
Thalisa Yuwa-amornpitak
Department of Biotechnology, Faculty of Technology, Mahasarakham University, Khamrieng, Kantarawichai, 44150, Thailand
Kannika Chookietwattana
Department of Biotechnology, Faculty of Technology, Mahasarakham University, Khamrieng, Kantarawichai, 44150, Thailand
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 4 | Page No.: 284-291







ABSTRACT
Lactic acid producing fungi were screened from soil samples at various temperatures. Several fungal strains were selected from 30 and 35°C and found that no fungal growth at 40 and 45°C. The results from starch hydrolysis and acid testing revealed that the highest lactic acid was produced by Rhizopus sp. LTH23. Effect of various parameters on lactic acid production were investigated. It was found that the enhancing temperature for higher lactic acid production was at 40°C from 5% starch medium and the medium should be adjusted to pH 9. Effect of N-sources on lactic acid production from NH4(SO4)2 and Urea at various concentrations were also studied. The results showed that more lactic acid production from the medium containing Urea than NH4(SO4)2. The optimum concentration of urea was at 7.5 g L-1. The effect of agitation speed on lactic acid production in batch fermentation showed that lactic acid of 20 g L-1 was produced during 60 h of cultivation at an agitation speed of 200 rpm, air flow rate of 0.5 vvm and maintained at a temperature of 40°C. Fed-batch fermentation system, a method for enhancing higher production yield was used. The result was indicated that lactic acid was produced 3 times higher than in the batch processing by adding 25% fresh medium at 10 h of cultivation. Fungal identification by ITS region sequence was used. It was proved that this fungus was 99.85% similar to Rhizopus microspores (Accession No. JX661044.1) from GenBank data base. Then named it as Rhizopus microsporus strain LTH23.
PDF Abstract XML References Citation
Received: October 29, 2013;
Accepted: January 23, 2014;
Published: March 18, 2014
How to cite this article
Thalisa Yuwa-amornpitak and Kannika Chookietwattana, 2014. L-lactic Acid Production from Cassava Starch by Thermotolerant Rhizopus
microsporus LTH23. Journal of Biological Sciences, 14: 284-291.
DOI: 10.3923/jbs.2014.284.291
URL: https://scialert.net/abstract/?doi=jbs.2014.284.291
DOI: 10.3923/jbs.2014.284.291
URL: https://scialert.net/abstract/?doi=jbs.2014.284.291
INTRODUCTION
Due to plastic derived from petrochemical materials provided unfriendly environmental waste for several centuries. An alternative compostable plastic such as polylactic acid (PLA) was discovered around 1890. Polylactic acid (PLA) is the most common bioplastic that worldwide use today. Lactic acid is a basic building block for PLA which was first discovered in 1780 from sour milk by the Swedish chemist Carl Wihelm Scheele. Lactic acid exists as two enantiomers, D(-)-lactic and L-lactic acid. It was known that d-isomer is harmful to humans while L(+)-lactic acid is the safe isomer for food and beverage products and including others application. Lactic acid can be produced either by chemical or fermentation process. Lactic acid production from chemical synthesis method by petrochemical products is racemic mixture of two optical isomers, D(-) and L(+) from. While fermentation process can yield an optically pure form of lactic acid or racemate, depending on microorganisms. There are reported that Lactic Acid Bacteria (LAB) and filamentous fungi are producers of lactic acid. Some species of LAB produces only D(-) or L(+)-lactic acid and some species produce racemate DL-lactic acid (Connolly and Lonnerdal, 2004; Benthin and Villadsen, 1995). Whereas filamentous fungus, Rhizopus sp. produce only L(+)-lactic acid (Ganguly et al., 2007; Huang et al., 2005).
Fungi usage for lactic acid production is benefit over than LAB in various items, such as fungi can be used various sources of renewable materials including sugars, starch, lignocelluloses as substrate (Park et al., 2004; Woiciechowski et al., 1999), fungi need low nutritional requirements than LAB, the removal fungal mycelium from fermentation broth by filtration is cheap and easy. The recovery cost and purification process of lactic acid from fermentation broth by fungi is cheaper and easier than LAB. In addition, fungal biomass can be used in biosorption and purification the effluents (Fourest et al., 1994), for chitosan production (Yoshihara et al., 2003) and improve the feed quality (Kusumaningtyas et al., 2006).
