
Amir Husni
Department of Fisheries, Faculty of Agriculture, Universitas Gadjah Mada, Jl. Flora Gedung A4 Bulaksumur Yogyakarta, 55281, Indonesia
Renita Wijayanti
Department of Fisheries, Faculty of Agriculture, Universitas Gadjah Mada, Jl. Flora Gedung A4 Bulaksumur Yogyakarta, 55281, Indonesia
Ustadi
Department of Fisheries, Faculty of Agriculture, Universitas Gadjah Mada, Jl. Flora Gedung A4 Bulaksumur Yogyakarta, 55281, Indonesia
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 8 | Page No.: 515-520







ABSTRACT
The objective of this study was to determine the potential of polyphenols and phlorotannins extract from Padina pavonica in inhibiting α-amylase and α-glucosidase. Total phenolic content of P. pavonica was 17.63±0.25 μg GAE/mg extract and total phlorotannins content was 1.94±0.19 μg PGE/mg extract. The results showed that polyphenols (IC50 = 0.36±0.05 mg mL-1) and phlorotannins (IC50 = 0.25±0.02 mg mL-1) can inhibit α-amylase, but lower than acarbose (IC50 = 0.06±0.00 mg mL-1). Polyphenols (IC50 = 26.57±2.08 mg mL-1) and phlorotannins (IC50 = 34.40±5.93 mg mL-1) can also inhibit α-glucosidase but also lower than acarbose (IC50 = 1.74±0.25 mg mL-1). Both polyphenol and phlorotannin extracts from P. pavonica have higher inhibitory activity of α-amylase than α-glucosidase. There were high correlations between total phenolic and phlorotannin contents with inhibition activity of α-amylase and α-glucosidase.
PDF Abstract XML References Citation
Received: November 29, 2014;
Accepted: February 06, 2015;
Published: March 25, 2015
How to cite this article
Amir Husni, Renita Wijayanti and Ustadi, 2014. Inhibitory Activity of α-Amylase and α-Glucosidase by Padina pavonica Extracts. Journal of Biological Sciences, 14: 515-520.
DOI: 10.3923/jbs.2014.515.520
URL: https://scialert.net/abstract/?doi=jbs.2014.515.520
DOI: 10.3923/jbs.2014.515.520
URL: https://scialert.net/abstract/?doi=jbs.2014.515.520
INTRODUCTION
Diabetes Mellitus (DM) is a group of metabolic diseases characterized by hyperglycemia resulting from defects in insulin secretion, insulin action, or both (American Diabetes Association, 2012). Chronic hyperglycemia in diabetes is related to long-term damage, dysfunction or failure of several organs of the body, especially the eyes, nerves, heart and blood vessels (Elekofehinti et al., 2013). WHO (2012) reported that approximately 347 million people in the world suffer from DM. The International Diabetes Federation (IDF., 2012) estimates that in 2030, people with diabetes would rise to 438 million.
Brown sea weed Padina pavonica is a species commonly found and abundant in the world. Seaweeds contain polysaccharides, proteins, peptides, amino acids, lipids, minerals and some vitamins. Polyphenols of seaweed are used as cosmetics and pharmacologicals as antioxidants, protection from radiation, antibiotics, anti-inflammatory, hypo-allergenic, antibacterial and antidiabetic (Holdt and Kraan, 2011). Ethanol extract of brown seaweed Sargassum pallidum has antioxidant activity (Ye et al., 2009). Phenol extract of seaweed Palmaria, Ascophyllum and Alaria able to inhibit the activity of α-amylase and α-glucosidase that can lower blood glucose levels (Nwosu et al., 2011). Seaweed also has a high content of antioxidants that can be used to ward off free radicals that increase due to the condition of hyperglycemia in patients with diabetes mellitus (Firdaus et al., 2010).
Research information on the antidiabetic activity of seaweed P. pavonica is lacking. The purpose of this study was to determine the activity of P. pavonica extracts in inhibiting α-amylase and α-glucosidase as an antidiabetic agent.
