
Keukeu K. Rosada
School of Life Sciences and Technology, Institute of Technology Bandung, Jl. Ganesa 10 Bandung, Indonesia
Nur F. Afianti
School of Life Sciences and Technology, Institute of Technology Bandung, Jl. Ganesa 10 Bandung, Indonesia
Dea Indriani Astuti
School of Life Sciences and Technology, Institute of Technology Bandung, Jl. Ganesa 10 Bandung, Indonesia
Gede Suantika
School of Life Sciences and Technology, Institute of Technology Bandung, Jl. Ganesa 10 Bandung, Indonesia
Pingkan Aditiawati
School of Life Sciences and Technology, Institute of Technology Bandung, Jl. Ganesa 10 Bandung, Indonesia
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 6 | Page No.: 414-424







ABSTRACT
Bacterial communities in planktonic cells and biofilm at Saguling Hydro Power were investigated using Denaturing Gradient Gel Electrophoresis (DGGE) technique. Physicochemical characteristic of aqueous medium indicated that the water source was classified as moderately polluted (eutrophic). The study inferred that isolates in biofilm mostly derived from the planktonic cells and the Mann-Withney U tests showed that there was no significantly difference between the bacterial communities of planktonic cells and biofilm. The 16S rRNA sequences revealed that bacteria recovered from the planktonic cells were affiliated with Betaproteobacteria and Bacteroidetes phyla. Meanwhile, the sequences bacteria revealed from biofilm were closed to Betaproteobacteria, Alphaproteobacteria and Gammaproteobacteria groups. Overall, the majority of microbes identified from the two samples were belong to Betaproteobacteria group.
PDF Abstract XML References Citation
Received: April 01, 2014;
Accepted: May 06, 2014;
Published: August 08, 2014
How to cite this article
Keukeu K. Rosada, Nur F. Afianti, Dea Indriani Astuti, Gede Suantika and Pingkan Aditiawati, 2014. Bacterial Community Structures of Planktonic Cells and Biofilm at Saguling Hydro Power using Denaturing Gradient Gel Electrophoresis (DGGE). Journal of Biological Sciences, 14: 414-424.
DOI: 10.3923/jbs.2014.414.424
URL: https://scialert.net/abstract/?doi=jbs.2014.414.424
DOI: 10.3923/jbs.2014.414.424
URL: https://scialert.net/abstract/?doi=jbs.2014.414.424
INTRODUCTION
The deterioration of a metal due to microbial activity is generally termed as microbiologically influenced corrosion or biocorrosion (Puyate and Rim-Rukeh, 2008). Biocorrosion is an electrochemical process where participation of microorganisms is able to initiate, facilitate or accelerate corrosion reaction without changing its electrochemical nature (Rim-Rukeh, 2012). Owing to its economic and environmental importance, biocorrosion has been the subject of extensive studies for the last decades (Puyate and Rim-Rukeh, 2008).
Today, biocorrosion have been reported in many systems such as gas industry pipelines (Zhu et al., 2003), oilfield materials (Akpabio et al., 2011) and drinking water distribution systems (USEPA, 1992), including Saguling Hydro Power facilities. Saguling Hydro Power is a power station that included in the electricity supply system of Java and Bali islands. The source of water used by this power plant comes from Saguling Dam that damming the Citarum river, the largest watershed in West Java. Recently, water quality of Saguling Dam is decreased and polluted by various wastes, particularly organic and inorganic matter, due to many kinds of people activities such as domestic, agriculture, animal husbandry and industry at the upper part of the river (Farida et al., 2006). This is a favorable condition for bacterial growth. According to the Environment Protection Agency (USEPA, 1992), water is the main requirement for microorganisms to grow and also for corrosion to occur. In addition, other components that are prerequisite for the presence of microbes are carbon source, energy source, electron donor and electron acceptor (Beech and Flemming, 2000; Hamilton, 2003). Puyate and Rim-Rukeh (2008) stated that those physicochemical and biological interaction between a metallic material and its environment can lead to corrosion.
In nature, bacterial induced corrosion do not work solely but rather in a consortium (Huggins, 2004). According to Beech and Flemming (2000), biocorrosion refers to the influence of microorganisms on the kinetics of corrosion processes due to the attachment of those microorganisms on the metal surface called biofilm. Biofilms are extremely complex microbial ecosystems consisting of microorganisms attached to a surface and embedded in an organic polymer matrix of microbial origin (Percival et al., 2011). Biofilm formation on the metallic surface plays a critical role in creating a microenvironment that is very different from the surrounding environment (Akpabio et al., 2011). The change of pH, Dissolved Oxygen (DO), organic and inorganic compounds of those microenvironment may cause an electrochemical reaction that increase the rate of corrosion (Pope and Zintel, 1988). According to Sutherland (2001) and Branda et al. (2005), chemical and physiological structure of biofilm varies depend on the microorganisms that occupied and may be the characteristics of the surrounding environment determined by the concentration of the substrates that are available (Wimpenny and Colasanti, 1997). Therefore, in order to control the rate of corrosion because of the existence of biofilm, further study on biofilms including the microbial community residing in those biofilms is recommended.