There were reported that many species of Rhizopus, such as R. oryzae, R. arrhizus and R. microsporus can produce lactic acid (Kitpreechavanich et al., 2008). Mostly reported that fungus produced lactic acid at moderated temperature (30°C). However it had been reported that higher temperature about 40-45°C enhanced higher lactic acid production. In this study fungal strains were isolated from soil samples at various temperatures and screened thermotolerant fungi for lactic acid production from cassava starch. The optimum condition in terms of substrate concentration, pH, temperature and N-sources concentration were investigated for lactic acid production. Including batch and fed-batch process for lactic acid production were also examined.
MATERIALS AND METHODS
Fungal screening and storage: The molds were isolated from soil samples on Yeast Medium (YM) agar (3 g L-1 yeast extract, 3 g L-1 malt extract, 5 g L-1 peptone, 10 g L-1 glucose, 15 g L-1 agar) at 30, 35, 40 and 45°C. Fungal strains were maintained on YM agar medium at their screening temperature for 3-5 days before temporarily stored in the refrigerator. For long term storage, a fungal mycelium disc that were punched with 5 mm cork borer and immersed in 15% sterilized glycerol. They were kept at -20°C until used.
Acid production and starch hydrolysis testing: Acid production by fungal strains were tested on acid production medium containing 3 g L-1 yeast extract, 10 g L-1 cassava starch, 15 g L-1 agar and 0.02 g L-1 bromocresol green. Yellow clear zone was measured after incubation at those screening temperatures for 16-24 h. Starch utilization by those fungal strains were tested on starch yeast medium (3 g L-1 yeast extract, 3 g L-1 malt extract, 5 g L-1 peptone, 10 g L-1 cassava starch, 15 g L-1 agar). Clear zone was measured after it was overspread with iodine solution (1 g L, 2 g KI in 300 mL DW and 10 times dilution).
Flask culture and medium: Rice koji medium containing ready cooked rice 15 g, cassava starch 1.5 g, distilled water 5-10 mL 1 bottle-1 was prepared. After sterilized by steam three disc fungal strain were inoculated on rice koji medium and incubated at 35°C for 2 days. Ten grams of koji inoculum was inoculated into a 300 mL flask containing 90 mL starch medium (50 g L-1 cassava starch, 2 g L-1 (NH4)2SO4, 0.6 g L-1 KH2PO4, 0.25 g L-1 MgSO4.7H2O, 0.04 g L-1 ZnSO4.7H2O). Various parameters, pH (5-9), temperature (30-40°C), starch concentration (3-11%), N-sources and concentration (1.5-7.5 g L-1), were studied by using starch medium. Starch concentration was analyzed by idoine solution (Wuttisela et al., 2008). Glucose residue was determined by dinitrosalicylic acid method (Miller, 1959) using glucose as the standard.
Batch and fed-batch culture: Batch fermentation in bioreactor (Biostat B, B Braun) was carried out using 900 mL of production starch medium containing (g L-1): cassava starch 50, urea 7.5, KH2PO4 0.6, MgSO4.7H2O 0.25, ZnSO4.7H2O 0.04, CaCO3 10, the medium was autoclaved at 121°C for 15 min. Rice koji starter 50 g was inoculated in the fermentor and maintained the fermentation process at 40°C with aeration at 0.5 vvm. The agitation effect on lactic acid production was studied between 100-500 rpm. Fed-batch fermentation was studied by adding 400 mL fresh medium containing 5% cassava starch and minerals 10 h after inoculation. The final volume was reached at 1.5 L. The temperature, aeration and agitation were controlled at 40°C, 0.5 vvm and 200 rpm, respectively.
DNA analysis: The ITS region of rDNA was amplified using primers ITS-1F (5'-CTT GGT CAT TTA GAG GAA GTA A-3') and ITS-4B (5'-CAG GAG CTT GTA CAC GGT CCA G-3') (White et al., 1990). The reaction components for the PCR were: 1-20 ng of the total DNA, 0.25 μM of each primer, 2 unit of Taq DNA polymerase, 200 μM of each dATP, dGTP, dCTP, dTTP, 2 mM MgCl2 and 1x PCR buffer. The final volume was adjusted to 25 μL with deionized water. Cycle sequences of ITS region were directly determined using PCR products with those two primers that were mentioned above by ABI PrismTM BigDyeTM Termination v3.1 Cycle Sequence Ready Reaction Kit (Applied Biosystems, Foster, CA) according to the manufacturer’s instruction. The sequences were compared by BLASTn Homology Search . The phylogenetic trees were constructed by ClustalX version 2.1.
