
K.A. Naveen
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Uttar Pradesh, 243 122, India
S.D. Singh
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Uttar Pradesh, 243 122, India
J.M. Kataria
Central Avian Research Institute, Izatnagar, Uttar Pradesh, 243 122, India
R. Barathidasan
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Uttar Pradesh, 243 122, India
K. Dhama
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Uttar Pradesh, 243 122, India
Journal of Biological Sciences
Year: 2014 | Volume: 14 | Issue: 2 | Page No.: 134-141







ABSTRACT
Newcastle Disease (ND) is a highly contagious infection of poultry which manifest in a wide range of severity from subclinical infection to lethal disease. In the past, a number of Newcastle disease outbreaks in poultry and other bird species have been ascribed to pigeon paramyxovirus type-1 (PPMV-1) infection. The conventional in vivo pathogenicity tests to assess the pathogenicity of PPMV-1 viruses have provided equivocal results. Lately, restriction enzyme analysis technique has been employed for unequivocal identification of individual strains of Newcastle Disease Virus (NDV) in poultry. In this study, sequence analysis of the F1/F2 cleavage site of the F gene of APMV-1 isolated from pigeons in India was attempted for pathotype prediction and determination of molecular epidemiology. Six pigeon origin NDV isolated in India between 1991 and 2001 were selected for this study. A portion of NDV F gene including the cleavage site was amplified by Polymerase Chain Reaction (PCR) and sequenced directly. The total number of nucleotide substitution among all six isolates ranged from 6 to 20; whereas, only four amino acid substitutions were observed. Nucleotides at position 304 and 357 were unique to all the pigeon isolates. The cleavage-activation site (380-397) had no nucleotide substitution and all the six pigeon isolates shared the amino acid sequence 112RRQKRF117 as that of velogenic viruses. The results of this molecular characterization study of Indian PPMV-1 isolates would help design better prevention and control measures for this important pathogen.
PDF Abstract XML References Citation
Received: October 16, 2013;
Accepted: October 31, 2013;
Published: January 11, 2014
How to cite this article
K.A. Naveen, S.D. Singh, J.M. Kataria, R. Barathidasan and K. Dhama, 2014. Molecular Characterization and Phylogenetic Analysis of Selected Pigeon Paramyxovirus Type-1 (PPMV-1) Indian Isolates. Journal of Biological Sciences, 14: 134-141.
DOI: 10.3923/jbs.2014.134.141
URL: https://scialert.net/abstract/?doi=jbs.2014.134.141
DOI: 10.3923/jbs.2014.134.141
URL: https://scialert.net/abstract/?doi=jbs.2014.134.141
INTRODUCTION
Newcastle Disease (ND) is a highly contagious infection of poultry that, depending on the virulence of the virus strain, may manifest in a wide range of severity from subclinical infection to lethal disease (Alexander, 2001; Mohanty et al., 2001). It is caused by members of Avian Paramyxovirus (APMV) serotype-1 of genus Avulavirus in the family Paramyxoviridae and has a wide host range infecting about 27 of the 50 order of birds which include over 250 species (Alexander and Parsons, 1984; Miller et al., 2010; Alexander et al., 2012). The course of ND in pigeons (Columba livia) is similar to those seen in chickens (Stewart, 1971; Utterbak and Schwartz, 1973) leading to the earlier assumption that the disease in pigeons was a result of spread from domestic poultry (Pearsons and MaCann, 1975; Cattoli et al., 2011). During 1981 to 1985, infection of racing and show pigeons with NDV became worldwide causing a frequently fatal disease primarily associated with neurological signs. The virus responsible for the panzootic was characterised by pathogenicity tests and monoclonal antibody (mAb) binding studies and found to be a variant form of classical Newcastle Disease Virus (NDV). The variant virus was termed ‘Pigeon paramyxovirus-1 (PPMV -1)’ (Alexander et al., 1984, 1987; Meulemans et al., 1987; Pearson et al., 1987).