Most of microorganisms in nature, including those within biofilms, live in mixed populations. PCR-based molecular genetic techniques are very useful in studying microbial diversity since unculturable as well as culturable organisms can be investigated. One such technique is Denaturing Gradient Gel Electrophoresis (DGGE) which separates PCR-amplified community 16S rRNA (and other gene) sequences on the basis of G+C content (Welsh and McLean, 2007). This method is rapid and affordable, allowing multiple samples to be processed simultaneously (Green et al., 2009) and has been extensively used to monitor bacterial communities in space and time (Van der Gucht et al., 2001). In addition, unlike other community fingerprinting techniques, bands from DGGE gels can be excised and sequenced to identify the community members. Thus DGGE can be used to describe overall microbial diversity as well as to identify individual community members (Welsh and McLean, 2007).
In this study, we investigated the community composition of the bacteria present in the Power House of Saguling Hydro Power by Denaturing Gradient Gel Electrophoresis (DGGE) and compared the bacterial composition in biofilm of metal sufaces to those in water sample (planktonic cells) that surrounding the biofilm. The relationship between the bands generated by PCR-DGGE of biofilm and planktonic cells was analyzed graphically as a dendrogram using hierarchical cluster analyses. Furhermore, a statistical test has been applied to identify significant difference between the two samples.
MATERIALS AND METHODS
Site and sample collection: Saguling Hydro Power is located in West Bandung Regency, West Java Province, Indonesia (S. 06°51'49.8"E. 107°20'57", the altitude 643 m). Biofilms and water samples were taken from the Power House of Saguling Hydro Power in February 2013. Biofilm samples were gently scrapped from metal surfaces and composited in a sterile tube before stored in an ice-chest. For assessing planktonic cells diversity, water sample was collected in one litre plastic containers and filtered through a 0.22 μm pore size cellulose membrane filter (Sartorius, Germany). The cells on membrane were re-suspended in 500 μL sodium phosphate buffer and precipitated by centrifugation. Biofilm sample and pellet containing microbial communities were stored at -20°C until use. Upon thawing, the total community DNA was extracted and purified as discribe previously by Fatima et al. (2011) for biofilm sample and Bacosa et al. (2010) for planktonic cells. For water analysis, water samples were collected into several containers with particular preservations (APHA, 2005).
Determination of physicochemical of water sample: At the field and the laboratory, water samples were analysed for the following parameters; pH, temperature, electrical conductivity, Dissolved Oxygen (DO), Biochemical Oxygen Demand (BOD), Chemical Oxygen Demand (COD), nitrate ion (NO3+), nitrite ion (NO2¯), sulphate ion (SO42¯), sulphuric acid (H2SO4), sulphide (S2+) and chloride ion (Cl¯). All analysis were performed according to Standard Methods (APHA, 2005). Those parameters are good environmental impact indicators for biocorrosion problem (Puyate and Rim-Rukeh, 2008; Videla, 1996) due to the prerequisites of the presence of bacteria.