RESULTS AND DISCUSSION
Lactic acid selected fungi: Twenty and twenty three of fungal strains were isolated from soils at 30 and 35°C, respectively. However no fungus were screened from the higher temperature at 40 and 45°C. The results of acid and starch hydrolysis testing on starch agar plate (data not shown) revealed that fungal strains from 35°C were produced higher lactic acid than the fungal strain from 30°C. Then 9 fungal strains were selected and the secondary screening in broth medium was investigated. Lactic acid production from 3% starch medium by the selected fungal strains at 35°C were showed in Fig. 1. All of them were Rhizopus sp. The result indicated that starch was quickly hydrolyzed to glucose in the first day (high glucose residue at day 1) and it was utilized by the fungus. Therefore, glucose residue was decreased at day 2 and 3. Glucose was rapidly consumed by some strains such as LTH3, LT2 and LTH1.
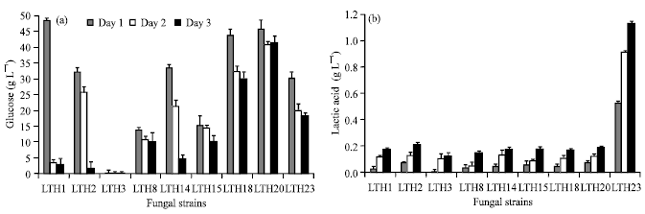 | |
| Fig. 1(a-b): | (a) Time course of glucose residue and (b) Lactic acid production from selected Rhizopus sp. strain in 3% starch medium at 35°C for 3 days |
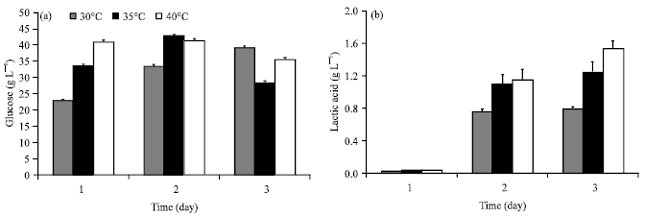 | |
| Fig. 2(a-b): | Temperature effect on lactic acid production by Rhizopus sp. strain LTH23 from 3% starch medium: (a) Glucose residue and (b) Lactic acid |
Except the strain of LTH20, LTH18 and LTH23 glucose in the fermentation broth was remained higher at day 2 and day 3 than the strains of LTH15, LTH8 and LTH14 (Fig. 1a). On the other hand, lactic acid was produced and more accumulated at day 2 and highest at day 3. However the results showed that the highest lactic acid was produced by Rhizopus sp. strain LTH23 (Fig. 1b).
Temperature effect on lactic acid production: Temperature effect on lactic acid production by Rhizopus sp. strain LTH23 is shown in Fig. 2. Starch medium at 3% concentration was hydrolysed and glucose residue was consumed. Remaining glucose was accumulated until it reached its highest at day 3 when incubated at 30°C. While at 35 and 40°C glucose were consumed and highest accumulation at day 2 and declining at day 3 (Fig. 2a). Low lactic acid was produced at day1 at all temperature level and increasing at day 2 and day 3 at all temperature levels (Fig. 2b). Lower lactic acid was produced at 30°C and increased when increasing incubation temperature. However the highest lactic acid was produced at 40°C (Fig. 2b). This result showed that Rhizopus sp. strain LTH23 was thermotolerant strain and the enhancing temperature for higher lactic acid production was at 40°C. Kitpreechavanich et al. (2008) reported that Rhizopus microsporus TISTR 3518 was a thermotolerant fungi that produced L-lactic acid. It was isolated from loog-pang and grew well at 45°C.
pH effect on lactic acid production: Five different initial pH medium were studied for lactic acid production from 3% starch medium. Figure 3 showed the profiled of residual glucose that derived from starch hydrolysis and lactic acid production by Rhizopus sp. strain LTH23 at 3 days fermentation. Higher glucose residues were accumulated at the first day of cultivation and decreased at day 2 and 3 (Fig. 3a). At the same time, lactic acid was produced and increased at day 2 and highest at day 3 of each pH level. Beside this more lactic was increased at higher pH level and highest at pH 9 (Fig. 3b).
