
A.K. Singh
Department of Veterinary Parasitology, Vigyan Vishwavidyalaya Evam Go-Anusandhan Sansthan, 281001, Mathura (U.P.), India
A.K. Verma
Department of Veterinary Epidemiology and Preventive Medicine, Vigyan Vishwavidyalaya Evam Go-Anusandhan Sansthan, 281001, Mathura (U.P.), India
Neha
Department of Veterinary Epidemiology and Preventive Medicine, Vigyan Vishwavidyalaya Evam Go-Anusandhan Sansthan, 281001, Mathura (U.P.), India
Ruchi Tiwari
Department of Microbiology and Immunology, Uttar Pradesh Pandit Deen Dayal Upadhayay Pashu Chikitsa Vigyan Vishwavidyalaya Evam Go-Anusandhan Sansthan, 281001, Mathura (U.P.), India
K. Karthik
Division of Bacteriology and Mycology, Vigyan Vishwavidyalaya Evam Go-Anusandhan Sansthan, 281001, Mathura (U.P.), India
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly (U.P.), India
S.V. Singh
Microbiology Laboratory, Animal Health Division, Central Institute for Research on Goats, Mathura, India
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 2 | Page No.: 95-109







ABSTRACT
By 2050 to feed the estimated human population of around 9 billion, there is requirement of 50% increase the food production, which can only be fulfilled by clean, healthy and sustainable food animal production. Livestock industry is facing considerable economic losses due to infectious diseases. So an effective control strategy is need of today to control these infectious diseases and contribute in augmentation of livestock production. Parasitic diseases have a major impact on livestock production, reproduction and hence economy. Protozoan parasites are major causes of human and animal disease causing extensive morbidity and mortality, particularly parasitic disease in tropical and sub-tropical climatic regions. Many protozoal parasitic diseases are zoonotic. Limiting the impact of parasitism in both man and livestock relies almost exclusively on the use of antiparasitic drugs. Development of resistance towards chemotherapeutic agents has forced the scientist to discover some alternative for control of parasitic diseases. Recent advances in immunology and biotechnology have sensitized the scientists or researchers to develop the newer and safer vaccines for control of parasitic diseases. This review is intended to provide state-of-art information to the reader with an overview on the trends, advances and perspectives in vaccines and vaccinology against important parasitic diseases of livestock and poultry viz., coccidiosis, anaplasmosis, giardiosis, babesiosis, Neospora infection, toxoplasmosis, theleriosis, sarcocyst infestation, leishmaniasis, trypanosomiasis and trichomoniasis, which altogether play crucial role in the prevention of protozoan parasitic diseases of animals.
PDF Abstract XML References Citation
Received: October 08, 2013;
Accepted: October 31, 2013;
Published: January 11, 2014
How to cite this article
A.K. Singh, A.K. Verma, Neha, Ruchi Tiwari, K. Karthik, Kuldeep Dhama and S.V. Singh, 2014. Trends and Advances in Vaccines Against Protozoan Parasites of Veterinary Importance: A Review. Journal of Biological Sciences, 14: 95-109.
DOI: 10.3923/jbs.2014.95.109
URL: https://scialert.net/abstract/?doi=jbs.2014.95.109
DOI: 10.3923/jbs.2014.95.109
URL: https://scialert.net/abstract/?doi=jbs.2014.95.109
INTRODUCTION
According to an estimate, by 2050 to feed the estimated human population of around 9 billion, there is requirement of 50% increase the food production. This requirement can only be fulfilled by clean, healthy and sustainable food animal production (Mahima et al., 2012a; Fitzpatrick, 2013). In modern days, the livestock industry is facing considerable economic losses due to infectious diseases of viral, bacterial, fungal, parasitic origin (Rogers and Randolph, 2006; Jones et al., 2008; Cascio et al., 2011; Verma et al., 2008, 2012a, b; Kumar et al., 2011; Dhama et al., 2013a, b). So an effective control strategy is need of today to control these infectious diseases and contribute in augmentation of livestock production (Innes et al., 2011; Dhama et al., 2013c, d). In past or till now, the limiting of impact of parasitism relies on use of chemotherapy like anthelminthic, antiprotozoal drugs etc (Vercruysse et al., 2004). Due to development of resistance towards these chemotherapeutic agents, scientists or researchers are now thinking towards the prevention of parasitic diseases through use of vaccines (Barriga, 1994; Vercruysse et al., 2004; Stern and Markel, 2005; Innes and Vermeulen, 2006; Paul-Pierre, 2009; Knox, 2010; Sharman et al., 2010; Liu et al., 2012; Dhama et al., 2008, 2013e). All these facts have forced the researchers to develop some alternatives for prevention and control measures like effective vaccination, immunomodulators and novel therapeutic regimens (Meeusen et al., 2007; Vercruysse et al., 2007; Manzano-Roman et al., 2012; Dhama et al., 2013d; Jacob et al., 2013; Mahima et al., 2012b). Effective ways to combat parasites are very limited due to their complex nature and complicated relationship with the hosts. Vaccine is a substance used to stimulate the production of antibodies and provide immunity against one or more diseases. It is prepared from causative agent of disease, its products, treated to act as antigen without causing disease (Stern and Markel, 2005). With the development of immunology, a continuous flow of vaccines to the market is going on, but among them the percentage of parasitic vaccine is very low (Vercruysse et al., 2004). The development of vaccines against the parasitic diseases for domestic animals is most fascinating, promising and challenging field (Innes and Vermeulen, 2006). Majority of the problems are due to their complex life cycle and difficulty in their in vitro culture (Cornelissen and Schetters, 1996).
Parasitic diseases have a major impact on livestock production worldwide with infection arising from a range of helminth, protozoan and ecto-parasites and out of these, many parasitic diseases are zoonotic (Innes and Vermeulen, 2006; Cornelissen and Schetters, 1996; Paul-Pierre, 2009; Dhama et al., 2013a, b, c, f; Reichel et al., 2013). This review is intended to provide state-of-art information to the reader with an overview of vaccines available against parasitic diseases of livestock viz., coccidiosis, anaplasmosis, giardiosis, babesiosis, Neospora infection, toxoplasmosis, theleriosis, sarcocyst infestation, leishmaniasis, trypanosomiasis and trichomoniasis, which are very useful for prevention and control of parasitic diseases in animals.
PROTOZOAN DISEASES
Protozoa are unicellular, eukaryotic, microscopic organisms, belongs to subkingdom protozoa, having a distinct nucleus as well as endoplasmic reticulum, golgi apparatus, mitochondria in the cytoplasm. About 65,000 species of protozoa have so far been named of which a great majority are free-living, while only 7000 protozoan species are parasitic both in vertebrate and invertebrate animals (Levine, 1985).
Metazoan and protozoan parasites are one of the major causes of disease causing extensive morbidity and mortality in animals (Knox, 2010). These parasites are major impediment to the introduction of high-productivity breeds in poorer or developing countries. Among these many of the parasites are zoonotic in nature that also increases their economic importance. At present, no vaccine for human protozoal disease is available; however, several veterinary vaccines are available in the market (Meeusen et al., 2007; Vercruysse et al., 2007). Among vaccines against protozoan diseases of livestock, many are based on live organisms, but recently there is progress in development and commercialization of killed subunit vaccines (Lightowlers, 1994; Sharman et al., 2010).
ANTIPROTOZOAN DRUGS- PROBLEMS IN THE NEAR FUTURE
Drugs against protozoan parasites are widely used to prevent the infection but protozoans are able to develop resistance against these drugs, which is a serious problem in human and veterinary (Vercruysse et al., 2007). Broiler industry is facing the crisis of antiprotozoan drug resistance against the common coccidiostats used in the farm (Stephan et al., 1997). This has lead to the shuttling of drugs used against Eimeria so as to keep resistance development under check. Resistances against antiprotozoan drugs are now reported in Trypanasoma (Geerts et al., 2001) and also in canine Babesiosis (Collett, 2000). Extreme use of antiprotozoan drugs has not only lead to the resistance against these drugs but also the residues in the animal products enter the food chain causing cross transfer of resistance (De Ruyck et al., 2000; Geary, 2002). Apart from these problems these drugs can also enter environment from the secretions and excretions of the animals, threatening the public further (Steel and Wardhaugh, 2002). Hence the current regime for the treatment several protozoan infections are not proper and the search for new antiprotozoan drugs has not yielded a fruitful result (Von Samson-Himmelstjerna and Blackhall, 2005). To solve these problems vaccination will be a good means to control the disease.