The contrast between NDV strains of high and low virulence has been explained to some extent by studies evaluating the molecular basis for pathogenicity. The presence of two pairs of basic amino acids at position 112, 113 and 115, 116 along with phenylanaline at 117, makes the fusion protein of virulent viruses more susceptible to cleavage by ubiquitous proteases, helping them to spread throughout the host. In contrast, the presence of two single basic amino acids at 113 and 116, along with leucine at 117, allows the fusion protein of avirulent NDV isolates to be cleaved only by trypsin-like proteases present mainly in the respiratory and digestive tracts (Toyoda et al., 1987; Collins et al., 1993). Sequencing of PPMV-1 showed isolates with both high and low Intravenous Pathogenicity Index (IVPI) values to have an identical cleavage site moiety of two pairs of basic amino acids between 113 and 116 and phenylanaline at 117 (Collins et al., 1996). Thus the wide variation in the pathogenicity of PPMV-1 in chickens could not be related to the amino acid sequence variation at the F protein cleavage site. The deduced amino acid sequence of the protein cleavage site of F gene between 112 and 117 of PPMV-1 isolates had shown some changes over the years. While initial isolates possessed the motif 112GRQKRF117, PPMV-1 isolated after 1990's have been shown to possess 112RRQKRF117 and more lately 112RRKKRF117 (Oberdorfer and Werner, 1998; Meulemans et al., 2002) denoting an evolution in the sequence of the cleavage site of these viruses. The new sequence 112RRQKRF117 found in majority of the PPMV-1 viruses is identical to that associated with highly virulent NDV though the intracerebral pathogenicity index (ICPI) values of such isolates are highly variable, characteristic of ‘pigeon variant’.
The spread of PPMV-1 infection has been worldwide (Alexander, 1997; Kataria et al., 2006; Alexander et al., 2012; Naveen et al., 2013) and has recently been reported from Indian sub-continent (Senthilkumar et al., 2003; Naveen and Singh, 2004; Naveen et al., 2009). All other outbreaks of ND in Indian pigeons were reportedly caused by classical NDV (Mangat et al., 1988; Singh et al., 1989, 1991; Sulochana and Mathew, 1991; Roy et al., 2000; Kumanan et al., 2005). In this study, sequence analysis of the F1/F2 cleavage site of the F gene of APMV-1 isolated from pigeons in India was attempted for pathotype prediction and determination of molecular epidemiology.
MATERIALS AND METHODS
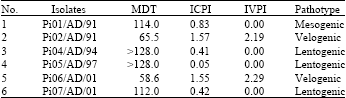
Viruses: Six pigeon origin NDVs isolated in India between 1991 and 2001 were selected for this study (Table 1). The viruses were propagated in 11-day-old embryonated chicken eggs and used for extraction of RNA and generation of ‘F’ gene amplicon for sequencing. The conventional pathogenicity test indices for the PPMV-1 isolates used are given in Table 1.
RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR): Total RNA was isolated from infected Amnio-allantoic Fluid (AAF) using TRIZOL reagent (GIBCO-BRL, USA) as per manufacturer’s protocol. For cDNA synthesis, 100 ng viral RNA and 50 ng of random hexamer primer, in a 20 μL reaction mixture, were heated at 70°C for 5 min and snap-chilled on ice.
| Table 1: | Details of pigeon-origin Newcastle disease virus used and their biological characteristics |
 | |
After adding 4 μL of 5x RT buffer, 0.5 μL RNAse inhibitor, 10 mM dNTPs and 0.5 μL MMLV-RT (Promega, USA) and allowing the random primers to anneal further at 25°C for 10 min, the reaction was carried out at 37°C for 1 h, followed by inactivation of the enzyme at 95°C for 3 min. The PCR primer pair published elsewhere to amplify 356 bp sequence of NDV F gene was used (Nanthakumar et al., 2000a). The primer sequences were as follows: Forward, 5'-GCAGCTGCACGGATTGTGGT-3': (nt 158-177), reverse, 5'-TCTTTGAGCAGGAGGATGTTG-3' (nt 513-493). PCR was carried out in a 50 μL reaction mixture using 0.3 μM each forward and reverse primers, 200 μM dNTPs, 1x PCR buffer with 2 mM MgCl2. 3 μL cDNA and 2.5 U of Taq DNA Polymerase (Promega, USA). After denaturation of the DNA template at 94°C for 2.5 min, amplification was carried out for 35 cycles of 94, 58 and 72°C, each for 45 sec, with final extension at 72°C for 5 min. The PCR amplified products were confirmed by their size in 1.5% agarose gel stained with ethidium bromide (0.5 μg mL-1).