Starch concentration effect on lactic acid production: Lactic acid production was studied at 5 levels of initial starch concentration. Due to the glucose residues were the sum of the formation by saccharification and fungal consumption.
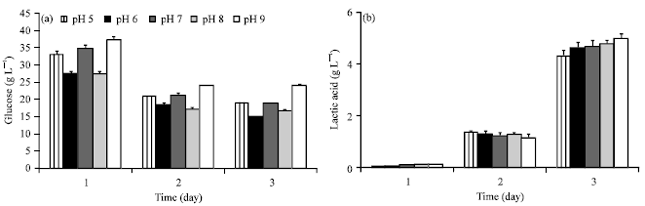 | |
| Fig. 3(a-b): | Effect of pH on lactic acid production by Rhizopus sp. strain LTH23 from 3% starch medium: (a) Glucose residue and (b) Lactic acid |
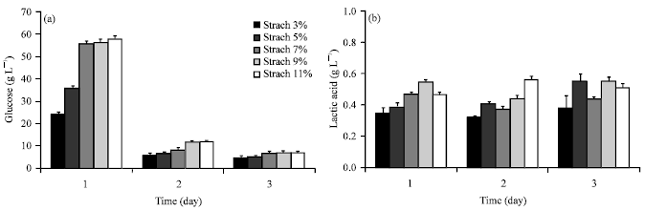 | |
| Fig. 4(a-b): | Starch concentration effect on lactic acid production by Rhizopus sp. strain LTH23 at various starch concentration: (a) Glucose residue and (b) Lactic acid |
It was found that starch was rapidly converted to glucose residue at day 1. Whereas glucose residue at day 2 and 3 at all levels of initial starch concentration were sharply decreased because of no starch remaining in the medium (Fig. 4a). However higher initial starch concentration, higher sugars remaining in the medium was found at 7, 9 and 11% starch medium (day 1) and nearly the same level. Lactic acid production from those of 5 starch concentrations were showed at Fig. 4b. The higher lactic acid was produced at the higher starch concentration in the medium, except at 11% starch (day 1). Due to the starch solution was more viscous at higher starch concentration. Beside this it was found that lactic acid production from 7, 9 and 11% starch solutions were lower at day 2 and 3. Huang et al. (2005) reported that due to substrate inhibition, a low lactic acid was produced at the initial higher starch concentration between 3.75 and 6.28%. In this studied it was showed that 5% starch concentration was suitable for lactic acid production. Because the amount of lactic acid concentration was continuously produced until at day 3. Furthermore no effect of substrate concentration and low viscosity were found at this concentration.
N-sources effect on lactic acid production: Two N-sources, (NH4)2SO4 and urea at various concentration (1.5-7.5g L-1) were added in 5% starch medium. Starch was rapidly converted to glucose by Rhizopus sp. strain LTH23. High glucose residues between 20-35 g L-1 were detected at day 1 in various concentration of both N-sources (Fig. 5a, b). Glucose residues were consumed and low remaining (4-7 g L-1) at day 3. Whereas lactic acid was low produced at the first 2 days and increasing at day 3 (Fig. 5c, d). However the lowest concentration of (NH4)2SO4 at 1.5 g L-1 promoted the highest lactic acid production. In contrast lactic acid was increased at the higher concentration of urea. The highest lactic acid was gained from urea at 7.5 g L-1 (Fig. 5d). While lactic acid was decreased at the higher level of (NH4)2SO4 (Fig. 5c). In contrast the highest lactic acid was detected at the lowest concentration of (NH4)2SO4.
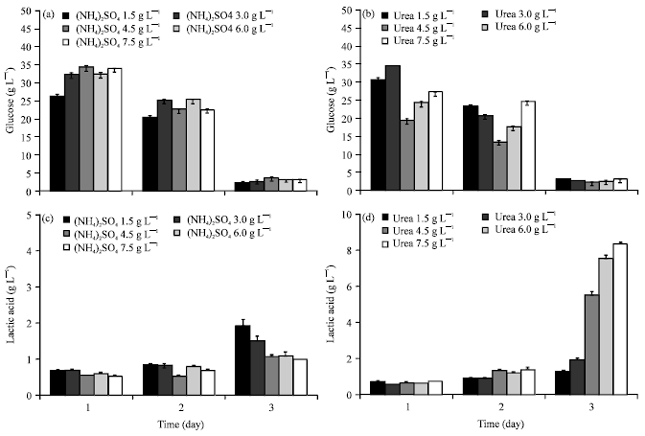 | |
| Fig. 5(a-d): | N-sources effect on lactic acid production by Rhizopus sp. strain LTH23 in 5% starch medium at 40°C: (a) (NH4)2SO4; glucose residue, (b) Urea; glucose residue (c) (NH4)2SO4; lactic acid and (d) Urea; lactic acid |
Nitrogen source promote mycelium growth and also enhanced lactic acid production. The fungal strain of Rhizopus oryzae mostly preferred ammonium sulfate than ammonium nitrate, urea, yeast extract, peptone and corn steep liquor for lactic acid production (Jin et al., 2003; Yin et al., 1997; Zhou et al., 1999). Furthermore, dairy manure can be used as nitrogen source for lactic acid production. Lactic acid concentration was increased from 6.48-57.7 g L-1 by adding crude protein from 0.21-1.68 g L-1, respectively (Yao et al., 2010).