MATERIALS AND METHODS
Materials: The main material used in this study was seaweed Padina pavonica obtained from coastal Gunungkidul Yogyakarta, Indonesia. The materials used for the extraction of polyphenols and phlorotannin namely: Methanol (E. Merck), Folin-Ciocalteu’s reagent (E. Merck), sodium carbonate (E. Merck), acid gallate (E. Merck). The materials were used to analyze the α-amylase inhibition activity: α-amylase from Bacillus sp. type II-A (Sigma-Aldrich), essential oils (E. Merck), 3,5-dinitrosalicylate (DNS) acid (Sigma-Aldrich) and the materials to analyze the α-glucosidase inhibition activity was: α-glucosidase from Saccharomyces cerevisiae type I (Sigma-Aldrich), p-nitrophenyl-α-D-glucopyranocide (Sigma-Aldrich) and acarbose (Bayer Pharmaceuticals).
Seaweed extractions: Polyphenols extraction was conducted as described by Kang et al. (2010) with modification that the seaweed was washed thoroughly and dried at room temperature for 3-4 days. Dried seaweed then cut into small pieces of ±0.5 cm in size, using scissors. A total of 40 g of seaweed pieces was put into Erlenmeyer flask that has been covered with aluminum foil and macerated using 600 mL of methanol 50% for 48 h at room temperature. After that, the extract was filtered using a coarse filter paper and then centrifuged at 2500 rpm, 20°C, for 10 min. Methanol was evaporated by using a rotary evaporator at 40°C and then the extract was freeze dried and stored at -20°C.
Phlorotannin extract was obtained by extraction of the seaweed using methods as described by Al-Mola (2009). A total of 40 g of seaweed pieces put into Erlenmeyer flask that has been covered with aluminum foil and macerated using 600 mL of methanol 50% for 48 h at room temperature. The extract was filtered using a coarse filter paper and then centrifuged at 2500 rpm, 20°C, for 10 min. Methanol was evaporated by using a rotary evaporator at 40°C and was added with 120 mL of methanol, 240 mL of chloroform and 90 mL of distilled water. The third mixture solution was stirred for 20 min. The solution mixture was transferred to a separating funnel and then waited until two layers were formed. The top layer (non-lipid) were taken and partitioned twice with 150 mL of ethyl acetate. The mixture was evaporated, freeze dried and stored at -20°C until used in further research.
Measurement of total polyphenol content: Total Polyphenol Content (TPC) was measured using the method of Kang et al. (2010). For polyphenol standard, a stock solution (1 mg mL-1) of gallic acid (in distilled water) was prepared and the stock was diluted to give working standards of 0, 3.125, 6.25, 12.5, 25, 50, 100 and 200 μg mL-1, respectively. For sample, 5 mg of several seaweeds methanol extract was dissolved in 1 mL of distilled water and diluted to give same concentration of working standards to measure TPC. An aliquot (10 μL) of each sample and standard solutions was mixed with 50 μL of Folin-Ciocalteu’s reagent in a 96-well microplate and incubated for 5 min. The reactant was mixed with 40 μL of sodium carbonate 7.5% and incubated in the dark place for 2 h. Absorbance was measured at 750 nm with a spectrophotometric microplate reader (BioTech Instruments, Inc., Winooski, USA). The TPC was expressed as Gallic Acid Equivalents (GAE) in μg g-1 extract.
Measurement of total phlorotannin content: Total phlorotannin performed using the method of Koivikko et al. (2005) with modifications of the reaction volume and concentration. Phloroglucinol standard solution were made at various concentrations, i.e., 6.25, 12.5, 25, 50 and 100 μg mL-1. Phlorotannin extract was made in a concentration of 1 mg mL-1. Then, the standard solution and each sample of 250 mL was taken and put into a test tube. Test tube plus 250 mL Folin-Ciocalteu’s reagent and 500 mL of Na2CO3 20% and then left to stand for 3 min. The solution was incubated in a test tube in a dark room at room temperature (27oC) for 45 min. Subsequently, the solution was centrifuged at 3500 rpm for 10 min. The supernatant was taken and immediately read the absorbance at 730 nm. Total phlorotannin contents was expressed as phloroglucinol equivalents (PGE) in μg mg-1 extract.