PCR-DGGE analysis: The variable V3 region of 16S rRNA was amplified (Lyautey et al., 2005; Muyzer et al., 1993; Zhu et al., 2003) by nested PCR method (Guan et al., 2012). A nested PCR technique was applied in order to increase the sensitivity and to facilitate the DGGE by analysing fragments of the same length (Boon et al., 2002). The first round amplification was carried out by using primers 27F (5'-AGAGTTTGATCMTGGCTCAG-3') (Lane, 1991) and 902R (5'-GTCAATTCITT-TGAGTTT YARYC-3') (Hodkinson and Lutzoni, 2009). The final concentrations of the different components in the mastermix were: 5 μL 10X buffer GC, 0.5 μL 10 mM dNTP Mix, 1 μL of each primer (10 μM), 0.1 μL KAPA2G Robust DNA Polymerase (5 U μL-1), 15.4 μL ddH2O and 2 μL cDNA. All components were mixed by vortex with total amount 25 μL. PCRs were performed by thermal cycler (Applied Biosystem, BioRad). The temperature cycle for PCR was 3 min of denaturizing at 95°C, 5 min of annealing and 5 min of primer extention at 72°C. During an initial touchdown cycle, the annealing temperature of 24 cycles was continually decreased from 55°C in intervals of 0.4°C per cycle for 30 sec with initial denaturation at 95°C for 30 sec and primer extention at 72°C for 60 sec; 12 additional annealing cycles were performed at 45°C for 30 sec, with 30 sec of denaturizing at 95°C and 2 min of primer extention at 72°C. During the second PCR round, the obtained fragments were re-amplified by using primers 338F (5'-CGCCCGCCGCGCGCGGCGGGC GGGGCGGGGGCACGGGGGGACTCCTACGGGAGGCA GCAG-3') which contains a GC-rich clamp and 518R (5'-ATTACCGCGGCTGCTGG-3') (Duarte et al., 2008; Nakatsu et al., 2000). The final concentration of the different components in the mastermix were: 5 μL 10X buffer GC, 0.5 μL 10 mM dNTP Mix, 1 μL of each primer (10 μM), 0.1 μL KAPA2G Robust DNA Polymerase (5 U μL-1) and 15.4 μL ddH2O. All of the components were mixed by vortex with total amount 23 μL. A 900 bp amplicon was amplified after dillution (100-fold). Furthermore, 2 μL of dilluted amplicon was added to 23 μL of PCR mastermix. Standard PCR was performed in 5 min of denaturizing at 95°C, 3 min of annealing and 5 min of primer extention at 72°C. The annealing temperature of 35 PCR cycles was 53°C for 60 sec, with initial denaturation at 94°C for 60 sec and primer extention at 72°C for 60 sec. After each PCR amplification round, the size of the PCR product was verified on a 1% agarose gel. DGGE of PCR products based on the protocol of Muyzer et al. (1993) was performed using the Bio Rad D Gene systems (Bio-Rad Laboratories). PCR fragments were loaded onto 10% (w/v) polyacrylamide gels in 1X TAE (20 mM Tris, 10 mM acetate, 0.5 mM EDTA, pH 7.4). To seperate the amplified DNA fragments, the polyacrylamide gels were made with denaturing gradients ranging from 30 to 70% (where 100% denaturant contains 7M urea and 40% formamide). The electrophoresis was run for 7 h at 60°C and 70 V. After the electrophoresis, the gels were stained with silver staining method (Creste et al., 2001). Each band in the DGGE gel was excised with a scalpel, added with 35 μL TE buffer, placed in water at a temperature of 60°C for 30 min and then incubated overnight at 4°C. The eluted DNA was amplified using the same primers as previous amplification on the second round without GC clamp sequence. The mastermix components using for re-amplification were 1 μL of each primer (10 μM), 12.5 μL KAPA2G Fast mastermix, 8.5 μL nuclease free water and 2 μL cDNA. PCRs were run at the following condition: 5 min of an initial denaturation at 94°C followed by 25 PCR cycles of 30 sec at 94°C, 15 sec at 55°C, 10 sec at 72°C and 1 min of a final elongation step at 72°C. All of confirmed DGGE bands were subjected to DNA sequencing by the Macrogen Sequencing Service (Korea).
DGGE bands analysis: One way to analyse DGGE fingerprinting patterns is to observe the possible changes in the presence/absence of a single band (Murray et al., 1996). Relatedness of microbial community was determined using similarity coefficient of bands common to two samples. Sorensen index [Cs = 2j/(a+b)] was used to make pairwise calculation of band sharing between samples where, a is the number of bands in sample A, b is the number of bands in sample B and j is the number of bands common to A and B (Nakatsu et al., 2000). The relationship between the bands generated by PCR-DGGE of biofilm and planktonic cells were analyzed graphically as a dendrogram. The clustering algorithms used to calculate the dendrogram was an Unweight Pair Group Method with Arithmetic averages (UPGMA) (Boon et al., 2002; Fromin et al., 2002; Gafan et al., 2005) using Mega5.2. A non parametrics analysis using a Mann-Withney U tests was performed where a p<0.05 was interpreted to be statistically significant (Gafan et al., 2005). Statistical analysis was done using the R (http://www.r-project.org).
Phylogenetic analysis: DNA sequences were analysed and compared to 16S rRNA gene sequences from GenBank database at NCBI (National Centre of Biotechnological Information) using BLAST algorithm for screening of sequencing similarity (Altschul et al., 1997). Phylogenetic trees were constructed from distance matrices by neighbor-joining methods (Saitou and Nei, 1987) with 1.000 bootsrap replicates using Mega5.2.