Agitation effect on lactic acid production: Figure 6 showed the effect of agitation speed on lactic acid production in batch process. Starch was quickly hydrolyzed and sharply decreased at agitation speed of 200, 300 and 400 rpm and slowly decreased at 100 and 500 rpm. Finally it was absolutely hydrolyzed at 24 h of fermentation (Fig. 6a). Glucose residue accumulation at various times were performed in Fig. 6b. Corresponding to starch hydrolysis, glucose residues at all agitation speeds were rapidly increased and reached the highest level at 24 h. Glucose level were remained high after that sharply decreased at 60 h of agitation speed 100, 300 and 400 rpm. At 200 rpm, glucose accumulation was constantly high after 20 h and rapidly declined after 78 h. However the glucose residues of all agitation speeds were used by the fungi and low remaining at the end of fermentation process. Effect of agitation speed on lactic acid production was showed in Fig. 6c. It was revealed that at 200 rpm was the optimum speed for lactic acid production by Rhizopus sp. strains LTH23. The maximum lactic acid concentration (20 g L-1) was produced after 60 h cultivation. Bai et al. (2003) studied on the effect of agitation speed and aeration on morphology and lactic acid by R. oryzae R1021. They found that the highest lactic acid production (76.2 g L-1) was gained by agitation speed and aeration rate were 300 rpm and 0.6 vvm, respectively after 36 h cultivation from glucose 100 g L-1. Agitation speed of submerge culture has a variety of effect on microorganisms, including rupture of the cell wall, change in morphology of mycelium, influence on rate of growth and variation of formation product (Darah et al., 2011). In order to improve lactic acid production by Rhizopus sp. strains LTH23, fed-batch process was investigated in the next section.
Lactic acid production in fed batch fermentation: Fed batch process was carried out by adding fresh medium at Lactic acid production by Rhizopus sp. strain LTH23 from 5% starch medium at 40°C, 200 rpm and 0.5 vvm in fed-batch process, adding 400 mL of fresh medium at 10 h cultivation with 700 mL of initial medium volume 10 h of cultivation.
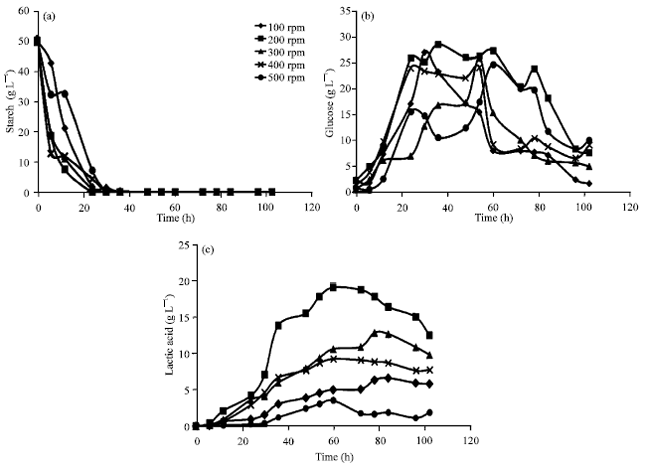 | |
| Fig. 6(a-c): | Effect of agitation speed on lactic acid production by Rhizopus sp. strain LTH23 in 5% (a) Starch medium, (b) Glucose and (c) Lactic acid with 1 L at 40°C, aeration 0.5 vvm |
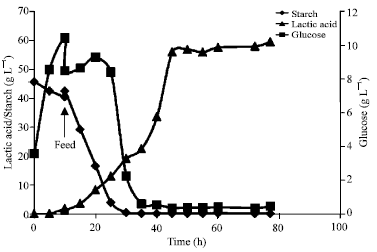 | |
| Fig. 7: | Lactic acid production by Rhizopus sp. strain LTH23 from 5% starch medium at 40°C, 200 rpm and 0.5 vvm in fed-batch process, adding 400 mL of fresh medium at 10 h cultivation with 700 mL of initial medium volume |
The starch, glucose residue and lactic acid profile were shown in Fig. 7. Starch was rapidly hydrolyzed by Rhizopus sp. strains LTH23 after 30 h of cultivation. While high glucose residue was accumulated and rapidly decreased after 35 h of fermentaion. High lactic acid concentration (56.1 g L-1) was reached at 45 h and it was increased to 59.6 g L-1 at 77 h. The corresponding productivity were to 1.25 g L-1 h-1 at 45 h and 0.77 g L-1 h-1 at 77 h. In this process, more lactic acid was produced than batch process 3 times. This result showed that fed-batch process enhanced lactic acid production.