Inhibition of α-amylase activity: Inhibition of α-amylase activity was determined by measuring changes in 3.5-dinitrosalicylate acid into nitro-aminosalicylate acid (catalyzed by α-amylase) using spectrophotometry (Apostolidis et al., 2011). Test solution consisting of 25 mL sample extract at various concentrations and 25 mL of sodium phosphate buffer 0.02 M (pH 6.9 with NaCl 0.006 M) containing 13 U mL-1 of α-amylase. The test solution was mixed and incubated at 37°C for 10 min. After pre-incubation, add 25 mL of soluble starch 1% in sodium phosphate buffer 0.02 M (pH 6.9 with NaCl 0.006 M) and incubated at 37°C for 10 min. Then proceeded with the addition of 50 mL 3,5-dinitrosalicylate acid (DNS) 96 mM as the color reagent solution and incubated for 5 min in boiling water. The solution was cooled to room temperature and the absorbance of the solution was read at a wavelength of 550 nm. Absorbance values were used to calculate the inhibition activity of the enzyme as follows:
Where:
| K | = | Absorbance of control-blank |
| S1 | = | Absorbance of sample with enzyme |
| S0 | = | Absorbance of sample without enzyme |
Inhibition of α-glucosidase activity: Inhibition test of α-glucosidase was performed according to the method of Mayur et al. (2010). Test solution consisting of 50 mL phosphate buffer (KH2PO4) 0.1 M pH 7, 25 mL substrate p-nitrophenyl-α-D-glucopyranoside (PNP-G) 0.5 mM, 10 mL sample extract at various concentrations and 25 mL α-glucosidase at a concentration of 0.2 U mL-1. The solution was mixed and incubated at 37°C for 30 min. The reaction was stopped by addition of 100 mL Na2CO3 0.2 M. Inhibition of enzyme activity was measured by the amount of p-nitrophenyl formed by measuring absorbance using a microplate reader at a wavelength of 405 nm. Absorbance values were used to calculate the inhibition activity of the enzyme as follows:
Where:
| K | = | Absorbance of control-blank |
| S1 | = | Absorbance of sample with enzyme |
| S0 | = | Absorbance of sample without enzyme |
Statistical analysis: The data in this study were extract concentration versus percent inhibition of the enzyme, were plotted to obtain the regression equation. The IC50 activity value of P. pavonica extracts against α-amylase and α-glucosidase were obtained from the regression equation. The IC50 values were tested statistically with one-way analysis of variance (ANOVA) using the Statistical Package for Social Sciences (SPSS) at 95%.
RESULTS AND DISCUSSION
Total polyphenol and phlorotannin contents: The result of extraction of polyphenols and phlorotannins along with the content of each compound is shown in Table 1. The total phenol content of P. pavonica was obtained 17.63 mg GAE/mg extract or 1.76%. These results were relatively higher when compared with Khaled et al. (2012) who studied the same species of algae with total phenol content at 10.55-10.76 mg GAE/g extract. However, the results in this study was relatively lower compared to the total phenol content in Padina tetrastromatica (43.75±4.83 mg PGE/g extract) (Raju et al., 2010). Zubia et al. (2007) reported that the total phenol content of the Padina gymnospora was 5.58±0.30%. Damongilala et al. (2013) showed that P. pavonica was extracted using methanol 60% resulted in the total phenol content of 4.98-5.87 mg GAE/g extract. The differences in total phenol content can be influenced by intrinsic factors (age, type, etc.) and extrinsic factors (tidal cycles, salinity, etc.) (Connan et al., 2007).