VACCINE AND ITS TYPES
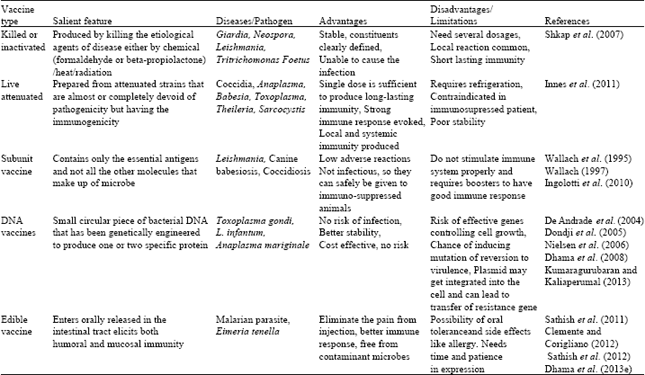
Vaccines are dead or inactivated organisms or purified products derived from them. There are several types of vaccines in use (Table 1):
| • | Killed or inactivated |
| • | Live attenuated |
| • | Subunit |
| • | DNA Vaccine |
| • | Edible vaccine |
Killed or inactivated: It is produced by killing the etiological agents of disease either by chemical (formaldehyde or beta-propiolactone)/heat/radiation and such vaccines are more safe and stable than live vaccines.
| Table 1: | Types of protozoan vaccines |
 | |
These are inactivated vaccines and easy to prepare. In this type of vaccine, the replicative function of etiological agent should be destroyed, while the outer coat of agent should be left intact. For its effectiveness, large amount of antigen is required in comparison to live vaccine. Excessive treatment can destroy immunogenicity whereas insufficient treatment can leave infectious agent capable of causing disease. The commercially available killed/subunit vaccines available against protozoan parasites are given in Table 2.
Advantages:
| • | Killed vaccines are stable |
| • | Constituents clearly defined |
| • | Unable to cause the infection |
Disadvantages:
| • | Need several dosages |
| • | Local reaction common |
| • | Short lasting immunity (Shkap et al., 2007) |
Live attenuated: Live vaccines are prepared from attenuated strains that are almost or completely devoid of pathogenicity but having the immunogenicity therefore they are capable of inducing a protective immune response. They multiply in the host and provide continuous antigenic stimulation over a period of time. The commercially available live protozoal vaccine against animal diseases is given in Table 2.
Advantages:
| • | Single dose is sufficient to produce long-lasting immunity |
| • | Strong immune response evoked |
| • | Local and systemic immunity produced (Innes et al., 2011) |
Disadvantages:
| • | It required refrigeration |
| • | Contraindicated in immunosupressed patient |
| • | Poor stability |
Subunit vaccine: Instead of entire microbe, subunit vaccines include only that best antigen which stimulates the immune system. It contains only the essential antigens and not all the other molecules that make up of microbe. It is of three types: (a) Natural tissue purified protein, (b) Recombinant protein antigens and (c) Chemical small peptide vaccines. Two subunit vaccines are available in market for canine babesiosis caused by B. canis. These vaccines contain soluble antigens from the pathogen of canine babesiosis.
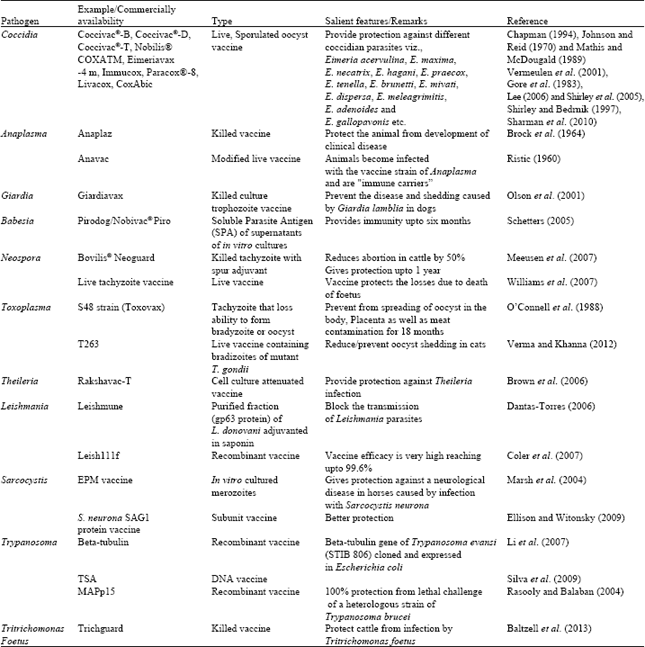
| Table 2: | Progress in protozoan vaccines and their commercial availability |
 | |
NobivacPiro® is a newly marketed vaccine which contains soluble protein antigen of two pathogens namely B. canis and Babesia rossi that broadens the immunity. ABIC Veterinary Products, Israel markets a novel killed subunit vaccine for poultry coccidiosis (Wallach, 1997). The peculiarity of this vaccine is that it targets macrogametocyte stages of coccidia which results in formation of oocyst unlike live vaccines which target merozoite stages. Laying hens are immunized by this vaccine rather than chicks, transferring immunoglobulins to the yolk which is the added advantage of this vaccine (Wallach et al., 1995). A subunit vaccine comprising of antigens such as Gam82 and others purified from the Eimeria gametocytes has been reported to be safer and effective in preventing coccidiosis in birds (Vermeulen, 1998; Wallach et al., 2008; Jang et al., 2010).
Advantage:
| • | Low adverse reactions |
| • | Not infectious, so they can safely be given to immuno-suppressed animals |
Disadvantage:
| • | As native structure of antigens is difficult to retain, so antibodies produced after immunization may not recognize the same protein on the pathogen surface |
| • | Do not stimulate immune system properly and requires boosters to have good immune response (Ingolotti et al., 2010) |
DNA vaccines: These vaccines are the third generation vaccines, in which DNA responsible for particular protein is directly injected into host to produce the desired immune response. At present this technology is proving helpful in control of protozoan infections (Dumonteil, 2007; Liu et al., 2006). DNA vaccines are developed for a handful of protozoan infections of animals and their studies on laboratory animals showed good results that these vaccines can be adopted for animals (Da’dara and Harn, 2005; Dhama et al., 2008; Carvalho et al., 2010; Kumaragurubaran and Kaliaperumal, 2013).
DNA vaccine for Toxoplasma gondi targeting the bradyzoite stage antigens BAG1 and MAG1 reduced cyst burden in mice when challenged with infection (Nielsen et al., 2006). Recombinant plasmid using T. gondii surface antigen 1 (SAG1) and 14-3-3 protein has been found to induce protective immunity in BALB/c mice (Meng et al., 2012). Recently, a DNA vaccine expressing eukaryotic translation initiation factor (eIF4A) protein of T. gondii has been reported to be promising in inducing protective immunity against acute toxoplasmosis in mice (Chen et al., 2013). Development of DNA vaccines has been tried against protozoan parasites, Leishmania and Trypanosoma cruzi also which affects human being. Leishmaniasis, a major public health zoonoses, is a complex disease caused by at least 18 species of genus Leishmania transmitted by hematophagous sandflies while T. cruzi is the causative agent of Chagas disease. Cross-protection studies showed that infection by one species of Leishmania may or may not protect from subsequent infection by another species of same genera (Xu and Liew, 1994, 1995). In dogs, therapeutic DNA vaccine has also been developed against Trypanosoma cruzi infection (Quijano-Hernandez et al., 2008). Another DNA vaccine targeting the Leishmania infantum acidic ribosomal protein P0 (LiPO), Leishmania major DNA vaccine encoding PSA-2 antigen, Cysteine Proteinase (CP) a and b DNA vaccines also had good immune response studies in mice (Dondji et al., 2005). Another DNA vaccine encoding L. donovani nucleoside hydrolase NH36 is therapeutically active against murine visceral leishmaniasis produced by L. chagasi (Handman et al., 2000; Aguilar-Be et al., 2005; Gamboa-Leon et al., 2006). Nucleoside hydrolase DNA vaccine (Leishmune®’s) has been found effective for immunotherapy of canine visceral leishmaniasis (Borja-Cabrera et al., 2012). DNA vaccine for Anaplasma mariginale targeting the major surface protein namely MSP1b yielded good antibody response and fair protection was noticed in challenge studies in cattle (De Andrade et al., 2004). Development of DNA vaccine targeting babesiosis disease in canines has been attempted with protein p50 gene coding the Babesia canis, which generated protective immunity, while the p36/LACK antigen gene and L. major GP63 antigen encoding gene inducing Th1 immune response has been utilized for Leishmaniosis (Walker et al., 1998; Fukumoto et al., 2007). DNA vaccines have also been developed against coccidiosis in birds, employing the 3-1E and EtMIC2 genes (Min et al., 2001; Ding et al., 2005).