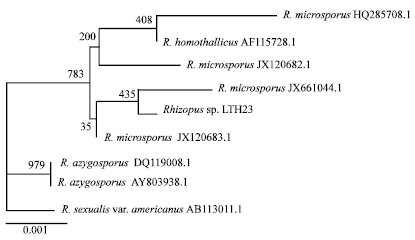 | |
| Fig. 8: | A phylogenetic tree of ITS sequences of Rhizopus sp. LTH23 with Rhizopus strain from GeneBank data base by neighbor joining method using Clustalx version 2.1 |
Because there was no effect of substrate inhibition from glucose accumulation after feeding with fresh medium at 10 h. Due to glucose residue in the fermentation broth was diluted by adding the medium.
Identification of fungal species by ITS sequence: Schipper (1984) divided genus Rhizopus into three groups, R. stolonifer, R. oryzae and R. microsporus by morphology and growth temperatures. Recently, fungal systematic and classification by the Internal Transcribed Spacer (ITS) regions of the ribosomal operon have been used. The phylogenetic relationships of Rhizopus sp., LTH23 was reconstructed using the DNA sequences of ITS region (669 bp) as showed in Fig. 8. The fungal strain was closely related with Rhizopus microsporus JX 661044.1 with 99.85% similarity. Then it was identified as Rhizopus microsporus LTH23.
CONCLUSION
In the present study, mesophilic fungus Rhizopus sp. LTH23 could be screened from soil sample. It is a thermotolerant strain that the highest lactic acid could be produced at 40°C. The medium containing 5% starch was hydrolyzed directly from this strain without pretreatment with enzyme. In order to control the growth pH during the fermentation, neutralizing agent such as calcium carbonate was added into the medium. However, in this study Rhizopus sp., LTH23 can be produced highest lactic acid at pH 9 without adding CaCO3 and supplement with urea 7 g L-1. The influence of agitation speed on lactic acid production between 100-500 rpm were investigated in batch fermentation process. Maximum lactic acid at 20 g L-1 was gained from 200 rpm at 60 h of batch fermentation. In order to enhance production of lactic acid, fed-batch process was conducted. Lactic acid concentration from fed-batch fermentation was 3 times higher than batch process at 60 h of cultivation.
Molecular techniques were used to identify the fungus at species level using Internal Transcribed Spacer region of nuclear rDNA gene. The phylogenetic tree was reconstructed by the sequences from GenBank data base. The fungus was placed in the same clade of Rhizopus microsporus and very closely related with Rhizopus microsporus JX661044.1. Then it was named as Rhizopus microsporus LTH23.
ACKNOWLEDGMENTS
The authors are grateful for the financial support provided by Mahasaraham University. We thank Lisa Vohle, Busala Nonthapa and Chonlada Sangsri for technical assistance.
REFERENCES
- Bai, D.M., M.Z. Jia, X.M. Zhao, R. Ban, F. Shen, X.G. Li and S.M. Xu, 2003. L(+)-Lactic acid production by pellet-form Rhizopus oryzae R1021 in a stirred tank fermentor. Chem. Eng. Sci., 58: 785-791.
CrossRefDirect Link - Benthin, S. and J. Villadsen, 1995. Production of optically pure D-lactate by Lactobacillus bulgaricus and purification by crystallisation and liquid/liquid extraction. Applied Microbiol. Biotechnol., 42: 826-829.