The results showed that phlorotannin content was lower than phlorotannin content in Sargassum echinocarpum (6.75 mg PGE/g extract) extracted using methanol (Firdaus, 2011). Shibata et al. (2004) stated that the phlorotannin content of the brown algae Eisenia bicyclis, E. cava and E. kurome were 3.1, 3.3 and 3.0%, respectively. Phlorotannin content is specific to each species.
| Table 1: | Yield and total content of polyphenols and phlorotannin extracts from Padina pavonica |
According to Jormalainen and Honkanen (2004), the phlorotannin concentration of the algae is influenced by several factors, including species, age and environment (location, season, waves, presence or absence of light, salinity, UV radiation, the presence or absence of herbivores and nutrients).
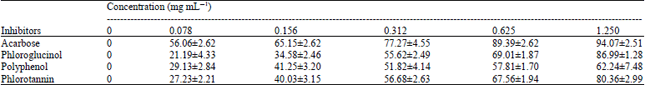
Inhibition of α-amylase activity: Table 2 showed that the highest inhibitory activity was obtained in acarbose (94.07±2.51%). At the same concentration, phloroglucinol showed a smaller inhibitory activity (86.99±1.28%) and the activity was not significantly different compared to phlorotannin of P. pavonica (80.36±2.99%). Phlorotannin at these concentrations showed higher inhibitory activity compared to polyphenols of P. pavonica. This can be caused by many types of phenolic compounds contained in polyphenols while phlorotannin already be more specific compounds, although not yet be called a pure compound. Polyphenols in brown algae are generally in the form of a compound unit was represented by a phloroglucine polymer called phlorotannin which is similar to the condensed tannins (Chkhikvishvili and Ramanov, 2000). According to Lopes et al. (2012), polyphenols consists of flavonoid or gallic acid polymer in terrestrial plants while the seaweed consisting of units of phloroglucinol (1,3,5-trihydroxybenzene).
Table 3 showed the IC50 of inhibition activity of α-amylase by P. pavonica extracts, acarbose and phloroglucinol. The IC50 values from the four test samples ranging from the smallest to largest were acarbose, phlorotannin, phloroglucinol and polyphenols. IC50 values of acarbose was significantly different from all the samples which means acarbose has the ability to inhibit α-amylase which was better than all the samples. There is no significance different between IC50 values of phloroglucinol and phlorotannin. The IC50 values of polyphenols significantly different, compared to all the samples. This indicates that inhibition activity of polyphenols is lower than phloroglucinol and phlorotannin. Based on these results it can be noted that the phlorotannin has the best ability after acarbose. The IC50 values showed higher activity compared to another marine algae but also lower than Ascophyllum nodosum. SenthilKumar and Sudha (2012), reported that IC50 value of the water extracts from marine algae G. edulis, S. policystum, R. corticata and G. lactuca in inhibiting of α-amylase were 83, 60, 67 and 82 μg mL-1, respectively. Nwosu et al. (2011) reported that methanol extract of A. nodosum has a smaller IC50 value (0.1 μg mL-1).
Inhibition of α-glucosidase activity: The inhibition activity of α-glucosidase showed that acarbose have the greatest ability to inhibit α-glucosidase activity, followed by polyphenol extracts of P. pavonica.
| Table 2: | Effect of polyphenols and phlorotannin extracts from Padina pavonica on inhibition activity (%) of α-amylase |
 | |
| Table 3: | Inhibition activities (%) of polyphenol and phlorotannin extracts from Padina pavonica against α-glucosidase |
 | |
| Table 4: | Inhibition activities (IC50) of polyphenol and phlorotannin extracts from Padina pavonica, acarbose and phloroglucinol against α-amylase and α-glucosidase |
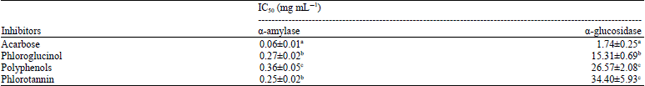 | |
Table 4 showed that increasing on concentrations of P. pavonica extracts, the level of α-glucosidase inhibitory activity also increased. The highest inhibitory activity was found in acarbose (86.91±4.84%). At the same concentration, phloroglucinol showed inhibitory activity, much smaller (36.45±0.41%), followed by polyphenols (29.60±0.39%) and phlorotannin (15.31±2.22%). The inhibition activity of polyphenols on α-glucosidase was similar compared to phlorotannin extracts. This can be caused by both extracts may containing similar phenolic compounds. The IC50 for P. pavonica extract and acarbose was 10.2 and 6.9 mg mL-1, respectively (Table 3). IC50 values of the four compounds that have the ability to inhibit the activity from the largest to the smallest were acarbose, polyphenols, phloroglucinol and phlorotannin. The analysis showed that polyphenols of P. pavonica have ability to inhibit the activity of α-glucosidase similar to acarbose. On the other hand, polyphenols have more inhibitory effects than phloroglucinol.