Advantages:
| • | Safe as do not contain any form of pathogen |
| • | No risk of infection |
| • | Administration is easy by either I/M or I/D routes |
| • | Manufacture of multivalent vaccines by combining multiple DNA |
| • | Stability of the vaccine for storage and shipping especially in tropical zones |
| • | As compared to live attenuated or recombinant vaccines DNA vaccines are more stable |
| • | Cost effective, hence ideally affordable |
| • | Produces both humoral and cellular immune response |
| • | Unlike live attenuated vaccine there is no risk of reversion to virulence |
Disadvantages:
| • | Risk of effective genes controlling cell growth |
| • | Possibility of including antibody production against DNA |
| • | Chance of inducing mutation |
| • | Complexity in vaccine formulation |
| • | Plasmid may get integrated into the cell and can lead to transfer of resistance gene (Kumaragurubaran and Kaliaperumal, 2013) |
Edible vaccine: Recent advances in sciences has made it possible that vaccine can be administered orally as edible vaccines hence eliminating the problem of pain which is associated with the needle pricks during vaccination (Dhama et al., 2013e). Though the concept of producing edible vaccine started in 1990’s but it had rapid strides and all sorts of plants are being exploited in the recent years so as to get good expression of the required gene of interest so that good immunity may be elicited against a particular organism (Curtiss and Cardineau, 1990). Edible vaccine which enters orally released in the intestinal tract elicits both humoral and mucosal immunity. More work has been carried out the malarian parasite which of importance in human aspect (Clemente and Corigliano, 2012). Tomato has been used as an expression plant so that it can be easily consumed by human (Chowdhury and Bagasra, 2007). In livestock sector edible vaccines has been developed against coccidiosis in poultry which is an important disease in poultry industry, caused by Eimeria tenella. Microneme protein EtMIC2 of this deadly pathogen was expressed in tobacco leaves (Sathish et al., 2011). Feeding trails of this tobacco leaves with the microneme protein resulted in good antibody synthesis and there was low oocyst shed down in the droppings. The combination of EtMIC2 and EtMIC1 yielded good antibody response along with increase in weight gain (Sathish et al., 2012).
Advantages:
| • | Plant cell wall act as a barrier protecting the antigen of interest to reach the target place safely (Streatfield, 2006; Hayden et al., 2012) |
| • | Plants contain some phytochemicals which act with co-ordination with the antigen so that it provides good immune response even in the absence of adjuvants (Pasquevich et al., 2011) |
| • | Secondary plant metabolites like lectins, saponins, alkaloids, phenolic compounds and flavonoids act as an immune stimulant (Kostrzak et al., 2009) |
| • | Edible vaccines are free from bacteria, fungi and viruses because plant pathogens are not deleterious to animals |
| • | Needle prick is eliminated and hence there is no pain and no damage to muscles |
| • | As the administration of oral vaccine is much easier than injection, it can be applied for mass vaccine and time can be saved |
| • | Production can be done in large scale with available plants locally and can be stored for longer period (Dhama et al., 2013e) |
Disadvantages:
| • | Negative impact on public because of the use of edible plants for production of vaccine |
| • | Possibility of inducing oral tolerance and side effects like allergy |
| • | To produce highly expressive plants it needs time and patience (Jacob et al., 2013) |
VACCINES AVAILABLE AGAINST SOME OF THE PROTOZOAN PARASITES
Vaccines against coccidia
Coccivac®-B: It is a live, sporulated oocyst vaccine produced from isolates that were collected in the late 1940's, before the current anticoccidial products were introduced. This vaccine is prepared from anticoccidial-sensitive strains of Eimeria acervulina, E. mivati, E. maxima and E. tenella, unlike present-day field oocysts, these isolates had never been subjected to selection pressure by anticoccidials resulting in resistance. Coccivac-B vaccine is a valuable tool to restore the performance of existing anticoccidials (Chapman, 1994). In experimental study on birds, this vaccine produced variable degrees of protection with the five different strains of E. tenella (Awad et al., 2013).
Coccivac®-D: It is a live, sporulated oocyst vaccine containing different species of Eimeria viz., Eimeria acervulina, E. maxima, E. necatrix, E. hagani, E. praecox, E. tenella, E. brunetti and E. mivati. To induce complete immunity, the original dose of coccidial oocysts must complete at least four life cycles in the flock (Johnson and Reid, 1970).
Coccivac®-T: It is a live coccidiosis vaccine containing sporulated oocysts of Eimeria dispersa, E. meleagrimitis, E. adenoides and E. gallopavonis that is administered to day old turkey poults via spray cabinet. The strains of Eimeria in Coccivac-T were isolated prior to common use of modern ionophore and chemical anticoccidial products and are highly sensitive to all in-feed anticoccidials (Mathis and McDougald, 1989).
Nobilis® COXATM: This vaccine is a live vaccine having a special property of being active even in presence of ionophore compounds. The vaccine contains strains of E. acervulina, E. tenella and E. maxima which are tolerant to ionophore compound. The benefit of this vaccine is that apart from production of immunity the vaccine allows the use of ionophore compounds during the 3-4 weeks of age when the immunity is poorly developed (Vermeulen et al., 2001).
Eimeriavax -4 m: This vaccine contains viable oocysts of Eimeria acervulina Strain RA, E. maxima Strain MCK+10, E. necatrix Strain mednec3+8 and E. tenella Strain Rt3+15 suspended in phosphate buffered saline (PBS). Each dose comprises a minimum of E. acervulina 50 oocysts, E. maxima 100 oocysts, E. necatrix 100 oocysts and E. tenella 150 oocysts, with a minimum predicted titre of 1.6x104 oocysts mL-1 at the end of the shelf-life (Gore et al., 1983). The advantages of Eimeria vax 4 m are:
| • | Simple, single dose, eye-drop or oral application |
| • | Safe in chickens from day-old |
| • | Immunity induced within 10 days against the four major species of Eimeria (E. acervulina, E. maxima, E. necatrix and E. tenella) |
| • | Effective in broiler breeders, broilers, free range and layer flocks |
| • | Supplied ready to use |
| • | Anticoccidial medications are not required |
| • | Provides productivity improvements (McDonald and Shirley, 2009) |
Immucox: Oral coccidiosis vaccine of live oocysts of Eimeria spp. designed to help healthy Chicken Broilers and Roasters to develop immunity to coccidiosis. This is a one-time vaccination that delivers protective immunity through the productive life of the bird. It is a vaccine approved for water or gel delivery in the hatchery (Lee, 2006).
Paracox®-8: It is a live attenuated oral vaccine consisting of translucent, suspension of oocysts derived from eight precocious lines of coccidian, used for the active immunization of chickens against different Eimeria spp. viz., Eimeria acervulina, E. tenella, E. brunetti, E. mitis, E. necatrix, E. praecox and E. maxima (Shirley et al., 2005). The dose of vaccine is 0.1 mL per chicken, which can be administered orally either by spray on feed or in drinking water or by hatchery spray.