Sequencing and phylogenetic analysis: The amplified F gene products were sequenced at the DNA sequencing facility, Department of Biochemistry, Delhi South Campus University, New Delhi. Comparisons of the sequence data obtained in this study were made with published sequences of ND viruses. The nucleotide and deduced amino acid sequences were aligned separately using the Clustal method of “Edit Seq” programme of the Lasergene software (DNASTAR Inc., USA). The aligned nucleotide sequences were subjected to phylogenetic analysis using ‘Meg Align’ programme of Lasergene software.
RESULTS
A 356 bp PCR product of NDV F gene was obtained using RT-PCR which was sequenced and aligned. All sequences were truncated to 283 nucleotides falling between nt positions 230 and 512 as per Toyoda et al. (1987). The aligned nucleotide and deduced amino acid sequences are shown in Fig. 1 and 2, respectively. The total number of nucleotide substitution among all six isolates ranged from 6 (Pi04/AD/94 and Pi07/AD/01) to 20 (Pi02/AD/91) whereas, only four amino acid substitutions were observed. Most of the nucleotide changes were noticed at the third nucleotide of a codon. Nucleotide substitution at position 424 in Pi01/AD/91 and 268 in Pi02/AD/91 were found to be unique.
 | |
| Fig. 1: | Aligned NDV ‘F’ gene nucleotide sequence of selected PPMV-1 isolates. All sequences were truncated to 283 nucleotides falling between nt positions 230 and 512. Variations are identified with checkboxes |
Four nucleotide substitution at position 289, 340, 469 and 490 were found in isolates Pi01/AD/91, Pi05/AD/97 and Pi07/AD/01 while there were 19 similar nucleotide substitution among isolates Pi02/AD/91 and Pi06/AD/01, of which only three at position 373, 407 and 416 resulted in changes of amino acids 108 T to A, 121 I to V and 124 G to S, respectively. Out of a total of 6 to 10 changes at nucleotide levels among isolates Pi01/AD/91, Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01, there was only one change in amino acid 132 A to S. Nucleotides at position 304 and 357 were unique to all the pigeon isolates. The cleavage-activation site (380-397) had no nucleotide substitution and all the six pigeon isolates shared the amino acid sequence 112RRQKRF117.
The divergence among the six pigeon isolates ranged from 0% (between Pi07/AD/01 and Pi04/AD/94) to 12.2% (between Pi02/AD/91and Pi05/AD/97) at nucleotide level. He divergence among the isolates at amino acid level ranged from 0 to 4.5%.
 | |
| Fig. 2: | Aligned fusion protein amino acid sequence (deduced) of PPMV-1 isolates. Variations are identified with checkboxes |
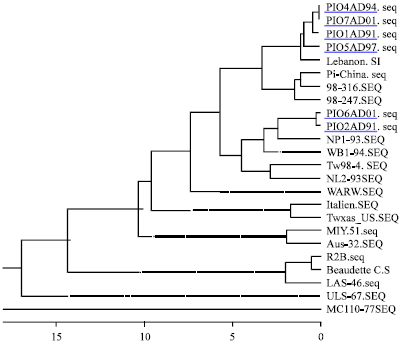 | |
| Fig. 3: | Phylogenetic tree based on the fusion protein’s partial amino acid sequence of Indian PPMV-1 isolates (underlined) and published sequences |
The pigeon isolates showed divergence of 10.6-14.4% from LaSota (vaccine strain) and 5.7-8% from R2B (vaccine strain) at amino acid level. The Indian pigeon isolates had divergence from a Chinese pigeon isolate (PiChina) ranging from 7.1-13.9% at nucleotide level and 3.2-5.5% at amino acid levels. Phylogenetic tree based on nucleotide prepared by “Meg Align” programme is presented in Fig. 3. The six pigeon isolates formed two major lineages. Isolates Pi01/AD/91, Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01 were placed along with velogenic NDV strains Lebanon 70 and formed a close cluster with pigeon isolates of China and Belgium. Isolates Pi02/AD/91 and Pi06/AD/01 were placed in a cluster with virulent NDV isolates from India, Nepal, Taiwan and the Netherlands. The Taiwanese strain TW98-4 is a representative of group VII lineage. Strain WARW which is a representative of lineage VI formed a distinct branch.