The level of inhibition activity of α-amylase and α-glucosidase was influenced by the total polyphenol content and phlorotannin. Figure 1a, b showed the relationship between total phenol and phlorotannin contents and the inhibition activity of α-amylase. The relationship between total phenol or phlorotannin contents and the inhibition activity of α-amylase showed a significant correlation (p<0.05), with a correlation coefficient of 0.85 and 0.93, respectively. This showed that the correlation between total phenol or phlorotannin contents and the inhibition activity of α- amylase was high. The coefficient of determination (R2) showed a value of 0.73 and 0.86, respectively which means that the total phenol or phlorotannin content affects α-amylase inhibition activity of 73 and 86%, respectively. Effect of total phenol or phlorotannin did not reach 100%. This is due to the presence of other compounds in the extract of brown algae. Fleury et al. (1994) mentioned that brown algae is mainly included in the order Fucales and Dictyotales containing polyphenolic compounds (phlorotannin), sterols and terpenes. However, more sterol content contained in the order Fucales while terpene content more widely available in order Dictyotales. According to De Paula et al. (2011), Dictyotaceae is a very rich source of secondary metabolites, especially terpenes. Sesquiterpenes compounds found in Padina, Dictyopteris and Taonia.
Figure 2a and 2b shows the relationship between total phenols or phlorotannin contents and the inhibition activity of α-glucosidase. The relationship between total phenols or phlorotannin content and the degree of inhibition activity of α-glucosidase showed a significant correlation (p<0.05), with a correlation coefficient of 0.97 and 0.96, respectively. This shows that the correlation between total phenols or phlorotannin content and the percentage of inhibition of α-glucosidase is also high with coefficients of determination at 0.95 and 0.93, respectively.
Phenols is one of the bioactive components that can inhibit the action of α-amylase. Polyphenols inhibit the enzyme in the breakdown of carbohydrates into glucose. The content of phenol has an inhibitory effect on α-amylase through bond hydroxylation and ring substitution on β bond. The principle is similar to acarbose inhibition i.e., by generating delays and disaccharide carbohydrate hydrolysis and absorption of glucose and inhibits the metabolism of sucrose into glucose and fructose (You et al., 2012). On the other hand phlorotannin is one of the phenolic components which can inhibit the work of the α-amylase. As polyphenols, phlorotannin inhibit enzymatic work in the breakdown of carbohydrates into glucose. The principle also is similar to acarbose inhibition, that is to produce a delay in hydrolysis and absorption of carbohydrates and disaccharides to inhibit the metabolism of glucose and sucrose into glucose and fructose (You et al., 2012).
Seaweed P. pavonica has potential as an antidiabetic substance. Inhibition activity of polyphenols extract of P. pavonica to inhibit the activity of α-amylase and α-glucosidase was higher than the phlorotannin extract. Both polyphenols and phlorotannin extracts are more potent to inhibit α-amylase activity than α-glucosidase. There were high correlations between total phenolic and phlorotannins with inhibition activity of α-amylase and α-glucosidase.
