
Mihretu Yonas
Gambela Agricultural Research Institute, Gambela, Ethiopia
Weyessa Garedew
Department of Horticulture and Plant Sciences, Jimma University College of Agriculture and Veterinary Medicine, Ethiopia
Adugna Debela
Department of Horticulture and Plant Sciences, Jimma University College of Agriculture and Veterinary Medicine, Ethiopia
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 5 | Page No.: 336-342







ABSTRACT
The objective of the study was to evaluate genetic variability among Okra accessions based on quantitative morphological traits. Twenty five Okra accessions were planted in 2011/2012 at Gambella in randomized complete block design with three replications. Data on 20 quantitative traits were collected and subjected to various statistical analyses. The analysis of variance showed significant differences (p<0.01) among the accessions for all quantitative characters measured. Estimate of phenotypic and genotypic coefficients of variation also showed the presence of variability among the accessions for the majority of the character. High heritability (96.76 and 96.50%) coupled with high genetic advance as percent of mean (106.32 and 97.25%) were recorded for internodes length and plant height, respectively. Correlation study between various quantitative characters highlighted significant association among characters. Fruit yield was positive and highly significant genotypic correlation with fruit length (r = 0.74), average fruit weight (r = 0.62), fruit diameter (r = 0.61), seed per pod (r = 0.56), hundred seed weight (r = 0.68) and number of pod per plant (r = 0.66). Path coefficient analysis at genotypic level revealed that internodes number had highly positive direct effect on fruit yield (p = 6.90) followed by average fruit weight (p = 6.89) which had positively genotypic correlation with yield. The present study indicated a considerable amount of variability for the majority of the quantitative characters in Okra for exploitation. However, it is recommended that the experiment should be repeated at more location and years with more collections to confirm the obtained results.
PDF Abstract XML References Citation
Received: November 17, 2013;
Accepted: February 07, 2014;
Published: April 15, 2014
How to cite this article
Mihretu Yonas, Weyessa Garedew and Adugna Debela, 2014. Variability and Association of Quantitative Characters among Okra (Abelmoschus
esculentus (L.) Moench) Collection in South Western Ethiopia. Journal of Biological Sciences, 14: 336-342.
DOI: 10.3923/jbs.2014.336.342
URL: https://scialert.net/abstract/?doi=jbs.2014.336.342
DOI: 10.3923/jbs.2014.336.342
URL: https://scialert.net/abstract/?doi=jbs.2014.336.342
INTRODUCTION
Okra (Abelmoschus esculentus (L.) Moench) is an economically important vegetable crop grown in tropical and sub-tropical parts of the world which belongs to the family Malvaceae. It is apparently originated in Ethiopia, the mountainous or plateau area of Eritrea and the eastern, higher part of the Anglo-Egyptian Sudan (Aladele et al., 2008). It is widely distributed from Africa to Asia, in Southern Europea, the Mediterranean and all of the America (Oyelade et al., 2003). Okra, commonly known as “lady finger”, is primarily suitable for cultivation as a garden crop as well as on large commercial farms. The crop grows well in hot weather, especially in the regions with warm nights (>20°C) (Ndunguru and Rajabu, 2004). It is sensitive to frost, water logging and drought conditions. Although, there is no complete record on production area and productivity of the crop under Ethiopian condition. It has high diversity in some parts of the country particularly in the Southwestern low lands (550-650 m asl) region (PGRC, 1995). Okra is a multipurpose crop due to its various uses of the fresh leaves, buds, flowers, pods and stems, pods and seeds (Schippers, 2000). For generation, farmers in Gambella and Asossa have been cultivating for its fruit and leaf to use as a food and medicine of different diseases. Nearly 10% of the recommended levels of vitamin B6 and folic acid are also present in a half cup of cooked Okra (FAO, 2004).
Despite its multi-directional importance and utility, there are no improved varieties for cultivation of Okra in Ethiopia. Okra has been collected and maintained for long period of time by Institute of Biodiversity Conservation (IBC) and other research center like Gambella Agricultural Institute (GARI) but they are not yet characterized and their variability is not known. Moreover, little has been previous attempted by breeders in improving the crop in terms of developing core collections for higher yield and quality. The accessions under cultivation, over the years in the various regions across the country are landraces.