Livacox: It is live attenuated coccidiosis vaccines for domestic poultry (Gallus domesticus). Its range consists of LIVACOX® T for broilers and LIVACOX® Q for breeder and layer pullets (Shirley and Bedrnik, 1997).
CoxAbic: It is first commercially available subunit vaccine for poultry and contains purified antigens isolated from the macrogametocyte (sexual stage) of Eimeria maxima (Belli et al., 2004). Vaccination using gametocyte antigen through the breast muscle will guide to production of antibody response (Sharman et al., 2010).
Vaccines for anaplasma
Anaplaz: It is the first anaplasmosis vaccine manufactured for cattle in the United States by Fort Dodge. More recently, Mallinkrodt (later Schering-Plough) marketed a vaccine called Plazvax®. Both of these vaccines protect against Anaplasmosis by similar mechanisms. The vaccines contain killed Anaplasma marginale, harvested from infected cattle (Brock et al., 1964). The vaccines do not prevent the animal against infection by the Anaplasma organism, but protect the animal from development of clinical disease. They are "immune carriers”. That is to say, they are "immune" to becoming sick from the agent but are carriers of the agent. Dose rate of 1 mL by sec cycle-1 route and repeated after 3-4 week and revaccinate annually by single dose, 1 mL.
Anavac: It is a modified live vaccine, which is safe and effective when given to young cattle. They become infected with the vaccine strain of Anaplasma and are "immune carriers”. Dose rate of 2 mL i/v and is given at 6-12 month of age. Booster doses are recommended every 1-2 years depending on herd history (Ristic, 1960).
Vaccines against giardia
GiardiaVax: It is a killed culture trophozoite vaccine for dogs and prevent the disease and shedding of Giardia lamblia. The vaccine is derived from G. duodenalis isolated from sheep. Dose of vaccine is 1ml by subcutaneous route. First dose at 8 weeks of age and second after 2-4 weeks and then repeated annually. Dogs which had failed to be cured of giardiasis following chemotherapeutic measures were treated with a Giardia vaccine (2-3 injections). After immunization, the clinical signs diminish within 16-42 days, therefore it is a good method for treating giardiasis in dogs.
Vaccines against babesia: It is live attenuated vaccine, developed by in-vitro culture (Levy and Ristic, 1980). Cattle were vaccinated with exo-antigen from culture (MASP) elicit humoral and cell mediated immunity (Alexis et al., 1993).
Pirodog/Nobivac® Piro: It is a Soluble Parasite Antigen (SPA) of supernatants of in vitro cultures (B. canis and B. rossi) gives 80% protection and immunity last for about 6 months (Schetters, 2005). It is given at 6 months of age and booster vaccination is required 3 to 6 weeks after the initial vaccination and thereafter revaccination every 6 months by intramuscular route. Both the vaccine produces some local reaction at the site of injection but this reaction is more in case of Nobivac Piro vaccine (Freyburger et al., 2011).
Vaccine against neospora: An ideal vaccine have to provide protection against both infection and the clinical signs, so there is requirement of vaccine that can induce a non-foetopathic cell mediated immune response (Goodswen et al., 2013). Recently, a Neospora caninum killed tachyzoite based vaccine has been reported to be efficacious in preventing abortion in dairy cattle (Williams et al., 2007; Weston et al., 2012). Proteins, which have important role in adhesion/invasion or other parasite-host-cell interaction processes can provide protection against Neospora infection and can be targets for the development of an effective vaccine against this important protozoan parasite (Hemphill et al., 2013).
Bovilis® Neoguard: It is prepared by killed tachyzoite of Neospora caninum with spur adjuvant which reduces abortion in cattle by more than 50% (Meeusen et al., 2007; Weston et al., 2012) but may increase the early embryonic death, if used in pregnancy (Weston et al., 2012). The vaccination induce antibody at a high level which give protection upto 1 year, so booster vaccination after 1 year is required. It is administered in two doses of 5 ml at one month apart, the first dose given between day 75 and 90 of gestation, booster in 3-4 weeks with 2 annual boosters 3-4 weeks apart by subcutaneous route.
Live tachyzoite vaccine: Tachyzoites were maintained under conditions in a continuous passage of vero cell-lines at 37°C and 5% CO2 in air and in RPMI 1640 medium supplemented with 2% horse serum and penicillin-streptomycin (100 IU mL-1 100 g mL-1). This vaccine protects the losses due to death of foetus (Williams et al., 2007; Hecker et al., 2012; Goodswen et al., 2013).
Vaccines against toxoplasma: Effective vaccines against Toxoplasma gondii, a protozoan parasite infecting both animals and humans, are very helpful in preventing and controlling toxoplasmosis (Innes et al., 2009; Liu et al., 2012). Invasion factors of T. gondii viz., microneme protein 6 and 8 (MIC6, MIC 8) have also been proposed to be a useful vaccine contender for toxoplasmosis (Peng et al., 2009; Liu et al., 2010). Some experimental trials have been going on the protective efficacy of recombinant T. gondii PDI (rTgPDI) as a vaccine candidate for combating toxoplasmosis (Wang et al., 2013).
S48 strain (Toxovax): It is a live vaccine containing originally isolated tachyzoite from aborted placenta and maintain in laboratory by repeated passage in mice. It was initially developed for sheep but in cats it inhibits sexual development of T. gondii (Verma and Khanna, 2012). Loss ability to form bradyzoites or oocysts and eliminated within 14 days by host immune response (INF-gamma). The S48 strain when ingested by cat after voided by cat does not cause production of oocyst. It prevent from spreading in the body; placenta as well as meat contamination for 18 months. It is given 4 weeks before mating by intramuscular injection on the neck (O’Connell et al., 1988). It should be given @ 2 mL intramuscularly. Basic vaccination should be given as single dose at least 3 weeks prior to mating. Re-vaccination after 2 years with a single dose atleast 3 weeks prior to mating is recommended.
T263: It is a bradizoite of live mutant T. gondii that does not formed an oocyst. The administration of T263 yield leads to reduction/prevention of oocyst shedding in cats (Verma and Khanna, 2012).
Vaccine against Theileria: The process of development of Theileria annulata cultures involves cultivation of the organism for a prolonged time (Pipano and Tsur, 1966). The organisms are collected from the infected animal either by Lymph node biopsy or whole blood. The in vitro culture yields agamogenic strain of the organism. This cell culture attenuated vaccine (Rakshavac-T) has an efficacy approaching 95-100% (Brown et al., 2006). The site for inoculation is mid-neck region@ 3 mL. Calves of 2 months age and above only should be vaccinated. This vaccine should not be used in advanced stage of pregnancy. Attenuated schizont vaccine are effective in prevention of theileriosis in cattle (Barriga, 1994; Pipano and Shkap, 2000) however more safer and effective vaccines need to be developed utilizing advances in molecular tools and techniques for control of theileriosis in bovines and ovines (Yin et al., 2008). Vaccines against Bovine theleriosis caused by Theileria parva should be mixture of several antigens derived from both sporozoite and schizont stages, leading to strong immunity (Morzaria et al., 2000).
Vaccine against Leishmania: Different types of vaccines for prevention of leishmaniasis have been developed (Brodskyn et al., 2003; Coler et al., 2007; Nagill and Kaur, 2011).
Leishmanization: It is inoculation of exudates from the active lesion of an infected person to non-infected susceptible person to give an immunization. It has been use from older times but the use is now limited.
Killed vaccine: The killed promastigote are use for immunization with or without BCG. Leishmania braziliensis promastigote killed by autoclaving give good results.
Leishmune: It blocks the transmission of canine visceral leishmaniasis (Nogueira et al., 2005). It has gp63 protein of L. donovani that is adjuvanted in saponin (Parra et al., 2007). This vaccine has 76-80% efficacy. It is transmission blocking because:
| • | The number of Leishmania protozoa in the skin of a dog is reduced in number. Thus the Phlebotomine flies does not readily get the the protozoa during blood feeding |
| • | The antibody produced does not allow the development of an infective promastigote inside the fly vector |
Leish111f: One of the most promising finding is the recombinant protein called Leish 111f along with an adjuvant MPL-SE (Monophosphoryl Lipid-stable emulsion) shown by Coler et al. (2007). It is a recombinant protein of:
| LeIF: | L. braziliensis initiation and elongation factor |
| TSA: | Thiol-specific antioxidant |
| LmSTI1: | L. major stress inducible protein |
The Leish 111f is having an added advantage like the vaccine efficacy is very high reaching upto 99.6%. Another is that it gives cross protection i.e., Leishmania donovani, which can be also used as a therapy especially trial in man. In mucocuteneos Leishmaniosis caused by L. briziliensis and other cuteneous Leishmaniosis. But the treatment schedule takes a very long time.