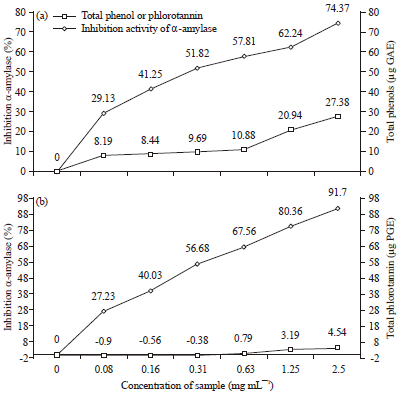 | |
| Fig. 1(a-b): | Relationship between total phenols (a) or phlorotannin (b) and inhibition activity of α-amylase |
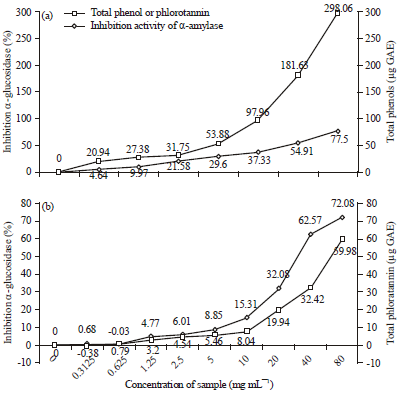 | |
| Fig. 2(a-b): | Relationship between total phenols (a) or phlorotannin (b) and inhibition activity of α-glucosidase |
CONCLUSION
Seaweed P. pavonica has potential as an antidiabetic substance. The results showed that polyphenols (IC50 = 0.36±0.05 mg mL-1) and phlorotannins (IC50 = 0.25±0.02 mg mL-1) can inhibit α-amylase. Polyphenols (IC50 = 26.57±2.08 mg mL-1) and phlorotannins (IC50 = 34.40±5.93 mg mL-1) can also inhibit α-glucosidase. Both polyphenol and phlorotannin extracts from P. pavonica have higher activity for inhibiting activity of α-amylase than α-glucosidase. There were high correlation between total phenolics and phlorotannins with inhibition activity of α-amylase and α-glucosidase.
ACKNOWLEDGMENT
This research was supported by Research Grants Flagship Universitas Gadjah Mada through DIPA UGM 2013 Number LPPM-UGM/1406/LIT/2013.
REFERENCES
- American Diabetes Association, 2012. Diagnosis and classification of diabetes mellitus. Diabetes Care, 35: S64-S71.
CrossRefDirect Link - Al-Mola, H.F., 2009. Antibacterial activity of crude extracts and phlorotannin isolated from the diatom Cymbella spp. J. Pharm. Res., 2: 304-308.
Direct Link - Apostolidis, E., P.D. Karayannakidis, Y.I. Kwon, C.M. Lee and N.P. Seeram, 2011. Seasonal variation of phenolic antioxidant-mediated α-glucosidase inhibition of Ascophyllum nodosum. Plant Foods Hum. Nutr., 66: 313-319.
CrossRefDirect Link - Chkhikvishvili, I.D. and Z.M. Ramazanov, 2000. Phenolic substances of brown algae and their antioxidant activity. Applied Biochem. Microbiol., 36: 289-291.
CrossRefDirect Link - Connan, S., E. Deslandes and E.A. Gall, 2007. Influence of day-night and tidal cycles on phenol content and antioxidant capacity in three temperate intertidal brown seaweeds. J. Exp. Mar. Biol. Ecol., 349: 359-369.
CrossRefDirect Link - Damongilala, L.J., S.B. Widjanarko, E. Zubaidah and M.R.J. Runtuwene, 2013. Antioxidant activity against methanol extraction of Eucheuma cotonii and E. spinosum collected from North Sulawesi waters, Indonesia. Food Sci. Qual. Manage., 17: 7-13.
Direct Link - Elekofehinti, O.O., J.P. Kamdem, I.J. Kade, J.B.T. Rocha and I.G. Adanlawo, 2013. Hypoglycemic, antiperoxidative and antihyperlipidemic effects of saponins from Solanum anguivi Lam. fruits in alloxan-induced diabetic rats. South Afr. J. Bot., 88: 56-61.
CrossRefDirect Link - Firdaus, M., I. Sarwono, D. Muchtadi, M. Astawan, S. Waspadji, T. Wresdiyati and S.K. Setyawati, 2010. [Effects of brown algae extract to function of aorta endothelium cells in diabetic rats]. Majalah Farmasi Indonesia, 21: 151-157, (In Indonesian).