Morpho-agronomical characters and diversity analysis of crops is also very essential in order to pave way for genetic improvement. It is first step in any crop improvement program (De Vicente et al., 2005). Adequate characterization for agronomic and morphological traits is necessary to facilitate utilization of germplasm by breeders. Characterization of genetic resources, therefore, refers to the process by which accessions are identified, differentiated or distinguished according to their character. It provides information on diversity within and between crop collections. This enables the identification of unique accessions essential for curators of gene banks (Ren et al., 1995). Moreover, information obtained on genetic relatedness among genetic resources of crop plants is useful, both for breeding and for the purposes of germplasm conservation. Morphological characterization is therefore a highly recommended first step that should be undertaken before more in-depth biochemical or molecular studies are employed in any diversity studies. Therefore, it is imperative to study the variability among the collected accessions. Hence, the objective of the study was to determine the range variability and association of quantitative characters among Okra collection with the ultimate goal of providing a basis for varietal improvement and conservation.
MATERIALS AND METHODS
Experimental site, materials and design: The experiment was conducted at Gambella (8°15' N , 34°35' E with an elevation of 526 meters above sea level). The experiment was conducted on alluvial soil type with a pH of 6.5. The average annual rainfall of the study area was 1020.5 mm year-1 with the average annual minimum and maximum temperature of 20.1 and 35.7°C, respectively. Twenty-five Okra accessions which have been collected by Gambella Agricultural Research Institute from Gambella and Asosa regions were used for the study (Table 1). The accessions were arranged in a Randomized Complete Block Design (RCBD) with 3 replications making a total of 75 plots.
Data collection: Data on 20 quantitative traits were recorded from 9 randomly selected plants from the three middle rows using International Plant Genetic Resources Institute (IPGRI, 1991) descriptor list for Okra species. These includes: Days to emergence, days to 50 percent flowering, leaf length, leaf width, internodes length, stem diameter, peduncle length, No. of epicalyxes, No. of primary branches per stem, days to maturity, plant height, fruit length, fruit diameter, average fruit weight, No. of pod per plant, No. of internodes, No. of ridges on fruit, No. of seeds per pod, hundred seed weight and yield per plot .
| Table 1: | Abelmoschus esculentus accessions and their area of collection |
 | |
Data analysis: Data of quantitative characters were subjected to analysis of variance (ANOVA) using SAS version 9.2 (SAS, 2008) to examine the presence of statistically significant differences among accessions for the characters measured. Assumption of ANOVA was checked. Least Significant Difference (LSD) was employed to make mean separation. Path coefficient analysis, Phenotypic Coefficient of Variation (PCV), Genotypic Coefficient of Variation (GCV), broad-sense heritability (h2) and expected Genetic Advance as percentage to Mean (GAM) was carried out by Genres software (GENRES, 1994).
The variability of each quantitative trait was estimated by simple measures such as mean, range, standard deviation, phenotypic and genotypic variances and coefficients of variation. The phenotypic and genotypic coefficients of variation were computed using the formula suggested by Burton and DeVane (1953). Broad sense heritability values were estimated based on the formula of Falconer and Mackay (1996). Genetic advance in absolute unit (GA) and percent of the mean (GAM), assuming selection of superior 5% of the genotypes was estimated in accordance with the methods illustrated by Johnson et al. (1955) with the standardized selection differential at 5% selection intensity (K = 2.063). Phenotypic correlation (rp), the observable correlation between two variables which includes both genotypes and environmental components between two variables, were estimated using the formula suggested by Johnson et al. (1955) and Singh and Chaudhury (1985). The direct and indirect effect of yield related traits on yield per plot were worked out through path coefficient analysis. The analysis was made following the method suggested by Dewey and Lu (1959).
RESULTS AND DISCUSSION
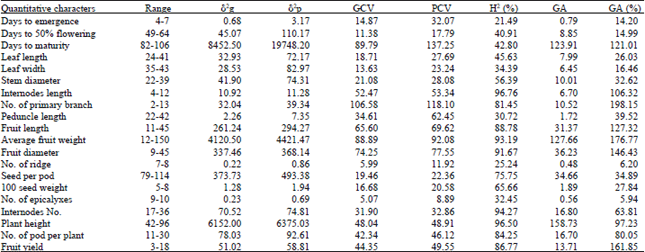
Analysis of variance (ANOVA) for the measured quantitative characters showed highly significant differences (p<0.01) for all the traits. Wide range of variation in character studied was observed. The highest value was almost two times of the minimum value for number of seed per pod, three fold for number of pod per plants, fivefold for fruit diameter and six fold for fruit yield per plot. The highest fruit yield (18 kg plot-1) was recorded from accession AS11, while low yield of (3 kg plot-1) was obtained from AS3 (Table 2).