Vaccines for Sarcocystis:
EPM vaccine: It consists of in vitro cultured merozoites, obtained from the spinal cord of horse, which are chemically inactivated and mixed with suitable adjuvants (Marsh et al., 2004). It gives protection against a neurological disease in horses caused by infection with Sarcocystis neurona, equine protozoal myeloencephalitis. It is given@ 1 mL intramuscularly and booster vaccination 3 to 6 weeks after the first dose, then revaccination annually.
S. neurona SAG1 protein vaccine: It is subunit vaccine prepared from major Surface Antigen Gene 1 (SAG1), which is conserved among members of Sarcocystidae (Elsheikha and Mansfield, 2004). Horses were vaccinated on days 0 and 21 with 1ml adjuvanted rSnSAG1 (50 μg) or 1 mL adjuvant alone by intramuscular injection in the left side of the neck (Ellison and Witonsky, 2009). Booster vaccination is required after 3 to 6 weeks of the first dose and then revaccination annually.
Vaccine against Trypanosoma:
Beta-tubulin: The beta-tubulin gene of Trypanosoma evansi (STIB 806) was cloned and expressed in Escherichia coli (Li et al., 2007). Beta-tubulin is important for cellular structure and physical functions. Recombinant beta-tubulin was expressed as inclusion bodies in E. coli.
TSA (T. brucei DNA vaccine encoding TSA Protein): The DNA vaccination process was able to protect 60% of mice submitted to a challenge assay with the infective form of T. brucei brucei parasites (Silva et al., 2009).
MAPp15 (Microtubule associated protein): In an experimental study, vaccination of mice with p15 (native or recombinant) provides complete protection from Trypanosoma brucei suggesting it as an effective vaccine (Rasooly and Balaban, 2004).
Vaccine against Tritrichomonas Foetus
Trichguard: It is killed protozoan vaccine for cattle for prevention of infection by Tritrichomonas foetus. It is given to cattle @ 1-2 mL subcutaneously, booster vaccination 2-4 weeks after first dose and then revaccination annually. The last injection should proceed the breeding season by 4 weeks.
COBWEB FOR VACCINE DEVELOPMENT AGAINST PROTOZOAN INFECTION
There are lot many factors which causes the pitfall for development of a successful protozoan vaccine which is of the supreme importance to keep infection at the gate way. The important factors are pathogen associated, vaccine market associated or funding (Vercruysse et al., 2007). Among all the factors pathogen associated problems needs to be dealt in a more scientific and clever way as some protozoa are clever enough to escape the immune system. One of the most important protozoa of this type is the Trypanasoma spp. which causes a serious problem in case of animals as well as humans (Vanhamme et al., 2001; La Greca and Magez, 2011). One of the noteworthy features of this protozoa is that it can change its antigenic surface protein in such a rapid manner so that the immune system of host cannot trace it (Pays et al., 2004). Many protozoa also have a complex life cycle during their survival in the host. Two stages of life cycle namely sexual and asexual exist in the life pattern of many protozoa and hence selecting a vaccine candidate becomes a difficult task. Certain situation occurs when a single host cell have protozoa at different stages of development giving a feel to the host immune system that there a number of protozoa in a single cell (Vercruysse et al., 2007).
CONCLUSION
Drugs remain central to alleviating clinical disease and for larger scale disease control programmes. However, available drugs have often been in use for decades and drug resistance in the target parasites is now prevalent and, particularly in the case of livestock, threatening sustainable control. Control is threatened by the widespread appearance of drug-resistant parasites in animals. Development of drug resistance could limit the use of anti-parasitic drugs which will remain important for a long time. Drug residues found in the food animal, development of drug resistance and lack of development of new drug are all major problems or reasons which diverts the scientist to make efforts on vaccine development related research. Many vaccines are included in the control of parasitic diseases programs, as these approaches may lead to a substantial reduction in the use of chemical drugs for prevention and control of parasitic diseases. The need of hour is to apply the immunization results of murine/lab animal model in veterinary science and for humankind. Vaccination may also aim to improve public health, so there is a need of cheap and effective vaccine to be developed and reach with success in the market. Development of vaccine against protozoan parasite is cumbersome process because of their complex life cycle, many developmental stages and sub clinical form of disease. Efforts for making economically feasible and effective protozoan vaccine, strategies to discover new antigen to act as novel vaccine candidates are still in continuation.
REFERENCES
- Aguilar-Be, I., R. da Silva Zardo, E. Paraguai de Souza, G.P. Borja-Cabrera and M. Rosado-Vallado et al., 2005. Cross-protective efficacy of a prophylactic Leishmania donovani DNA vaccine against visceral and cutaneous murine leishmaniasis. Infect. Immunity., 73: 812-819.
Direct Link - Alexis, V., E. Precigout, M. L'hostis, B. Carcy, A. Gorenflot and J. Schrevel, 1993. Cellular and humoral immune responses induced in cattle by vaccination with Babesia divergens culture-derived exoantigens correlate with protection. Infect. Immun., 61: 734-741.
Direct Link - Awad, A.M., A.F. El-Nahas and S.S. Abu-Akkada, 2013. Evaluation of the protective efficacy of the anticoccidial vaccine Coccivac-B in broilers, when challenged with Egyptian field isolates of E. tenella. Parasitol. Res., 112: 113-121.
CrossRefDirect Link - Baltzell, P., H. Newton and A.M. O'Connor, 2013. A critical review and meta-analysis of the efficacy of whole-cell killed Tritrichomonas foetus vaccines in beef cattle. J. Vet. Inter. Med., 27: 760-770.
CrossRef - Barriga, O.O., 1994. A review on vaccination against protozoa and arthropods of veterinary importance. Vet. Parasitol., 55: 29-55.
CrossRef - Borja-Cabrera, G.P., F.B. Santos, D. Nico, A.E. Gravino, L. Manna, M. Palatnik, C.B. Palatnik-de-Sousa, 2012. The Leishmune®’s nucleoside hydrolase DNA vaccine as an aid in immunotherapy of canine visceral leishmaniasis. Proc. Vaccinol., 6: 64-73.
CrossRef - Brodskyn, C., C.I. de Oliveira, A. Barral and M. Barral-Netto, 2003. Vaccines in leishmaniasis: Advances in the last five years. Expert Rev. Vaccines, 2: 705-717.
PubMed - Brown, W.C., J. Norimine, W.L. Goff, C.E. Suarez and T.F. Mcelwain, 2006. Prospects for recombinant vaccines against Babesia bovis and related parasites. Parasite Immunol., 28: 315-327.
CrossRefDirect Link - Carvalho, J.A., J. Rodgers, J. Atouguia, D.M.F. Prazeres and G.A. Monteiro, 2010. DNA vaccines: A rational design against parasitic diseases. Exp. Rev. Vaccines, 9: 175-191.
CrossRef - Cascio, A., M. Bosilkovski, A.J. Rodriguez-Morales and G. Pappas, 2011. The socio-ecology of zoonotic infections. Clin. Microbiol. Infect., 17: 336-342.
CrossRef - Chen, J., S.Y. Huang, Z.Y. Li, Z.G. Yuan and D.H. Zhou et al., 2013. Protective immunity induced by a DNA vaccine expressing eIF4A of Toxoplasma gondii against acute toxoplasmosis in mice. Vaccine, 31: 1734-1739.
CrossRef - Chowdhury, K. and O. Bagasra, 2007. An edible vaccine for malaria using transgenic tomatoes of varying sizes, shapes and colors to carry different antigens. Med. Hypotheses, 68: 22-30.