RESULTS AND DISCUSSION
Water characteristic: Physicochemical characteristic of aqueous medium such as pH, nutrients level, ionic strength and temperature, etc., may play an important role in the rate of microbial attachment to the surface (Kokare et al., 2009). The result of the measured parameters of water samples collected from Saguling Hydro Power are presented in Table 1. Temperature and pH are among the parameters that govern the bacterial growth (Manga et al., 2012). The pH and temperature of water samples were 6.9 and 27.3°C, respectively, indicating a good condition for bacterial growth. The suitable pH values for bacterial growth is within the range 4-9 (Costerton et al., 1995) while the optimum temperature lies between 25-30°C (Booth, 1971). The observed DO level in this study was 5.6 mg L-1.
| Table 1: | Measured parameters of water sample |
 | |
Those level of the dissolved oxygen indicates an environment that promotes the growth of aerobic bacteria. The level of oxygen in an environment plays an important role in corrosion process especially where oxygen reduction is generally the main cathodic reaction (Rim-Rukeh, 2012). The result also showed a high level of BOD and COD values as well as other nutrient contents; nitrate, nitrite and sulphate. Those parameters indicated that organic matter in the water are enough for bacteria to synthesize new cells and generate energy. The high level of sulphate content indicates that there are enough resourse for sulfate-reducing bacteria as the main microorganism causing biocorrosion to grow and transform sulphate (SO42¯) to sulphide (S2+) or hydrogen sulphide (H2S) in the catodhic reaction (Javaherdashti, 2009). Borenstein (1994) stated that region susceptible of biocorrosion would have characteristics of pH below 10.5 and temperature below 98.9°C, with the chemical composition of water environment including sulphate ions >1.0 mg L-1. In addition, according to the saprobial pollution index of The European Water Framework Directive (EUWFD) (EU, 2000), streams with BOD between 2 and 6 mg L-1 are classified as moderately polluted (eutrophic). BOD of this water sample was higher than 6 mg L-1 which is indicating critical loading with organic oxygen-consuming substances. Chloride ion (Cl¯) is composed of a combination of chlorine with a metal. Other parameter that indicating the presence of ions in the water bodies is conductivity. The presence of ions including chloride in the water may contribute to corrosion (Silva, 2013).
DGGE profile: DGGE technique can be used to seperate fragment DNA with over 1.500 bp, however this method would be more efficient to identify the difference of single nucleotide when it was conducted to smaller size of fragments which is around 100-300 bp (Maier et al., 2000). Van der Gucht et al. (2001) have been characterized the bacterioplankton community and its seasonal dynamics in two neighbouring hypertonic lakes by DGGE analysis of short (193 bp) 16S ribosomal DNA polymerase products obtained with primer specific for the domain bacteria. The length of V3 region of 16S rRNA products used as a variable distinguishing in this study is around 190 bp. The result was a pattern of bands, for which the number of bands corresponded to the number of predominant members in the microbial communities (Muyzer and Smalla, 1998).
The DGGE profiles (Fig. 1) showed a clear different pattern between the planktonic bacterial community and the community in the biofilm. Overall, there were 48 different bands derived from both planktonic cells and biofilm, of which 38 bands of them originated from planktonic cells whereas the number of bands derived from biofilm were 31. Moreover, 21 bands among them were found not only in planktonic cells but also in biofilm. The bands generated in this study would reflect the most abundant rRNA gene type from sample or perhaps those strains most applicable to amplification (Polz and Cavanaugh, 1998). Furthermore, ideally one band on the gel corresponds to one species and therefore, the number of bands gives an idea of the richness (S) of the sample. Based on the number of all confirmed bands, it can be assumed that bacterial richness in planktonic cells was higher than that in biofilm. However, similarity coefficient indicated that bacterial community structure of planktonic cells was similar to that of biofilm (Cs = 0.61).
Clustering analysis generated an UPGMA dendrogram (Fig. 2) showed that there are some clades associated with bands that found both in planktonic and biofilm. Four clades are closed to planktonic cells and two clades are closed to biofilm. This inferred that isolates in biofilm mostly derived from the planktonic cells. The Mann-Whitney U test revealed that there was no significantly difference between the bacterial composition of planktonic cells and biofilm. This result means that microbial diversity in biofilm was not significantly different from that in planktonic cells and support the previously result.