DISCUSSION
Pathotyping of PPMV-1 isolates by conventional velogenic, mesogenic and lentogenic criteria have been more difficult than with most other NDV isolates. Virulent and non-virulent NDV strains can be differentiated by polymerase chain reaction (PCR) (Kant et al., 1997). Sequence analysis of the F gene encoding the fusion protein cleavage site has been exploited by many workers for seeking an explanation for the unusual pathogenecity of PPMV-1 (Collins et al., 1994, 1996; King, 1996; Kommers et al., 2001). Among the six pigeon origin NDV selected for this study, isolates Pi02/AD/91 and Pi06/AD/01 had velogenic parameters, isolates Pi01/AD/91 had mesogenic parameters and isolates Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01 had lentogenic parameters under conventional pathogenicity tests. They lacked close nucleotide homology but were clearly divided into two groups on the basis of amino acid sequence. Isolate Pi02/AD/91 and Pi06/AD/01 had 100% homology at amino acid level while isolates Pi01/AD/91, Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01 shared complete amino acid homology. All Indian pigeon isolates possessed G at position 104 and S at 107 which is common to pigeon isolates Pichina, 1168, 760; Virulent NDV’s NP-1, WB1 and UP1. This is a common feature of all Taiwanese isolates placed in group VIII (Yang et al., 1999). However, all Indian pigeon NDV isolates originated between 1998 and 2002 were found to have E at 104 and four had T at 107 (Kumanan et al., 2005) suggesting evolution of PPMVs. Substitution of I_V at 121 and S at 124 found in Pi02/AD/91 and Pi06/AD/01 is characteristic of velogenic ND viruses though the substitution of T_A at 108 is unique to the two pigeon isolates as it has not been reported for any NDV till date. The S for A at 132 in Pi01/AD/91, Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01 is also reported for isolates PiChina, 1168/84, 730/83, PiTA and appears to be a unique feature among pigeon variants.
In keeping with the results obtained in other studies, all pigeon origin NDV examined in this study had fusion protein cleavage site sequence that would place them in the virulent pathotype (112RRQKRF117). Meulemans et al. (2002) have reported that most PPMV-1 isolated after 1990's possessed this motif. They further reported that the isolates with motif 112RRQKRF117 had highly variable but largely lower ICPI values (mean 0.69) than those reported for PPMV-1 viruses displaying the motif 112GRQKRF117 (mean 1.44). Among all isolates in the present study with motif 112RRQKRF117, four had fairly low ICPI values. The velogenic pathotype of isolate Pi02/AD/91 and Pi06/AD/01 earlier assessed by ICPI and IVPI can be confirmed by the presence of two pairs of basic amino acid at the fusion protein cleavage sequence and amino acid S at 124 in the N terminal region of the F1protein, reported in reference velogenic strain (McGinnes and Morrison, 1986; Toyoda et al., 1987; Seal et al., 1995; Collins et al., 1996). Both the isolates, however, differed from other velogenic NDV by possessing A at 108. Isolates Pi01/AD/91, Pi04/AD/94, Pi05/AD/97 and Pi07/AD/01 were found to be the pigeon variant by having F protein cleavage sequence and amino acid substitute at 104, 107 and 132 similar to those observed in all pigeon PMV-1. The Indian pigeon isolates were distinguished by the presence of G at 124 indicating them to be intermediate in their evolution. The fusion inducing hydrophobic amino acid stretch at N-terminus of F1 protein (117-142 aa residues), lysine at position 76 important in disulphide bond formation between F1 and F2 and amino acids at potential glycosylation site (85-NRT-87) were found to be conserved in all the pigeon isolates, as reported by earlier workers (Toyoda et al., 1987; Seal et al., 1995; Nanthakumar et al., 2000b).