CrossRefDirect Link - Connolly, E. and B. Lonnerdal, 2004. D(-)-Lactic acid-producing bacteria safe to use in infant formulas. Nutrafoods Res., 3: 37-49.
Direct Link - Darah, I., G. Sumathi, K. Jain and S.H. Lim, 2011. Influence of agitation speed on tannase production and morphology of Aspergillus niger FETL FT3 in submerged fermentation. Applied Biochem. Biotechnol., 165: 1682-1690.
CrossRefPubMedDirect Link - Fourest, E., C. Canal and J.C. Roux, 1994. Improvement of heavy metal biosorption by mycelial dead biomasses (Rhizopus arrhizus, Mucor miehei and Penicillium chrysogenum): pH control and cationic activation. FEMS Microbiol. Rev., 14: 325-332.
CrossRefPubMedDirect Link - Ganguly, R., P. Dwivedi and R.P. Singh, 2007. Production of lactic acid with loofa sponge immobilized Rhizopus oryzae RBU2-10. Bioresour. Technol., 98: 1246-1251.
CrossRefDirect Link - Huang, L.P., B. Jin, P. Lant and J. Zhou, 2005. Simultaneous saccharification and fermentation of potato starch wastewater to lactic acid by Rhizopus oryzae and Rhizopus arrhizus. Biochem. Eng. J., 23: 265-276.
Direct Link - Jin, B., L.P. Huang and P. Lant, 2003. Rhizopus arrhizus--A producer for simultaneous saccharification and fermentation of starch waste materials to L(+)-Lactic acid. Biotechnol. Lett., 25: 1983-1987.
PubMed - Kitpreechavanich, V., T. Maneeboon, Y. Kayano and K. Sakai, 2008. Compartive characterization of L-lactic acid -producing thermotolerant Rhizopus fungi. J. Bioresour. Bioeng., 106: 541-546.
CrossRefDirect Link - Kusumaningtyas, E., R. Widiastuti and R. Maryam, 2006. Reduction of aflatoxin B1 in chicken feed by using Saccharomyces cerevisiae, Rhizopus oligosporus and their combination. Mycopathologia, 162: 307-311.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Park, E.Y., P.N. Anh and N. Okuda, 2004. Bioconversion of waste office paper to L(+)-lactic acid by the filamentous fungus Rhizopus oryzae. Bioresour. Technol., 93: 77-83.
CrossRef - Schipper, M.A.A., 1984. A revision of the genus Rhizopus. I. The Rhizopus stolonifer-group and Rhizopus oryzae. Stud. Mycol., 25: 1-19.
Direct Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Yao, W., X. Wu, J. Zhu, B. Sun and C. Miller, 2010. Utilization of protein extract from dairy manure as a nitrogen source by Rhizopus oryzae NRRL-395 for l-lactic acid production. Bioresour. Technol., 101: 4132-4138.
CrossRef - Woiciechowski, A.L., C.R. Soccol, L.P. Ramos and A. Pandey, 1999. Experimental design to enhance the production of L-(+)-lactic acid from steam-exploded wood hydrolysate using Rhizopus oryzae in a mixed-acid fermentation. Process Biochem., 34: 949-955.
CrossRefDirect Link - Yin, P., N. Nishina, Y. Kosakai, K. Yahiro, Y. Pakr and M. Okabe, 1997. Enhanced production of L(+)-Lactic acid from corn starch in a culture of Rhizopus oryzae using an air-lift bioreactor. J. Ferment. Bioeng., 84: 249-253.
CrossRefDirect Link - Yoshihara, K., Y. Shinohara, T. Hirotsu and K. Izumori, 2003. Chitosan productivity enhancement in Rhizopus oryzae YPF-61A by A-psicose. J. Biosci. Bioeng., 95: 293-297.
CrossRefDirect Link - Zhou, Y., J.M. Dominguez, N. Cao, J. Du and G.T. Tsao, 1999. Optimization of L-lactic acid production from glucose by Rhizopus oryzae ATCC 52311. Applied Biochem. Biotechnol., 78: 401-407.
CrossRefDirect Link - Wuttisela, K., S. Shobsngob, W. Triampo and D. Triampo, 2008. Amylose/amylopectin simple determination in acid hydrolyzed tapioca starch. J. Chil. Chem. Soc., 53: 1565-1567.
CrossRefDirect Link