Direct Link - Fleury, B.G., A. Kelecom, R.C. Pereira and V.L. Teixeira, 1994. Polyphenols, terpenes and sterols in Brazilian Dictyotales and Fucales (Phaeophyta). Botanica Marina, 37: 457-462.
CrossRefDirect Link - Holdt, S.L. and S. Kraan, 2011. Bioactive compounds in seaweed: Functional food applications and legislation. J. Applied Phycol., 23: 543-597.
CrossRefDirect Link - Jormalainen, V. and T. Honkanen, 2004. Variation in natural selection for growth and phlorotannins in the brown alga Fucus vesiculosus. J. Evol. Biol., 17: 807-820.
CrossRefDirect Link - Kang, C., Y.B. Jin, H. Lee, M. Cha and E.T. Sohn et al., 2010. Brown alga Ecklonia cava attenuates type 1 diabetes by activating AMPK and Akt signaling pathways. Food Chem. Toxicol., 48: 509-516.
CrossRefDirect Link - Khaled, N., M. Hiba and C. Asma, 2012. Antioxidant and antifungal activities of Padina pavonica and Sargassum vulgare from the Lebanese Mediterranean coast. Adv. Environ. Biol., 6: 42-48.
Direct Link - Koivikko, R., J. Loponen, T. Honkanen and V. Jormalainen, 2005. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga Fucus vesiculosus, with implications on their ecological functions. J. Chem. Ecol., 31: 195-212.
CrossRefDirect Link - Raju, S.N., D.S. Kumar, D. Banji, A. Harani, P. Shankar, P.A. Kumar and V.R. Reddy, 2010. Inhibitory effects of ethanolic extract of Piper trioicum on amylase, lipase and α-glucosidase. Der Pharmacia Lettre, 2 : 237-244.
Direct Link - Lopes, G., C. Sousa, L.R. Silva, E. Pinto and P.B. Andrade et al., 2012. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE, Vol. 7.
CrossRefDirect Link - Mayur, B., S. Sandesh, S. Shruti and S. Sung-Yum, 2010. Antioxidant and α-glucosidase inhibitory properties of Carpesium abrotanoides L. J. Med. Plant Res., 4: 1547-1553.
Direct Link - Nwosu, F., J. Morris, V.A. Lund, D. Stewart, H.A. Ross and G.J. McDougall, 2011. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem., 126: 1006-1012.
CrossRefDirect Link - De Paula, J.C., M.A. Vallim and V.L. Teixeira, 2011. What are and where are the bioactive terpenoids metabolites from Dictyotaceae (Phaeophyceae). Revista Brasileira Farmacognosia, 21: 216-228.
CrossRefDirect Link - SenthilKumar, P. and S. Sudha, 2012. Evaluation of alpha-amylase and alpha-glucosidase inhibitory properties of selected seaweeds from Gulf of Mannar. Int. Res. J. Pharm., 3: 128-130.
Direct Link - Shibata, T., S. Kawaguchi, Y. Hama, M. Inagaki, K. Yamaguchi and T. Nakamura, 2004. Local and chemical distribution of phlorotannins in brown algae. J. Applied Phycol., 16: 291-296.
CrossRefDirect Link - Ye, H., C. Zhou, Y. Sun, X. Zhang, J. Liu, Q. Hu and X. Zeng, 2009. Antioxidant activities in vitro of ethanol extract from brown seaweed Sargassum pallidum. Eur. Food Res. Technol., 230: 101-109.
CrossRefDirect Link - You, Q., F. Chen, X. Wang, Y. Jiang and S. Lin, 2012. Anti-diabetic activities of phenolic compounds in muscadine against alpha-glucosidase and pancreatic lipase. LWT-Food Sci. Technol., 46: 164-168.
CrossRefDirect Link - Zubia, M., D.Y. Robledo and Y. Freile-Pelegrin, 2007. Antioxidant activities in tropical marine macroalgae from the Yucatan Peninsula, Mexico. J. Applied Phycol., 19: 449-458.
CrossRefDirect Link