On the basis of phylogenetic analysis, isolates Pi02/AD/91 and Pi06/AD/01 were found to be closely related. These isolates were placed in group VIIa of which NL-2-93 and TW98-4 are the members. Yang et al. (1999) had proposed that genotype VII viruses might have caused a fourth panzootic of ND which is distinct from the third panzootic caused by PPMV-1. Pi02/AD/91 and Pi06/AD/01 seems to be the first pigeon isolates which has been placed in this group. They were phylogenetically related to velogenic chicken isolates of Indian sub-continent NP, WB and UP suggesting an adaptation of chicken NDV in pigeons. The substitution of A for T at 108 seems to be one of the changes needed for adaptation to pigeons. The other four isolates showed relatedness to velogenic strain Lebanon-70 and PPMV-1 isolates of Belgium and China. Seeing the emergence and prevalence of PPMV-1 in India (Senthilkumar et al., 2003; Naveen and Singh, 2004; Kataria et al., 2006; Naveen et al., 2009, 2013), authors suggested that advanced and rapid diagnostics (Deb et al., 2013; Dhama et al., 2011, 2013a, b), effective vaccines (Kataria et al., 2004; Dhama et al., 2008, 2013c), emerging therapeutics (Mahima et al., 2012; Dhama et al., 2013d, e; Tiwari et al., 2013) and suitable prevention and control measures (Kataria et al., 2005; Dhama et al., 2013f; Tiwari et al., 2013) need to be adapted to tackle this important pathogen and safeguard health of pigeons which would help in alleviating economic losses and sufferings to pet/pigeon owners.
CONCLUSION
In conclusion, the present study reiterates the previous findings that conventional pathogenicity tests are not conclusive markers to identify pathotype of PPMV isolates. Also, F gene cleavage site amino acid motif analysis does not help in classification of PPMV-1 isolates and pathotyping. There is every chance to overlook potential virulence and molecular characterisation should be carried out regularly to keep track of evolution of ND viruses. Moreover, molecular basis of pathogenicity as claimed is not 100% conclusive at least for PPMV isolates. Alternate strategies for detection and differentiation as well as appropriate prevention and control strategies for PPMV-1 viruses are to be adopted for safeguarding health of pigeons.
REFERENCES
- Alexander, D.J., E.W. Aldous and C.M. Fuller, 2012. The long view: A selective review of 40 years of Newcastle disease research. Avian Pathol., 41: 329-335.
CrossRefPubMedDirect Link - Alexander, D.J. and G. Parsons, 1984. Avian paramyxovirus type 1 infections of racing pigeons: 2. Pathogenicity experiments in pigeons and chickens. Vet. Rec., 114: 466-469.
CrossRefDirect Link - Alexander, D.J., P.H. Russell and M.S. Collins, 1984. Paramyxoviru stype 1 infections of racing pigeon: Characterization of isolated viruses. Vet. Record, 114: 444-446.
PubMedDirect Link - Alexander, D.J., R.J. Manvell, P.A. Kemp, G. Parsons and M.S. Collins et al., 1987. Use of monoclonal antibodies in the characterisation of avian paramyxovirus type 1 (Newcastle disease virus) isolates submitted to an international reference laboratory. Avian Pathol., 16: 553-565.
CrossRefPubMedDirect Link - Cattoli, G., L. Susta, C. Terregino and C. Brown, 2011. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Invest., 23: 637-656.
CrossRefPubMedDirect Link - Collins, M.S., J.B. Bashiruddin and D.J. Alexander, 1993. Deduced amino acid sequences at the fusion protein cleavage site of Newcastle disease viruses showing variation in antigenicity and pahtogenicity. Arch. Virol., 128: 363-370.
CrossRefDirect Link - Collins, M.S., I. Strong and D.J. Alexander, 1994. Evaluation of the molecular basis of pathogenicity of variant Newcastle disease viruses termed pigeon PMV-1 viruses. Arch. Virol., 134: 403-411.
PubMed - Collins, M.S., I. Strong and D.J. Alexander, 1996. Pathogenicity and phylogenetic evaluation of the variant Newcastle disease viruses termed pigeon PMV-1 viruses based on the nucleotide sequence of the fusion protein gene. Arch. Virol., 141: 635-647.
CrossRefPubMedDirect Link - Deb, R., S. Chakraborty, B. Veeregowda, A.K. Verma, R. Tiwari and K. Dhama, 2013. Monoclonal antibody and its use in the diagnosis of livestock diseases. Adv. Biosci. Biotechnol., 4: 50-62.
CrossRefDirect Link - Dhama, K., M. Mahendran, P.K. Gupta and A. Rai, 2008. DNA vaccines and their applications in veterinary practice: Current perspectives. Vet. Res. Commun., 32: 341-356.