In nature, microbial cells that attach to the surfaces and develop a biofilm (Davey and O’Toole, 2000; Hall-Stoodley et al., 2004) occur in response to a variety of environmental signals (O’Toole et al., 2000). The biofilm associated cells are differentiated from suspended counterparts by reduced growth rate, up and down regulation of gene and generation of extracellular polymeric matrix (Donlan, 2002). Genetic approaches have revealed that biofilms are formed through multiple steps, which require intracellular signaling and transcribe different set of genes that differ from planktonic cells (Kokare et al., 2009). As it has been revealed in this study that the isolates in biofilm mostly derived from the planktonic cells, the metabolism between them may be different. Moreno-Paz et al. (2010) research indicated that specific transcriptomic fingerprints can be inferred for both planktonic and biofilm of Leptospirillum spp. cells. The planktonic cells showed a more active TCA cycle whereas the mixed acid fermentation process dominate in the biofilm cells. Additionally, Booth (1971) found that metabolic changes in response to metal exposure in biofilms of Pseudomonas fluorescens were significantly different compared to planktonic cultures. It was demonstrated that planktonic metabolism indicated an oxidative stress response that was characerized by changes to the TCA cycle, glycolysis, pyruvate,nicotinate and niacotinamide metabolism while response in biofilm dominated by shifts in exopolysaccharide related metabolism.
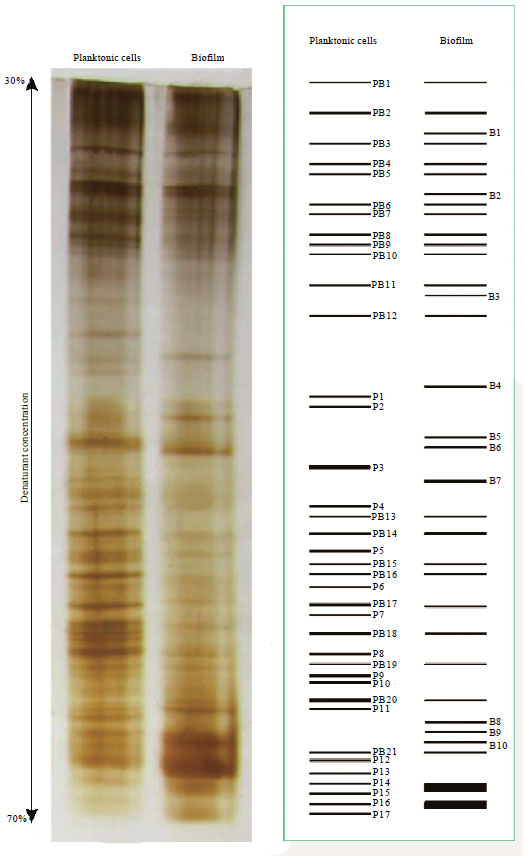 | |
| Fig. 1: | DGGE profiles of planktonic cells and biofilm sampels. P: Planktonic cells, B: Biofilm and PB: Planktonic cells and biofilm |
 | |
| Fig. 2: | Hierarchical cluster analysis. Solid boxes surround ‘the bands that found in both planktonic cells and biofilm that closed to planktonic cells’ clades and broken boxes surround ‘the bands that found in both planktonic cells and biofilm that closed to biofilm’ clades |
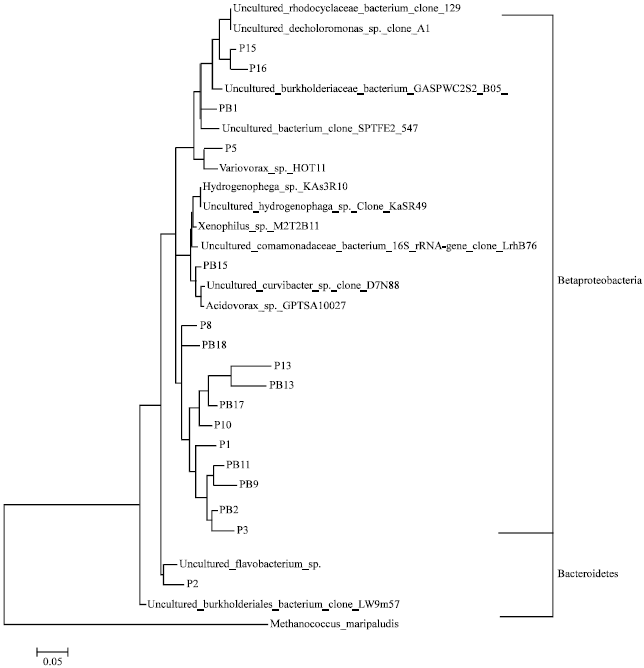 | |
| Fig. 3: | Phylogenetic relationships of the 16S rRNA gene fragment sequences obtained from planktonic cells and related sequences |
Therefore, Booth et al. (2011) suggested that metal stress in biofilms induces a protective response rather than the reactive changes observed for the planktonic cells and that differential metabolic shifts play a role in biofilm specific multimetal resistence and tolerance. The difference of metabolism occuring in biofilm is considered to be caused by gradients of chemicals in the biofilm matrix that not present in planktonic cultures (Stewart and Franklin, 2008).