In this typical study, the highest GCV was recorded form number of primary branches while the lowest was from number of epicalyxes (Table 2). On the other hand, lowest PCV were recorded for only number of epicalyxes indicating a narrow range of variability for these characters and restricting the scope for selection for the crop improvement. Characters showed medium GCV values are days to emergence, days to fifty percent flowering, leaf length and leaf width. A wide variations between PCV and GCV were observed in most of the characters like days to emergency, days to maturity, leaf length, leaf width, peduncle length, stem diameter, of No. primary of branch, indicating high pressure of the environment on these characters and less effect of genetic factors. In general, the study confirmed that the Okra accessions collected from Gambella and Asossa and used in the current study were phenotypically and genotypically diverse. This indicates the existence of large diversity in Okra for quantitative characters implying the need for further collection of germplasm from the untouched geographical areas of the country to broaden the genetic base for future breeding program.
Estimates of heritability and expected genetic advance: Heritability estimates range from 21.49 for days to emergency to 96.76 for internodes length. Maximum heritability was obtained from internodes length followed by plant height, inter node No. average fruit weight and fruit diameter. Therefore, the above characters may respond effectively to selection pressure. On the other hand, days to emergence, No. of ridge, peduncle length, No. of epicalyxes and No. of seed per pod had relatively low heritability (Table 2) indicating limited possibility of improvement of these characters via selection. The other parameters showed moderate heritability estimates. Singh (1990) observed that if heritability of a character is very high around 80% or more, selection for such a character should be fairly easy. This is because there would be a close correspondence between the genotype and phenotype due to relatively small contribution of the environment to the phenotype. But for characters with low heritability, say less than 40%, selection may be considerably difficult or virtually impracticable due to the masking effect of the environment on the characteristics of germplasm. In support to the findings of this investigation, Paiva and De-Costa (1998) reported high heritability for fruit length, diameter, fruit weight, plant height and No. of branches per plant.
| Table 2: | Estimate of components of variance, PCV, GCV, heritability and genetic advance for 20 quantitative characters in Okra studied at Gambela in 2011/2012 |
 | |
On the other study, Dhall et al. (2001) recorded that characters like fruit length, plant height and No. of fruits per plant exhibited high heritability.
The values of genetic advance for different characters of Okra accessions in this particular were different. Genetic advance as per cent mean was categorized as high (≥20%), moderate (10-20%) and low (0-10%) (Johnson et al., 1955). These values are also expressed as percentage of the accession mean for each character so that comparisons could be made among various characters which had different units of measurement. Accordingly, the result indicated that the progress that could be expected from selection of accessions ranged from 5.94% for No. of epicalyxes to 198.15% for No. of primary branches (Table 2). High heritability coupled with high genetic advance is an important instrument for ensuing selection of the best individuals. In this study, high heritability along with high genetic advance as percent of the mean was obtained for internodes length, No. of primary branch, No. of pod per plant, fruit length, average fruit weight, fruit diameter, plant height and fruit yield (Table 2). High GCV along with high heritability and high genetic advance will provide better information than single parameters alone (Ikram et al., 2010). Hence, in this study, number of branch, fruit length, average fruit weight, fruit diameter, plant height and fruit yield exhibited high genotypic coefficients of variation, high heritability together with high genetic advance as percent of means. These results indicate that these characters are controlled by additive gene action; phenotypic selection for the improvement of these characters may be effective.
Genotypic correlation: Fruit yield was positive and highly significant genotypic correlation with fruit length (r = 0.74), average fruit weight (r = 0.62), fruit diameter (r = 0.61), seed per pod (r = 0.56), 100 seed weight (r = 0.68), internodes (r = 0.89 and No. of pod per plant (r = 0.66) (Table 3). It is of interest to note that the significant positive correlation coefficients estimated at genotypic level were also found significant and positive at phenotypic level. Moreover, the significantly higher magnitudes of positive genotypic correlation than the corresponding phenotypic correlation in respect to some of the characters suggest that these characters were genetically controlled.