Direct Link - Clemente, M. and M.G. Corigliano, 2012. Overview of plant-made vaccine antigens against malaria. J. Bio. Biotechnol., Vol. 2012.
CrossRef - Coler, R.N., Y. Goto, L. Bogatzki, V. Raman and S. Reed, 2007. Leish-111f, a recombinant polyprotein vaccine that protects against visceral Leishmaniasis by elicitation of CD4+ T cells. Infect. Immun., 75: 4648-4654.
Direct Link - Collett, M.G., 2000. Survey of canine babesiosis in South Africa. J. S. Afr. Vet. Assoc., 71: 180-186.
Direct Link - Cornelissen, A.W.C.A. and T.P.M. Schetters, 1996. Vaccines against protozoal diseases of veterinary importance. FEMS Immunol. Med. Microbiol., 15: 61-72.
CrossRef - Da’dara, A.A. and D.A. Harn, 2005. DNA vaccines against tropical parasitic diseases. Exp. Rev. Vaccines, 4: 575-589.
CrossRef - Dantas-Torres, F., 2006. Leishmune vaccine: the newest tool for prevention and control of canine visceral leishmaniosis and its potential as a transmission-blocking vaccine. Vet. Parasitol., 141: 1-8.
Direct Link - De Andrade, G.M., R.Z. Machado, M.C. Vidotto and O. Vidotto, 2004. Immunization of bovines using a DNA vaccine (pcDNA3.1/MSP1b) prepared from the jaboticabal strain of Anaplasma marginale. Ann. N. Y. Acad. Sci., 1026: 257-266.
CrossRef - De Ruyck, H., R. Van Renterghem, H. De Ridder and D. De Brabander, 2000. Determination of anthelmintic residues in milk by high performance liquid chromatography. Food Control, 11: 165-173.
CrossRef - Dhama, K., M. Mahendran, P.K. Gupta and A. Rai, 2008. DNA vaccines and their applications in veterinary practice: Current perspectives. Vet. Res. Commun., 32: 341-356.
CrossRefDirect Link - Dhama, K., S. Chakraborty, S. Kapoor, R. Tiwari and A. Kumar et al., 2013. One world, one health-veterinary perspectives. Adv. Anim. Vet. Sci., 1: 5-13.
Direct Link - Dhama, K., S. Chakraborty, Mahima, M.Y. Wani and A.K. Verma et al., 2013. Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Biol. Sci., 16: 101-111.
CrossRefDirect Link - Dhama, K., S. Rajagunalan, S. Chakraborty, A.K. Verma, A. Kumar, R. Tiwari and S. Kapoor, 2013. Food-borne pathogens of animal origin-diagnosis, prevention, control and their zoonotic significance: A review. Pak. J. Biol. Sci., 16: 1076-1085.
CrossRefDirect Link - Dhama, K., R. Tiwari, S. Chakraborty, A. Kumar, M. Karikalan, R. Singh and R.B. Rai, 2013. Global warming and emerging infectious diseases of animals and humans: Current scenario, challenges, solutions and future perspectives: A review. Int. J. Curr. Res., 5: 1942-1958.
Direct Link - Dhama, K., A.K. Verma, R. Tiwari, S. Chakraborty and K. Vora et al., 2013. A perspective on applications of Geographical Information System (GIS): An advanced tracking tool for disease surveillance and monitoring in veterinary epidemiology. Adv. Anim. Vet. Sci., 1: 14-24.
Direct Link - Ding, X., H.S. Lillehoj, R.A. Dalloul, W. Min, T. Sato, A. Yasuda and E.P. Lillehoj, 2005. In ovo vaccination with the Eimeria tenella EtMIC2 gene induces protective immunity against coccidiosis. Vaccine, 23: 3733-3740.
CrossRef - Dondji, B., E. Perez-Jimenez, K. Goldsmith-Pestana, M. Esteban and D. McMahon-Pratt, 2005. Heterologous prime-boost vaccination with the LACK antigen protects against murine visceral leishmaniasis. Infect. Immun., 73: 5286-5289.
Direct Link - Dumonteil, E., 2007. DNA vaccines against protozoan parasites: Advances and challenges. J. Bio. Biotechnol., Vol. 2007.
CrossRef - Ellison, S. and S. Witonsky, 2009. Evidence that antibodies against recombinant SnSAG1 of Sarcocystis neurona merozoites are involved in infection and immunity in equine protozoal myeloencephalitis. Can. J. Vet. Res., 73: 176-183.
Direct Link - Meeusen, E.N.T., J. Walker, A. Peters, P.P. Pastoret and G. Jungersen, 2007. Current status of veterinary vaccines. Clin. Microbiol. Rev., 20: 489-510.
CrossRefDirect Link - Elsheikha, H.M. and L.S. Mansfield, 2004. Sarcocystis neurona major Surface Antigen Gene 1 (SAG1) shows evidence of having evolved under positive selection pressure. Parasitol Res., 94: 452-459.
Direct Link - Fitzpatrick, J.L., 2013. Global food security: The impact of veterinary parasites and parasitologists. Vet. Parasitol., 195: 233-248.
CrossRefDirect Link - Freyburger, L., L. Lemaitre, C. Medaille, F. Oberli, L. Fanchon and P. Bergamo, 2011. Comparative safety study of two commercialised vaccines against canine babesiosis induced by Babesia canis. Parasite, 18: 311-318.
Direct Link - Fukumoto, S., Y. Tamaki, M. Okamura, H. Bannai and N. Yokoyama et al., 2007. Prime-boost immunization with DNA followed by a recombinant vaccinia virus expressing P50 induced protective immunity against Babesia gibsoni infection in dogs. Vaccine, 25: 1334-1341.
CrossRef - Gamboa-Leon, R., E.P. de Souza, G.P. Borja-Cabrera, F.N. Santos and L.M. Myashiro et al., 2006. Immunotherapy against visceral leishmaniasis with the nucleoside hydrolase-DNA vaccine of Leishmania donovani. Vaccine, 24: 4863-4873.
CrossRef - Geerts, S., P.H. Holmes, M.C. Eisler and O. Diall, 2001. African bovine trypanosomiasis: The problem of drug resistance. Trends Parasitol., 17: 25-28.
PubMed - Goodswen, S.J., P.J. Kennedy and J.T. Ellis, 2013. A review of the infection, genetics and evolution of Neospora caninum: From the past to the present. Infection Genet. Evol., 13: 133-150.
CrossRef - Gore, T.C., P.L. Long, M. Kogut and J. Johnson, 1983. Attenuation of MEimeria necatrix and E. tenella of U. S. origin by serial embryo passage. Avian Dis., 27: 569-576.
Direct Link - La Greca, F. and S. Magez, 2011. Vaccination against trypanosomiasis: Can it be done or is the trypanosome truly the ultimate immune destroyer and escape artist? Hum Vaccin, 7: 1225-1233.
CrossRef - Handman, E., A.H. Noormohammadi, J.M. Curtis, T. Baldwin and A. Sjolander, 2000. Therapy of murine cutaneous leishmaniasis by DNA vaccination. Vaccine, 18: 3011-3017.
CrossRef - Hayden, C.A., S.J. Streatfield, B.J. Lamphear, G.M. Fake and T.K. Keener et al., 2012. Bioencapsulation of the hepatitis B surface antigen and its use as an effective oral immunogen. Vaccine, 30: 2937-2942.
CrossRefDirect Link - Hecker, Y.P., M.C. Venturini, C.M. Campero, A.C. Odeon and D.P. Moore, 2012. Advances in the development of vaccines for bovine neosporosis. Rev. Argent. Microbiol., 44: 216-230.
Direct Link - Hemphill, A., K. Debache, T. Monney, M. Schorer and C. Guionaud et al., 2013. Proteins mediating the Neospora caninum-host cell interaction as targets for vaccination. Front Biosci. (Elite Ed), 5: 23-36.
Direct Link - Ingolotti, M., O. Kawalekar, D.J. Shedlock, K. Muthumani and D.B. Weiner, 2010. DNA vaccines for targeting bacterial infections. Exp. Rev. Vaccines, 9: 747-763.
