
S. Jagannathan
Tissue Culture Anti Rabies Vaccine Section, Pasteur Institute of India, Coonoor, The Nilgiris, Tamilnadu, India
P. Rahul Gandhi
Department of Biotechnology, Jamal Mohammed College, Trichirapalli, Tamilnadu, India
R. Vijayakumar
Department of Biochemistry, Karpagam Arts and Science College, Coimbatore, Tamilnadu, India
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 6 | Page No.: 521-527







ABSTRACT
Rabies is an acute disease of vertebrate animals which may affect all animals from amphibian to man and it is a single stranded negative sense RNA virus belonging to the genus Lyssavirus of the family Rhabdoviridae. The production of vaccines, diagnosis and further research will therefore continue in this field because a cure for rabies would be a big boon for the developing countries. Virus inactivation is essential for the preparation of vaccines, diagnostic reagent and research purposes. The beta propiolactone (βPL) is widely used as an inactivatent; βPL is very considerable value in the production of biological for virus inactivation and perhaps for DNA inactivation. The inactivation results indicate that the rate of inactivation is directly proportional to the concentration of βPL used. Assessment of the residual βPL present in the vaccine is a mandatory test to vaccine. Vaccine manufacturing also assess the efficacy of inactivation procedure by various in process quality control tests such as tissue culture virus amplification test and inactivation kinetics. This experiments are based on the changing the parameters as well as the various concentration of βPL in the clarified, concentrated high titer of PV11 rabies strain, subsequently that are further purified and formulated with suitable additives and stabilizers. The immunogenicity of the vaccines is analyzed. During the βPL inactivation kinetics gives an idea of the predictable time of inactivation and also the order of inactivation, validation of the inactivation process is an essential part of quality assurance. In the immunobiological industry the time is a crucial factor because to get more inactivated viral harvest yield in a short period and they are processed to further purification and downstream processing. This findings is to reduce the time frame and the residual βPL in various level influenced the tissue culture derived anti rabies vaccine.
PDF Abstract XML References Citation
Received: March 14, 2013;
Accepted: April 02, 2013;
Published: July 12, 2013
How to cite this article
S. Jagannathan, P. Rahul Gandhi and R. Vijayakumar, 2013. Kinetics Analysis of Beta-propiolactone with Tangential Flow Filtration (TFF) Concentrated Vero Cell Derived Rabies Viral Protein. Journal of Biological Sciences, 13: 521-527.
DOI: 10.3923/jbs.2013.521.527
URL: https://scialert.net/abstract/?doi=jbs.2013.521.527
DOI: 10.3923/jbs.2013.521.527
URL: https://scialert.net/abstract/?doi=jbs.2013.521.527
INTRODUCTION
Rabies is a worldwide fatal disease and represents a severe public health problem in developing countries. The rabies is considered to be a reemerging zoonoses in many parts of the world; among particularly in Asia, Africa and Latin America where the disease is enzootic despite the availability of proven and control tools. India is highly endemic country for rabies and tops the list regarding contribution of human deaths in the globe (Sudarshan et al., 2007). The rabies virus is a single stranded negative sense RNA virus belonging to the genus Lyssavirus of the family Rhabdoviridae. The virus causes fatal encephalitis both in humans and animals which still a major public health problem in developing countries in Asia, Africa and Latin America (Trabelsi et al., 2005). Ten million post-exposure treatments are reported among humans (Martinzn, 2000). Production of vero cell based rabies viral vaccine in Roller poly ethylene terephthalate bottles (Jagannathan et al., 2009) and higher vero cell density was achieved in the Fibracel disks verses Cytodex-1 in spinner flask culture for the production rabies virus (Hassanzadeh et al., 2011) showed highly encouraging results. The production of vaccines, diagnosis and further research will therefore continue in this field. Inactivation of this highly neurotrophic virus is essential for the preparation of vaccines, diagnostic reagent and research purposes. In addition to rendering the virus noninfectious, the inactivation method should have the least adverse effect on the vaccine's immunogenicity. Furthermore, the inactivating method should not introduce a toxic substance into the vaccine, in the form of either residual reagent or altered vaccine components which may be toxic to the recipient (Wiktor et al., 1972).