Phylogenetic and bacterial community: Bacterial communities present in biofilm and planktonic cells were characterized by 16S rRNA gene fragments analysis. All of the sequences shared 84-99% similarity with the sequences from Genbank (data not shown). The majority of the sequences recovered from planktonic cells were affiliated with the Betaproteobacteria group and only one sequence was closed to Bacteroidetes phylum (Fig. 3). Similar to the microbial diversity in the planktonic cells, the majority of the sequences recovered from biofilm were affiliated with the Betaproteobacteria group while other sequences were closed to Alphaproteobacteria and Gammaproteobacteria (Fig. 4).
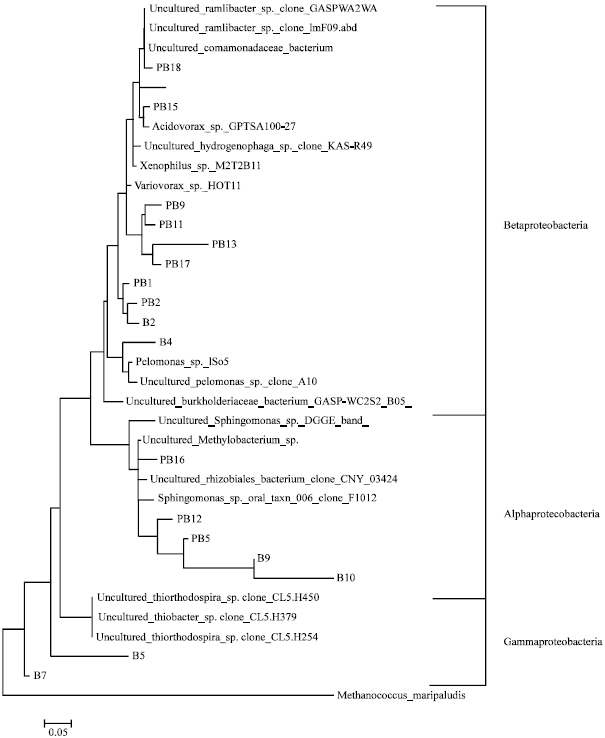 | |
| Fig. 4: | Phylogenetic relationships of the 16S rRNA gene fragment sequences obtained from biofilm samples and related sequences. |
Thus microorganisms that belongs to Betaproteobacteria group mostly found both in planktonic cells and biofilm. Meanwhile, sequences detected not only in planktonic cells but also in biofilm were closed to Betaproteobacteria and Alphaproteobacteria group. Kirchman et al. (2005) noted that Betaproteobacteria were abundant in the freshwater section of estuary whereas, Alphaproteobacteria dominated the bay. According to Elifantz et al. (2005), Betaproteobacteria was the most bacterial group that contribute to Extracellular Polymeric Substances (EPS) formation in freshwater. However, bacteria from Gammaproteobacteria group were the least bacteria that contribute to biofilm formation. DGGE profiles showed that these groups of bacteria appeared from the biofilm sample.
Extracellular polymeric substances produced by microorganisms is a complex mixture of biopolymers particularly composed by polysaccharides, proteins, nucleic acids, lipids and humic substances (Branda et al., 2005; Vu et al., 2009). Those substances play a role in the formation and function of biofilm (Kokare et al., 2009), providing nutrients, protection and construction (Flemming et al., 2007). Donlan and Costerton (2002) noted that production of extracellular polymeric substances is known to be affected by nutrient status of the growth medium.
CONCLUSION
The results have shown that bacterial compositions of planktonic cells and biofilm as well as its relationship can be characterized by using DGGE technique. From DGGE profiles, it can be concluded that isolates in biofilm mostly derived from the planktonic cells and that microbial diversity in biofilm was not significantly different from that in planktonic cells. Furthermore, Betaproteobacteria that is known to be the most bacterial group that contribute to Extracellular Polymeric Substances (EPS) formation in freshwater was the most bacterial group that frequently found both in planktonic cells and biofilm.
REFERENCES
- Akpabio, E.J., E.J. Ekott and M.E. Akpan, 2011. Inhibition and control of microbiologically influenced corrosion in oilfield materials. Environ. Res. J., 5: 59-65.
CrossRefDirect Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Bacosa, H., K. Suto and C. Inoue, 2010. Preferential degradation of aromatic hydrocarbons in kerosene by a microbial consortium. Int. Biodeterior. Biodegrad., 64: 702-710.
CrossRefDirect Link - Boon, N., W. de Windt, W. Verstraete and E.M. Top, 2002. Evaluation of nested PCR-DGGE (denaturing gradient gel electrophoresis) with group-specific 16S rRNA primers for the analysis of bacterial communities from different wastewater treatment plants. FEMS Microbiol. Ecol., 39: 101-112.
