. Hightower, 1974. Characteristics of dispersions and water-soluble extracts of crude and refined oils and their toxicity to estuarine crustaceans and fish. Mar. Biol., 27: 75-88.
CrossRefDirect Link