CrossRefDirect Link - Innes, E.A. and A.N. Vermeulen, 2006. Vaccination as a control strategy against the coccidial parasites Eimeria, Toxoplasma and Neospora. Parasitology, 133: S145-S168.
CrossRefDirect Link - Innes, E.A., P.M. Bartley, S. Maley, F. Katzer and D. Buxton, 2009. Veterinary vaccines against Toxoplasma gondii. Mem. Inst. Oswaldo Cruz., 104: 246-251.
Direct Link - Innes, E.A., P.M. Bartley, M. Rocchi, J.B. Silvan and A. Burrells et al., 2011. Developing vaccines to control protozoan parasites in ruminants: Dead or alive? Vet. Parasitol., 180: 155-163.
CrossRef - Jacob, S.S., S. Cherian, T.G. Sumithra, O.K. Raina and M. Sankar, 2013. Edible vaccines against veterinary parasitic diseases-Current status and future prospects. Vaccine, 31: 1879-1885.
CrossRef - Jang, S.I., H.S. Lillehoj, S.H. Lee, K.W. Lee and M.S. Park et al., 2010. Eimeria maxima recombinant Gam82 gametocyte antigen vaccine protects against coccidiosis and augments humoral and cell-mediated immunity. Vaccine, 28: 2980-2985.
CrossRef - Johnson, J. and W.M. Reid, 1970. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol., 28: 30-36.
CrossRefPubMedDirect Link - Jones, K.E., N.G. Patel, M.A. Levy, A. Storeygard, D. Balk, J.L. Gittleman and P. Daszak, 2008. Global trends in emerging infectious diseases. Nature, 451: 990-993.
CrossRefPubMedDirect Link - Knox, D.P., 2010. Parasite vaccines: Recent progress in and problems associated with their development. Open Infect. Dis. J., 4: 63-75.
Direct Link - Kostrzak, A., M.C. Gonzalez, D. Guetard, D. Guetard and D.B. Nagaraju et al., 2009. Oral administration of low doses of plant-based HBsAg induced antigen-specific IgAs and IgGs in mice, without increasing levels of regulatory T cells. Vaccine, 27: 4798-4807.
CrossRef - Kumar, A., A.K. Verma and A. Rahal, 2011. Mycoplasma bovis, a multi disease producing pathogen: An overview. Asian J. Anim. Vet. Adv., 6: 537-546.
CrossRef - Kumaragurubaran, K. and K. Kaliaperumal, 2013. DNA Vaccine: The miniature miracle. Vet. World, 6: 228-232.
CrossRef - Lee, E.H., 2006. Combination of vaccination and medication in the same crop for the control of coccidiosis in chickens and Turkeys. Int. Poult. Prod., 14: 23-23.
Direct Link - Levy, M.G. and M. Ristic, 1980. Babesia bovis: Continuous cultivation in a microaerophilous stationary phase culture. Science, 207: 1218-1220.
PubMed - Li, S.Q., M.C. Fung, S.A. Reid, N. Inoue and Z.R. Lun, 2007. Immunization with recombinant β-tubulin from Trypanosoma evansi induced protection against T. evansi, T. equiperdum and T. b. brucei infection in mice. Parasite Immunol., 29: 191-199.
CrossRefPubMedDirect Link - Lightowlers, M.W., 1994. Vaccination against animal parasites. Vet. Parasitol., 54: 177-204.
CrossRefPubMedDirect Link - Liu, M.A., B. Wahren and G.B.K. Hedestam, 2006. DNA vaccines: Recent developments and future possibilities. Hum. Gene Ther., 17: 1051-1061.
Direct Link - Liu, M.M., Z.G. Yuan, G.H. Peng, D.H. Zhou and X.H. He et al., 2010. Toxoplasma gondii microneme protein 8 (MIC8) is a potential vaccine candidate against toxoplasmosis. Parasitol. Res., 106: 1079-1084.
PubMed - Liu, Q., L.D. Singla and H. Zhou, 2012. Vaccines against Toxoplasma gondii: Status, challenges and future directions. Hum. Vaccines Immunotherapeut., 8: 1305-1308.
CrossRefPubMedDirect Link - Mahima, A. Rahal, R. Deb, S.K. Latheef and H.A. Samad et al., 2012. Immunomodulatory and therapeutic potentials of herbal, traditional/indigenous and ethnoveterinary medicines. Pak. J. Biol. Sci., 15: 754-774.
CrossRefDirect Link - Marsh, A.E., J. Lakritz, P.J. Johnson, M.A. Miller and H.J. Chu, 2004. Evaluation of immune responses in horses immunized using a killed Sarcocystis neurona vaccine. Vet. Ther., 5: 34-42.
PubMed - McDonald, V. and M.W. Shirley, 2009. Past and future: Vaccination against Eimeria. Parasitology, 136: 1477-1489.
CrossRefPubMedDirect Link - Meng, M., S. He, G. Zhao, Y. Bai, H. Zhou, H. Cong, G. Lu, Q. Zhao and X.Q. Zhu, 2012. Evaluation of protective immune responses induced by DNA vaccines encoding Toxoplasma gondii surface antigen 1 (SAG1) and 14-3-3 protein in BALB/c mice. Parasit. Vectors, 5: 273-273.
PubMed - Min, W., H.S. Lillehoj, J. Burnside, K.C. Weining, P. Staeheli and J.J. Zhu, 2001. Adjuvant effects of IL-1β, IL-2, IL-8, IL-15, IFN-α, IFN-γTGF-β4 and lymphotactin on DNA vaccination against Eimeria acervulina. Vaccine, 20: 267-274.
CrossRefDirect Link - Morzaria, S., V. Nene, R. Bishop and A. Musoke, 2000. Vaccines against Theileria parva. Ann. N. Y. Acad. Sci., 916: 464-473.
CrossRefPubMedDirect Link - Nagill, R. and S. Kaur, 2011. Vaccine candidates for leishmaniasis: A review. Int. Immunopharmacol., 11: 1464-1488.
CrossRefPubMedDirect Link - Nielsen, H.V., M. Di Cristina, E. Beghetto, A. Spadoni, E. Petersen and N. Gargano, 2006. Toxoplasma gondii: DNA vaccination with bradyzoite antigens induces protective immunity in mice against oral infection with parasite cysts. Exp. Parasitol., 112: 274-279.
PubMed - Nogueira, F.S., M.A. Moreira, G.P. Borja-Cabrera, F.N. Santos and I. Menz et al., 2005. Leishmune vaccine blocks the transmission of canine visceral leishmaniasis: Absence of Leishmania parasites in blood, skin and lymph nodes of vaccinated exposed dogs. Vaccine, 23: 4805-4810.
CrossRefPubMedDirect Link - O'Connell, E., M.F. Wilkins and W.A. te Punga, 1988. Toxoplasmosis in sheep. II. The ability of a live vaccine to prevent lamb losses after an intravenous challenge with Toxoplasma gondii. N. Z. Vet. J., 36: 1-4.
PubMed - Parra, L.E., G.P. Borja-Cabrera, F.N. Santos, L.O. Souza, C.B. Palatnik-de-Sousa and I. Menz, 2007. Safety trial using the Leishmune vaccine against canine visceral leishmaniasis in Brazil. Vaccine, 25: 2180-2186.
PubMed - Pasquevich, K.A., A.E. Ibanez, L.M. Coria, C. Garcia Samartino and S.M. Estein et al., 2011. An oral vaccine based on U-Omp19 induces protection against B. abortus mucosal challenge by inducing an adaptive IL-17 immune response in mice. PLoS ONE, Vol. 6.
CrossRefDirect Link - Paul-Pierre, P., 2009. Emerging diseases, zoonoses and vaccines to control them. Vaccine, 27: 6435-6438.
CrossRef - Pays, E., L. Vanhamme and D. Perez-Morga, 2004. Antigenic variation in Trypanosoma brucei: Facts, challenges and mysteries. Curr. Opin. Microbiol., 7: 369-374.