CrossRefDirect Link - Dhama, K., A.K. Verma, R. Tiwari, S. Chakraborty and K. Vora et al., 2013. A perspective on applications of Geographical Information System (GIS): An advanced tracking tool for disease surveillance and monitoring in veterinary epidemiology. Adv. Anim. Vet. Sci., 1: 14-24.
Direct Link - Dhama, K., K. Karthik, S. Chakraborty, R. Tiwari, S. Kapoor, A. Kumar and P. Thomas, 2014. Loop-mediated isothermal amplification of DNA (LAMP): A new diagnostic tool lights the world of diagnosis of animal and human pathogens: A review. Pak. J. Biol. Sci., 17: 151-166.
CrossRefDirect Link - Dhama, K., M.Y. Wani, R. Deb, K. Karthik and R. Tiwari et al., 2013. Plant based oral vaccines for human and animal pathogens-a new era of prophylaxis: Current and future prospective. J. Exp. Biol. Agric. Sci., Vol. 1.
Direct Link - Dhama, K., S. Chakraborty, Mahima, M.Y. Wani and A.K. Verma et al., 2013. Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Biol. Sci., 16: 101-111.
CrossRefDirect Link - Dhama, K., S. Chakraborty, M.Y. Wani, R. Tiwari and R. Barathidasan, 2013. Cytokine therapy for combating animal and human diseases-A review. Res. Opin. Anim. Vet. Sci., 3: 195-208.
Direct Link - Dhama, K., S. Chakraborty, S. Kapoor, R. Tiwari and A. Kumar et al., 2013. One world, one health-veterinary perspectives. Adv. Anim. Vet. Sci., 1: 5-13.
Direct Link - Kataria, J.M., C.M. Mohan, S. Dey, B.B. Dash and K. Dhama, 2005. Diagnosis and immunoprophylaxis of economically important poultry diseases: A review. Indian J. Anim. Sci., 75: 555-567.
Direct Link - Kant, A., G. Koch, D. Van Roozelaar, F. Balk and A.T. Huurne, 1997. Differentiation of virulent and non-virulent strains of Newcastle disease virus within 24 h by polymerase chain reaction. Avian Pathol., 26: 837-849.
PubMed - King, D.J., 1996. Avian paramyxovirus type 1 from pigeon: Isolate characterization and pathogenicity after chicken or embryo passage of selected isolates. Avian Dis., 40: 707-714.
Direct Link - Kommers, G.D., D.J. King, B.S. Seal and C.C. Brown, 2001. Virulence of pigeon-origin Newcastle disease virus isolates for domestic chickens. Avian Dis., 45: 906-921.
Direct Link - Kumanan, K., B. Mathivanan, K. Vijayarani, A. Arul Gandhi, K. Ramadass and K. Nachimuthu, 2005. Biological and molecular characterization of Indian isolates of Newcastle disease virus from pigeons. Acta Virol., 49: 105-109.
PubMed - Mahima, A. Rahal, R. Deb, S.K. Latheef and H.A. Samad et al., 2012. Immunomodulatory and therapeutic potentials of herbal, traditional/indigenous and ethnoveterinary medicines. Pak. J. Biol. Sci., 15: 754-774.
CrossRefDirect Link - Mangat, A.P., G. Singh and B.S. Gill, 1988. An outbreak of paramyxovirus encephalomyelitis in racing pigeons in India. Vet. Record, Vol. 123.
CrossRef - McGinnes, L.W. and T.G. Morrison, 1986. Nucleotide sequence of the gene encoding the Newcastle disease virus fusion protein and comparisons of paramyxovirus fusion protein sequences. Virus Res., 5: 343-356.
PubMed - Meulemans, G., M. Gonze, M.C. Carlier, P. Petit, A. Burny and L. Long, 1987. Evaluation of the use of monoclonal antibodies to haemagglutinin and fusion glycoproteins of Newcastle disease virus for virus identification and strain differentiation purposes. Arch. Virol., 92: 55-62.
PubMed - Meulemans, G., T.P. van den Berg, M. Decaesstecker and M. Boschmans, 2002. Evolution of pigeon newcastle disease virus strains. Avian Pathol., 31: 515-519.