The present study is agreed with the results reported by Niranjan and Mishra (2003) who observed fruit yield was positively and significantly correlated with No. of fruits per plant, fruit length, No. of seeds per fruit, average fruit weight, plant height and No. of branches per plant at both genotypic and phenotypic levels and also agreed with finding of Singh et al. (2007) who reported high magnitude of genotypic and phenotypic coefficient of variation for No. of branches per plant, plant height, No. of fruits per plant and fruit yield. Therefore, from the correlation analysis average fruit weight, fruit diameter, seed per pod, No. of pod per plant, internodes No., plant height and hundred seed weight, were found to be important yield components. In general, high genotypic correlation suggests that selection directed to one character directly affects the other. Conversely, low or non-correlation suggests independence of association that would be possible to select independently for the two characteristics for diverse directions.
| Table 3: | Genotypic correlation coefficient among 20 traits in 25 Abelmoschus esculentus accessions studied at Gmbella 2011/12 |
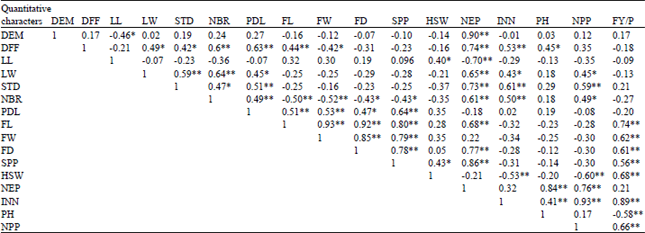 | |
| *Significant 0.05 (r = 0.39) probability level, **Highly significant at 0.01 (r = 0.50) level of probability level. DFF: Days to 50% flowering, DM: Days to maturity, LL: Leaf length (cm), LW: Leaf width (cm), ST: Stem diameter (mm), INL: Internodes length (cm), NBR: No. of primary of branch, PDL: Peduncle length (cm), FL: Fruit length (cm). FW: Average fruit weight (g), FD: Fruit diameter (mm), SPP: Seed per pod, SW: Hundred seed weight (g), NEP: No. of epicalyxes, INN: Internodes No. PH: Plant height (m), NPP: No. of pod per plant, FY/P: Fruit yield per plot (kg) | |
| Table 4: | Phenotypic correlation coefficient among 20 traits in 25 Abelmoschus esculentus accessions studied at Gambella in 2011/2012 |
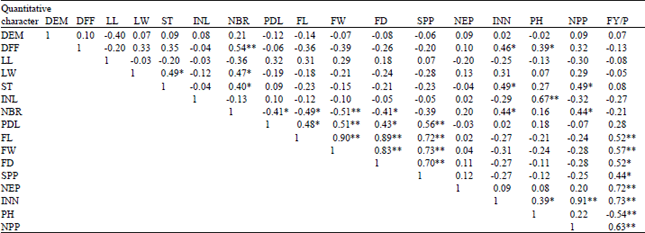 | |
| *Significant 0.05 (r = 0.39) probability level, **Highly significant at 0.01 (r = 0.50) level of probability level, DFF: Days to 50% flowering, DM: Days to maturity, LL: Leaf length (cm), LW: Leaf width (cm), ST: Stem diameter (mm), INL: Internodes length (cm), NBR: No. of primary of branch, PDL: Peduncle length (cm), FL: Fruit length (cm), FW: Average fruit weight (g), FD: Fruit diameter (mm), SPP: Seed per pod, SW:hundred seed weight (g), NEP: No. of epicalyxes, INN: Internodes No. PH: Plant height (m), NPP: No. of pod per plant, FY/P: Fruit yield per plot (kg) | |
| Table 5: | Genotypic direct (bold ) and indirect effects of some characters on fruit yield of Okra accessions studied at Gambella in 2011/2012 |
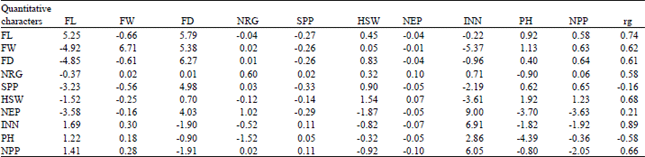 | |
| Residual effect = 0.25, FL: Fruit length (cm), FW: Average fruit weight (g), FD: Fruit diameter (mm), NRG: No. of ridge, SPP: Seed per pod, SW: Hundred seed weight (g), NEP: No. of epicalyxes, INN: Internodes No. PH: Plant height (m), NPP: No. of pod per plant | |
In this study, genotypic correlations were higher than the phenotypic correlations. This may be possibly due to the masking action of genes on the influence of environment in the expression of characters indicating the association is largely due to genetic reason which agrees with the findings of Bello et al. (2006).