βPL is widely used as an inactivatent; It is very considerable value in the production of biological for virus inactivation and perhaps for DNA inactivation. It has several advantages, It does not require a neutralizing agent to stop its reaction since it is completely hydrolyzed after 3 h at 37°C (Murata and Kanegawa, 1973; LoGrippo, 1960). It is still more quickly degraded in the presence of cellular debris and cell culture media, Its degradation products are not toxic. Vaccine manufacturing also assess the efficacy of inactivation procedure by various in process control tests such as tissue culture amplification test and inactivation kinetics. Inactivation kinetics gives an idea of the predictable time of inactivation and also the order of inactivation. There are many indeterminate factors which might extend the longevity of inactivation kinetics like aggregation, adsorption to walls of the vessel, or presence in aerosol droplets above the surface of the liquid. One point of difference is the initial reactant concentration (Hiatt, 1964). The effective virus inactivation should achieve without any irreversible loss of viral infectivity and with minimal loss of antigenic properties (Budowsky and Zalesskaya, 1991). From the study of Sheng-Fowler et al. (2009) the levels of clearance of HIV retroviral cell substrate DNA that can be achieved with either benzonase digestion of DNA or chemical inactivation by beta propiolactone. Furthermore, the inactivating method should not introduce a toxic substance into the vaccine, in the form of either residual reagent or altered vaccine components which may be toxic to the recipient.
Validation of the viral inactivation process is an crucial part of quality assurance in the immunobiological production industry and time is also a crucial factor because to get more inactivated viral harvest or yield in a short period and which it has to subjected to further purification and down stream processing, this findings make possible to diminish the time frame and to estimate the residual βPL in various levels in the tissue culture derived anti rabies vaccine. The inactivation process plays an important role in the viral vaccine preparation. The βPL and formaldehyde are important inactivators that are commonly used in biological preparations. Though βPL has more merits when compared with formaldehyde (Perrin and Morgeaux, 1995). In the rabies viral vaccine production, βPL has been used for inactivation and during the inactivation process, for the presence of residual βPL may be decreased as the time progress. The present study describes validation of the inactivation strategies, usage of βPL at in different concentrations, inactivation time and temperature.
MATERIALS AND METHODS
Cells: The Vero cell line was procured from the American type culture collection (ATCC) at the 113th passage. At 125th passage level Vero cells were received as monolayer and as frozen cells, from ATCC. Master cell seeds were prepared at 130th passage level and the working cell banks were established at 136th passage level. Quality control tests were performed as per requirement and found complying.
Revival of cells: Stored vials containing Vero cells were taken from liquid nitrogen and immediately placed in a sterile beaker containing sterile distilled water (DH2O) 37°C for rapid thawing. Cells were transferred quickly into two roux bottles containing growth medium (100 mL each with 10% serum). The cells were incubated at 37°C to form monolayer. To remove toxic DMSO and dead cells, a medium change was done after 12-15 h.
Media used for cell culture: Eagle’s Minimum Essential Medium (MEM) with non-essential amino acids, phenol red and 1.5 g L-1 of sodium bicarbonate, pH 7.0 to 7.5 (Himedia, Bombay) was used for Vero cell culture. The dry powder was dissolved in water for injection (WFI) to 1X concentration. For maintenance of Vero cells, the MEM with 2.2 g L-1 of Sodium bicarbonate instead of 1.5 g L-1 pH with a range of 7.4 to 7.6 was used. For pH adjustment CO2 was used. Required quantities of Neomycin and New Born Calf Serum (NBCS) were added at the time of batch culture.