CrossRefDirect Link - Booth, S.C., M.L. Workentine, J. Wen, R. Shaykhutdinov and H.J. Vogel et al., 2011. Differences in metabolism between the biofilm and planktonic response to metal stress. J. Proteome Res., 10: 3190-3199.
CrossRefDirect Link - Branda, S.S., A. Vik, L. Friedman and R. Kolter, 2005. Biofilms: The matrix revisited. Trends Microbiol., 13: 20-26.
CrossRefDirect Link - Costerton, J.W., Z. Lewandowski, D.E. Caldwell, D.R. Korber and H.M. Lappin-Scott, 1995. Microbial biofilms. Ann. Rev. Microbiol., 49: 711-745.
CrossRefDirect Link - Creste, S., A.T. Neto and A. Figueira, 2001. Detection of single sequence repeat polymorphisms in denaturing polyacrylamide sequencing gels by silver staining. Plant Mol. Biol. Rep., 19: 299-306.
CrossRefDirect Link - Davey, M.E. and G.A. O'Toole, 2000. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev., 64: 847-867.
PubMed - Donlan, R.M., 2002. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis., 8: 881-890.
CrossRefPubMedDirect Link - Donlan, R.M. and J.W. Costerton, 2002. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev., 15: 167-193.
CrossRefPubMedDirect Link - Duarte, S., C. Pascoal, A. Alves, A. Correia and F. Cassio, 2008. Copper and zinc mixtures induce shifts in microbial communities and reduce leaf litter decomposition in streams. Freshwater Biol., 53: 91-101.
CrossRefDirect Link - Elifantz, H., R.R. Malmstrom, M.T. Cottrell and D.L. Kirchman, 2005. Assimilation of polysaccharides and glucose by major bacterial groups in the delaware estuary. Applied Environ. Microbiol., 71: 7799-7805.
CrossRefDirect Link - EU, 2000. Directive 2000/60/EC of the European parliament and of the council of 23 October 2000 establishing a framework for the Community action in the field of water policy. Off. J. Eur. Commun. L, 327: 1-72.
Direct Link - Farida, W., R. Winurdiastri, S. Wangsaatmaja and L. Boer, 2006. The water quality measurement through PROKASIH program as water environment management policy in Citarum river, West Java Province, Indonesia. Report of Water Pollution Controlling Activity through PROKASIH, West Java Environmental Protection Agency.
- Fatima, F., I. Chaudhary, J. Ali, S. Rastogi and N. Pathak, 2011. Microbial DNA extraction from soil by different methods and its PCR amplification. Biochem. Cell. Arch., 11: 85-90.
Direct Link - Flemming, H.C., T.R. Neu and D.J. Wozniak, 2007. The EPS matrix: The house of biofilm cells. J. Bacteriol., 189: 7945-7947.
CrossRefDirect Link - Fromin, N., J. Hamelin, S. Tarnawski, D. Roesti and K. Jourdain-Miserez et al., 2002. Statistical analysis of Denaturing Gel Electrophoresis (DGE) fingerprinting patterns. Environ. Microbiol., 4: 634-643.
CrossRefDirect Link - Gafan, G.P., V.S. Lucas, G.J. Roberts, A. Petrie, M. Wilson and D.A. Spratt, 2005. Statistical analyses of complex denaturing gradient gel electrophoresis profiles. J. Clin. Microbiol., 43: 3971-3978.
CrossRefDirect Link - Guan, Z.B., Z.H. Zhang, Y. Cao, L.L. Chen, G.F. Xie and J. Lu, 2012. Analysis and comparison of bacterial communities in two types of wheat Qu, the starter culture of Shaoxing rice wine, using nested PCR-DGGE. J. Inst. Brew., 118: 127-132.
CrossRef - Hamilton, W.A., 2003. Microbially influenced corrosion as a model system for the study of metal microbe interactions: A unifying electron transfer hypothesis. Biofouling, 19: 65-76.
CrossRef - Hodkinson, B.P. and F. Lutzoni, 2009. A microbiotic survey of lichen-associated bacteria reveals a new lineage from the rhizobiales. Symbiosis, 49: 163-180.
CrossRef - Javaherdashti, R., 2009. A brief review of general patterns of MIC of carbon steel and biodegradation of concrete. IUFS J. Biol., 68: 65-73.
Direct Link - Kirchman, D.L., A.I. Dittel, R.R. Malmstrom and M.T. Cottrell, 2005. Biogeography of major bacterial groups in the delaware estuary. Limnol. Oceanogr., 50: 1697-1706.