CrossRef - Peng, G.H., Z.G. Yuan, D.H. Zhou, X.H. He and M.M. Liu et al., 2009. Toxoplasma gondii microneme protein 6 (MIC6) is a potential vaccine candidate against toxoplasmosis in mice. Vaccine, 27: 6570-6574.
PubMedDirect Link - Pipano, E. and V. Shkap, 2000. Vaccination against tropical theileriosis. Ann. N. Y. Acad. Sci., 916: 484-500.
PubMed - Rasooly, R. and N. Balaban, 2004. Trypanosome microtubule-associated protein p15 as a vaccine for the prevention of African sleeping sickness. Vaccine, 22: 1007-1015.
CrossRefDirect Link - Reichel, M.P., M.A. Ayanegui-Alcerreca, L.F.P. Gondim and J.T. Ellis, 2013. What is the global economic impact of Neospora caninum in cattle-the billion dollar question. Int. J. Parasitol., 43: 133-142.
CrossRefDirect Link - Rogers, D.J. and S.E. Randolph, 2006. Climate change and vector-borne diseases. Adv. Parasitol., 62: 345-381.
CrossRefPubMedDirect Link - Sathish, K., R. Sriraman, B.M. Subramanian, N.H. Rao, K. Balaji, M.L. Narasu and V.A. Srinivasan, 2011. Plant expressed EtMIC2 is an effective immunogen in conferring protection against chicken coccidiosis. Vaccine, 29: 9201-9208.
CrossRef - Sathish, K., R. Sriraman, B.M. Subramanian, N.H. Rao and B. Kasa et al., 2012. Plant expressed Coccidial antigens as potential vaccine candidates in protecting chicken against Coccidiosis. Vaccine, 30: 4460-4464.
Direct Link - Sharman, P.A., N.C. Smith, M.G. Wallach and M. Katrib, 2010. Chasing the golden egg: Vaccination against poultry coccidiosis. Parasite Immunol., 32: 590-598.
CrossRefDirect Link - Shirley, M.W. and P. Bedrnik, 1997. Live attenuated vaccines against avian coccidiosis: Success with precocious and egg-adapted lines of Eimeria. Parasitol. Today, 13: 481-484.
CrossRef - Shirley, M.W., A.L. Smith and F.M. Tomley, 2005. The biology of avian Eimeria with an emphasis on their control by vaccination. Adv. Parasitol., 60: 285-330.
CrossRef - Shkap, V., A.J. de Vos, E. Zweygarth and F. Jongejan, 2007. Attenuated vaccines for tropical theileriosis, babesiosis and heartwater: The continuing necessity. Trends. Parasitol., 23: 420-426.
CrossRef - Silva, M.S., D.M. Prazeres, A. Lanca, J. Atouguia and G.A. Monteiro, 2009. Trans-sialidase from Trypanosoma brucei as a potential target for DNA vaccine development against African trypanosomiasis. Parasitol. Res., 105: 1223-1229.
CrossRef - Stephan, B., M. Rommel, A. Daugschies and A. Haberkorn, 1997. Studies of resistance to anticoccidials in Eimeria tenella field isolates and pure Eimeria strains. Vet. Parasitol., 69: 19-29.
CrossRef - Stern, A.M. and H. Markel, 2005. The history of vaccines and immunization: Familiar patterns, new challenges. Health Affairs, 24: 611-621.
CrossRefDirect Link - Streatfield, S.J., 2006. Mucosal immunization using recombinant plant-based oral vaccines. Methods, 38: 150-157.
CrossRef - Schetters, T., 2005. Vaccination against canine babesiosis. Trends. Parasitol., 21: 179-184.
CrossRef - Vanhamme, L., E. Pays, R. McCulloch and J.D. Barry, 2001. An update on antigenic variation in African trypanosomes. Trends. Parasitol., 17: 338-343.
CrossRef - Vercruysse, J., D.P. Knox, T.P. Schetters and P. Willadsen, 2004. Veterinary parasitic vaccines: Pitfalls and future directions. Trends Parasitol., 20: 488-492.
CrossRef - Vercruysse, J., T.P. Schetters, D.P. Knox, P. Willadsen and E. Claerebout, 2007. Control of parasitic disease using vaccines: An answer to drug resistance? Rev. Sci. Tech., 26: 105-115.
Direct Link - Verma, R. and P. Khanna, 2012. Development of Toxoplasma gondii vaccine: A global challenge. Hum Vaccin Immunother, Vol. 9.
Direct Link - Verma, A.K., B.C. Pal, C.P. Singh, U. Jain, S.K. Yadav and Mahima, 2008. Studies of the outbreaks of foot and mouth disease in Uttar Pradesh, India, between 2000 and 2006. Asian J. Epidemiol., 1: 40-46.
CrossRefDirect Link - Verma, A.K., A. Kumar, K. Dhama, R. Deb, A. Rahal, Mahima and S. Chakraborty, 2012. Leptospirosis-Persistence of a dilemma: An overview with particular emphasis on trends and recent advances in vaccines and vaccination strategies. Pak. J. Biol. Sci., 15: 954-963.
CrossRefDirect Link - Verma, A.K., A. Kumar, Mahima and Sahzad, 2012. Epidemiology and diagnosis of foot-and-mouth disease: A review. Indian J. Anim. Sci., 82: 543-551.
Direct Link - Vermeulen, A.N., 1998. Progress in recombinant vaccine development against coccidiosis A review and prospects into the next millennium. Int. J. Parasitol., 28: 1121-1130.
CrossRef - Vermeulen, A.N., D.C. Schaap and T.P.M. Schetters, 2001. Control of coccidiosis in chickens by vaccination. Vet. Parasitol., 100: 13-20.
CrossRefPubMedDirect Link - Von Samson-Himmelstjerna, G. and W. Blackhall, 2005. Will technology provide solutions for drug resistance in veterinary helminthes. Vet. Parasitol., 132: 223-239.
CrossRefDirect Link - Walker, P.S., T. Scharton-Kersten, E.D. Rowton, U. Hengge, A. Bouloc, M.C. Udey and J.C. Vogel, 1998. Genetic immunization with glycoprotein 63 cDNA results in a helper T cell type 1 immune response and protection in a murine model of leishmaniasis. Human Gene. Ther., 9: 1899-1907.
Direct Link - Wallach, M.G., U. Ashash, A. Michael and N.C. Smith, 2008. Field application of a subunit vaccine against an enteric protozoan disease. PLoS ONE, Vol. 3.
Direct Link - Wallach, M., 1997. The importance of transmission-blocking immunity in the control of infections by apicomplexan parasites. Inter. J. Parasitol., 27: 1159-1167.
CrossRef - Wallach, M., N.C. Smith, M. Petracca, C.M. Miller, J. Eckert and R. Braun, 1995. Eimeria maxima gametocyte antigens: Potential use in a subunit maternal vaccine against coccidiosis in chickens. Vaccine, 13: 347-354.
CrossRef - Wang, H.L., Y.Q. Li, L.T. Yin, X.L. Meng and M. Guo et al., 2013. Toxoplasma gondii Protein Disulfide Isomerase (TgPDI) is a novel vaccine candidate against toxoplasmosis. PLoS One, Vol. 8.
Direct Link - Weston, J.F., C. Heuer and N.B. Williamson, 2012. Efficacy of a Neospora caninum killed tachyzoite vaccine in preventing abortion and vertical transmission in dairy cattle. Prev. Vet. Med., 103: 136-144.
CrossRef - Xu, D. and F.Y. Liew, 1994. Genetic vaccination against leishmaniasis. Vaccine, 12: 1534-1536.
CrossRef - Xu, D. and F.Y. Liew, 1995. Protection against leishmaniasis by injection of DNA encoding a major surface glycoprotein, gp63, of L. major. Immunology, 84: 173-178.
Direct Link - Yin, H., J. Luo and W. Lu, 2008. Control of tropical theileriosis with attenuated schizont vaccine in China. Vaccine, Vol. 26.
Direct Link - Dhama, K., M.Y. Wani, R. Deb, K. Karthik and R. Tiwari et al., 2013. Plant based oral vaccines for human and animal pathogens-a new era of prophylaxis: Current and future prospective. J. Exp. Biol. Agric. Sci., Vol. 1.
Direct Link