β-propiolactone: β-propiolactone is a colorless stable liquid with a slightly sweetish odour in its undiluted state but unstable in aqueous solution. It is soluble in water and miscible with ethanol, acetone, diethyl ether and chloroform. β propiolactone (FERAK Berlin GmbH; NMR 98.5%) is a highly reactive chemical because of the moderate fire hazard when exposed to heat or flame. The βPL has very high chemical reactivity due to the presence of a strained four membered lactone ring and it is available in a grade containing 97% minimum active ingredient (Jones and Feldman, 1981).
Virus: Fixed strain of Pasteur virus (PV-11) obtained from Institute Pasteur France was initially adopted to primary cultures of fetal bovine kidney cells and later adopted to Vero cells. The mother seed (PV-RAGE-01) was stored in liquid nitrogen. Master seeds were prepared from the mother seed on Vero cells by harvesting the viral fluid, stabilized with 10% New Born Calf Serum (NBCS) and distributed in 5 mL quantity into 50 mL vial and stored at -80°C/Liquid nitrogen. Working virus seed stocks were prepared from master seed as described above and tests for sterility, titre and adventitious agent were carried out.
Propagation of vero cells: Growth medium from the bottles showing confluent monolayer were removed and washed with warm PBS to remove dead cells, serum component and to enhance the trypsin activity. Small quantities of 0.25% trypsin in Dulbecco’s saline A with sodium bicarbonate was added to each bottle and incubated at 37°C for few minutes for the detachment of cells. Based on cell count, cells were dispersed in growth medium and transferred to roux/roller bottles, incubated at 37°C for 48 h (Jagannathan et al., 2009).
Dynamics of virus replication: Roller bottles were examined under microscope, washed with PBS, infected with virus seed material having a virus titre of 10-5 (10 mL per roller bottle) and incubated at 35°C about 30 min for adsorption of the virus. Then about 250 mL of maintenance medium was added. Samples of the maintenance medium were taken daily for virus titration. For maximum yield of virus from the cell culture, the exact multiplication of infection (MOI) relating to the infective virus material and the number of viable cells in the culture were analyzed and used. 5% of the bottles were kept as control culture bottles. For the detection of extreaneous viral agents by heamadsorption test, karyological study.
Titration of Virus infectivity: The virus infectivity in mice (LD50) was done as per the technique described by Koprowski (1973). Groups of 10 mice were inoculated intra cerebrally with 10 fold dilution of the viral harvest, each mouse receiving 0.03 mL. The mice were observed for 14 days and the number that died from rabies was recorded after the first 5 days from inoculation. Any mice showing signs of fixed-virus rabies infection (paralysis, convulsions) between 5th day and 14th day were also included and the titer was calculated by the method of Reed and Muench (1938).
Viral harvests: The supernatant fluids containing virus material were harvested into large sterile bottles and the same bottles containing infected cells used for another 4-5 harvests by replenishing with maintenance medium at an interval of 72 h. The rabies virus comes out in the fluid by budding out process and it does not produce any cytopathic effect (CPE) in the infected cells.
Concentration and purification of SVH: Based on titer value, viral harvests were pooled and sterile filtered using prefilter 0.45 μm membranes and collected in 100 L stainless steel pressure vessels, as Single Viral Harvest (SVH) the SVH were ensured for sterility further subjected to concentrated by the technique namely Tangential Flow ultra Filtration (TFF) using the TFF system (Millipore JMBM 0137) with A molecular weight cut off 100, 000 polysulfone membrane (BIOMAX) was used for concentration. The SVH was concentrated to 20X level by volume. Both retentate and filtrate materials were subjected to mouse titration test to check the integrity of the system and the run. After the concentration the material is Diafiltered with required volume of Phosphate Buffer Saline (PBS) pH 7.2 and ensured the residual BSA. The concentrated, inactivated rabies viral harvests are purified by chromatographic technique.