Direct Link - Kokare, C.R., S. Chakraborty, A.N. Khopade and K.R. Mahadik, 2009. Biofilm: Importance and applications. Ind. J. Biol. Technol., 8: 159-168.
Direct Link - Lane, D.J., 1991. 16S/23S rRNA Sequencing. In: Nucleic Acid Techniques in Bacterial Systematic, Stackebrandt, E. and M. Goodfellow (Eds.). John Wiley and Sons, New York, USA., ISBN-13: 9780471929062, pp: 115-175.
Direct Link - Lyautey, E., B. Lacoste, L. Ten-Hage, J.L. Rols and F. Garabetian, 2005. Analysis of bacterial diversity in river biofilms using 16S rDNA PCR-DGGE: Methodological settings and fingerprints interpretation. Water Res., 39: 380-388.
CrossRef - Manga, S.S., S.B. Oyeleke, A.D. Ibrahim, A.A. Aliero and A.I. Bagudo, 2012. Influence of bacteria associated with corrosion of metals. Cont. J. Microbiol., 6: 19-25.
Direct Link - Moreno-Paz, M., M.J. Gomez, A. Arcas and V. Parro, 2010. Environmental transcriptome analysis reveals physiological differences between biofilm and planktonic modes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural microbial community. BMC Genomics, Vol. 11.
CrossRef - Murray, A.E., J.T. Hollibaugh and C. Orrego, 1996. Phylogenetic compositions of bacterioplankton from two California estuaries compared by denaturing gradient gel electrophoresis of 16S rDNA fragments. Applied Environ. Microbiol., 62: 2676-2680.
Direct Link - Muyzer, G., E.C. de Waal and A.G. Uitterlinden, 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied Environ. Microbiol., 59: 695-700.
Direct Link - Muyzer, G. and K. Smalla, 1998. Application of Denaturing Gradient Gel Electrophoresis (DGGE) and Temperature Gradient Gel Electrophoresis (TGGE) in microbial ecology. Antonie Van Leeuwenhoek, 73: 127-141.
CrossRefPubMedDirect Link - Nakatsu, C.H., V. Torsvik and L. Ovreas, 2000. Soil community analysis using DGGE of 16S rDNA polymerase chain reaction products. Soil Sci. Soc. Am. J., 64: 1382-1388.
CrossRef - O'Toole, G., H.B. Kaplan and R. Kolter, 2000. Biofilm formation as microbial development. Annu. Rev. Microbiol., 54: 49-79.
CrossRefPubMedDirect Link - Polz, M.F. and C.M. Cavanaugh, 1998. Bias in template-to-product ratios in multitemplate PCR. Applied Environ. Microbiol., 64: 3724-3730.
PubMedDirect Link - Puyate, Y.T. and A. Rim-Rukeh, 2008. Some Physico-chemical and biological characteristics of soil and water samples of part of the Niger Delta area, Nigeria. J. Applied Sci. Environ. Manage., 12: 135-141.
Direct Link - Rim-Rukeh, A., 2012. Microbiologically influenced corrosion of S45C mild steel in cassava mill effluent. Res. J. Eng. Applied Sci., 1: 284-290.
Direct Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Stewart, P.S. and M.J. Franklin, 2008. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol., 6: 199-210.
CrossRefDirect Link - Hall-Stoodley, L., J.W. Costerton and P. Stoodley, 2004. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol., 2: 95-108.
CrossRefDirect Link - Sutherland, I.W., 2001. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology, 147: 3-9.
CrossRefDirect Link - Van der Gucht, K., K. Sabbe, L. de Meester, N. Vloemans, G. Zwart, M. Gillis and W. Vyverman, 2001. Contrasting bacterioplankton community composition and seasonal dynamics in two neighbouring hypertrophic freshwater lakes. Environ. Microbiol., 3: 680-690.
CrossRef - Vu, B., M. Chen, R.J. Crawford and E.P. Ivanova, 2009. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules, 14: 2535-2554.
PubMed - Welsh, A.K. and R.J.C. McLean, 2007. Characterization of bacteria in mixed biofilm communities using Denaturing Gradient Gel Electrophoresis (DGGE). Curr. Protocol Microbiol.
CrossRefDirect Link - Wimpenny, J.W.T. and R. Colasanti, 1997. A unifying hypothesis for the structure of microbial biofilms based on cellular automaton models. FEMS Microbiol. Ecol., 22: 1-16.
CrossRef - Zhu, X.Y., J. Lubeck and J.J. Kilbane, 2003. Characterization of microbial communities in gas industry pipelines. Applied Environ. Microbial., 69: 5354-5363.
CrossRefDirect Link