Phenotypic correlation: Like that of genotypic correlation fruit length (r = 0.52), average fruit weight (r = 0.57), fruit diameter (r = 0.52), seed per pod (r = 0.44), No. of epicalyxes (r = 0.72), internodes No. (r = 0.73) and No. of pod per plant (r = 0.63) (Table 4) had highly significant positive phenotypic correlation with fruit yield. Similar finding has been reported by Jaiprakashnarayan and Mulge (2004) who noticed that total yield per plant was positively and significantly correlated with No. of fruits per plant, average fruit weight, No. of nodes on main stem, fruit length, plant height at 60 and 100 days after sowing and number of leaves at 45 and 100 days. On the other study, Dhankar and Dhankar (2002) and Bendale et al. (2003) examined 30 Okra genotypes and found that pod length, pod weight, plant height, nodes per plant and No. of pods per plant were positively correlated with the yield observed that fruit yield was significantly and positively correlated with the number of fruits and branches per plant. The investigation indicated that possibilities of developing early, short and high yielding cultivars by exploiting aforesaid associations.
Path coefficient analysis: Path coefficient analysis (Table 5) for some of the characters at genotypic level revealed that internodes number had highly positive direct effect on fruit yield (p = 6.91) followed by average fruit weight (p = 6.71) which had positive genotypic correlation with yield. However, as internodes number becomes higher, it has a negative impact on the plant height. But it has a positive impact on fruit length and fruit diameter which could be a cause for the high correlation coefficient that existed between fruit yield and internodes number (r g = 0.89). Hence, while undertaking selection for fruit yield, one has to consider these two yield components in Okra because when selecting plants with higher No. of fruit, they may produce plant height.
On the other hand, though the direct effect of plant height on fruit yield was negative (p = -4.39), its correlation coefficient was negative (r g = -0.58) as it has high positive indirect effect internodes number and fruit length. The residual effect (h = 0.25) is relatively high indicating that the trait considered in this analysis failed to sufficiently explain the variation in Abelmoschus esculentus yield. This suggests that more yield components should be considered to account for the variation in Abelmoschus esculentus yield.
ACKNOWLEDGMENTS
We are gratefully acknowledged Jimma University College of Agriculture and Veterinary Medicine for financial support and Gambela Agricultural Research Institutes for Technical support for the execution of the study.
REFERENCES
- Aladele, S.E., O.J. Ariyo and R. de Lapena, 2008. Genetic relationship among West African okra (Abelmoschus caillei) and Asian genotypes (Abelmoschus esculentus) using RAPD. Afr. J. Biotechnol., 7: 1426-1431.
Direct Link - Burton, W.G. and E.H. Devane, 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J., 45: 478-481.
CrossRefDirect Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Ikram, U.H., A.A. Khan, F.M. Azhar and U. Ehsan, 2010. Genetic basis of variation for salinity tolerance in okra (Abelmoschus esculentus L.). Pak. J. Bot., 42: 1567-1581.
Direct Link - Johnson, H.W., H.F. Robinson and R.E. Comstock, 1955. Genotypic and phenotypic correlations in soybeans and their implications in selection. Agron. J., 47: 477-483.
CrossRefDirect Link - Ndunguru, J. and A.C. Rajabu, 2004. Effect of okra mosaic virus disease on the above-ground morphological yield components of okra in Tanzania. Sci. Hortic., 99: 225-235.
Direct Link - Oyelade, O.J., B.I.O. Ade-Omowaye and V.F. Adeomi, 2003. Influence of variety on protein, fat contents and some physical characteristics of okra seeds. J. Food Eng., 57: 111-114.
CrossRefDirect Link - Ren, J., J.R. McFerson, R. Li, S. Kresovich and W.F. Lamboy, 1995. Identities and relationships among Chinese vegetable Brassicas as determined by random amplified polymorphic DNA markers. J. Am. Soc. Hortic. Sci., 120: 548-555.
Direct Link
wubadis kenaw Reply
it is very interesting paper!