CrossRefPubMedDirect Link - Miller, P.J., E.L. Decanini and C.L. Afonso, 2010. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet Evol., 10: 26-35.
CrossRefDirect Link - Mohanty, S.K., J.M. Kataria, S. Mishra, K. Dhama, B.B. Dash and K.C. Verma, 2001. Biological and immunological characterization of a Newcastle disease virus isolated from guinea fowl. Indian J. Comp. Microbiol. Immunol. Infect. Dis., 22: 109-112.
Direct Link - Nanthakumar, T., R.S. Kataria, A.K. Tiwari, G. Buchaiah and J.M. Kataria, 2000. Pathotyping of Newcastle disease viruses by RT-PCR and restriction enzyme analysis. Vet. Res. Commun., 24: 275-286.
CrossRefPubMedDirect Link - Naveen, K.A. and S.D. Singh, 2004. Studies on characterization and pathogenicity of Newcastle disease virus isolated from pigeons in India. Indian J. Vet. Pathol., 28: 150-150.
Direct Link - Naveen, K.A., S.D. Singh and T.R. Mohanty, 2009. Pathobiological and molecular characterization of an Indian isolate of pigeon Paramyxovirus type-I. Indian J. Vet. Pathol., 33: 58-61.
Direct Link - Naveen, K.A., S.D. Singh, J.M. Kataria, R. Barathidasan and K. Dhama, 2013. Detection and differentiation of pigeon paramyxovirus serotype-1 (PPMV-1) isolates by RT-PCR and restriction enzyme analysis. Trop. Anim. Health Prod., 45: 1231-1236.
CrossRefPubMedDirect Link - Oberdorfer, A. and O. Werner, 1998. Newcastle disease virus: Detection and characterization differing in pathogenicity. Avian Pathol., 27: 237-243.
PubMedDirect Link - Pearson, G.L. and M.K. McCann, 1975. The role of indigenous, wild, semidomestic and exotic birds in the epizootiology of velogenic viscerotropic newcastle disease in Southern California 1972-1973. J. Am. Vet. Med. Associat., 167: 610-614.
PubMed - Pearson, J.E., D.A. Senne, D.J. Alexander, W.D. Taylor, L.A. Peterson and P.H. Russel, 1987. Characterization of newcastle disease virus (Avian paramyxovirus-1) isolated from pigeons. Avian Dis., 31: 105-111.
PubMedDirect Link - Roy, P., A.T. Venugopal and A. Koteeswaran, 2000. Antigenically unusual Newcastle disease virus from racing pigeons in India. Trop. Anim. Health Prod., 32: 183-188.
PubMed - Seal, B.S., D.J. King and J.D. Bennett, 1995. Characterization of Newcastle disease virus isolates by reverse transcription PCR coupled to direct nucleotide sequencing and development of sequence database for pathotype prediction and molecular epidemiological analysis. J. Clin. Microbiol., 33: 2624-2630.
Direct Link - Senthilkumar, N., J.M. Kataria, K. Dhama and R. Toroghi, 2003. Sequence analysis of the cleavage site-encoding region of the fusion protein gene of newcastle disease viruses obtained from pigeon, guinea fowl and quail. Indian J. Comp. Microbiol. Immunil. Infect. Dis., 24: 191-193.
Direct Link - Stewart, G.H., 1971. Naturally occurring clinical Newcastle disease in racing pigeons (Columbia livia). Vet. Record, 89: 225-226.
PubMed - Tiwari, R., S. Chakraborty, K. Dhama, M.Y. Wani, A. Kumar and S. Kapoor, 2013. Wonder world of phages: Potential biocontrol agents safeguarding biosphere and health of animals and humans-current scenario and perspectives. Pak. J. Biol. Sci., (In Press).
Direct Link - Utterbak, W.W. and J.H. Schwartz, 1973. Epizootiology of velogenic viscerotropic Newcastle disease in Southern California, 1971-1973. J. Am. Vet. Med. Associat., 163: 1080-1088.
PubMed - Yang, C.Y., H.K. Shieh, Y.L. Lin and P.C. Chang, 1999. Newcastle disease virus isolated from recent outbreaks in Taiwan phylogenetically related to viruses (genotypes VII) from recent outbreaks in Western Europe. Avian Dis., 43: 125-130.
Direct Link