Virus inactivation kinetics experiments: Concentrated virus stored at +4°C was dispensed in to 25 mL flasks. The purity, concentration of the βPL were rechecked before has been started the inactivation, the concentration was analyzed by the method of Tyler and Beesing (1952). The following conditions are followed for all experiments, before addition of βPL stock solution of 1:10 was prepared, double addition of βPL. First half of addition the volume of βPL, incubated at +22°C for 2 h followed by 2nd addition of the remaining volume of βPL incubated at the experimental temperatures (+37, +22 and +4°C). After addition of all βPL all the samples are under shaking up to 48 h for enhancing the inactivation.
The βPL were added as a two additions for obtain a final concentration of 1:1000, 1:2000 and 1:3000. These suspension are incubated at +37, +22 and +4°C with continuous stirring for 2 h. After 2 h all the samples are kept in +4°C. Samples were collected from each flask 2,24,48,96 and 168 h after addition of βPL. Prior to start the experiments the control βPL are hydrolyzed in the water by the inactivation ratio likes 1:1000, 1:2000 and 1:3000, the inactivation temperature +37°C, with the duration time of 2 h.
Avirulance test: The test was performed with undiluted inactivated sample. The 0.03 mL of sample was inoculated intracerebrally into a group of 10 mice weighing 12-15 g. Those four week old Swiss albino mice were procured from the Laboratory Animal facility department of Pasteur Institute of India, Coonoor. The animals were housed and looked after as per the regulations and permission to conduct the study was obtained from the Institutional animal ethics committee for experimentation of animals The Mice were observed for 14 days. Any deaths occurring during the first 4 days were disregarded. Simultaneously those samples are inoculated into the mice for conformation of the virulence factor.
Experimental vaccine formulation: The inactivated and purified rabies antigens were used for the formulation of inactivated rabies vaccine. Totally six formulation of inactivated tissue culture rabies vaccine (VARV A/08, B/08,C/08,D/08,E/08,F/08 and G/08) was taken. Vaccines were formulated using antigens with virus titer of log 10-5.1 before inactivation. human albumin and maltose were added as additives to a final concentration of 1 and 5%, respectively in the single human dose.
Immunization: Two sets of Guinea pigs each containing 10 numbers was immunized with three doses of each 1.0 mL of each of A/08,B/08,C/08,D/08,E/08,F/08 and G/08 formulations on days 0, 3 and 7 by intramuscular inoculation respectively. The serum samples from immunized guinea pigs were collected on the 90th day after the primary inoculation and titrated for the estimation of rabies specific antibodies by Rapid Fluorescent Focus Inhibition Test (RFFIT) method.
RFFIT: The standard rabies virus was titrated in neuroblastoma cells for the assessment of immunogenicity. The vaccine immunized guinea pig sera were diluted in cell culture medium subsequently the serial dilutions of serum and a fixed amount of rabies virus were mixed in sterile 96 well culture plates and incubated at 37°C for 1 h. A suspension of neuroblastoma cells (0.05 mL) containing 25,000 cells was added to each well. Positive and negative controls were incorporated and the plates (as a reference) were incubated at 37°C for 48 h. The cell culture fluid was then removed, the cells fixed with acetone and kept at -20°C for 1 h. The monolayers in the plates were stained with fluorescin isothiocyanate (nutritional Biochemical Corporation Cleveland OH) conjugate at 37°C for 1 h. The conjugate was removed, the plates gently washed with distilled water to all the wells and 1 drop of mounting buffer was added and observed under UV fluorescence microscope (Smith et al., 1996). The immunogenicity of the formulated materials was determined by the inhibition of infective particles.
Safety: A double dose of the vaccine was administered to two guinea pigs from each formulation. They were observed for local as well as systemic reaction for a period of 14 days.
RESULTS AND DISCUSSION
The vero cells monolayer’s were inoculated and infected with PV-11 (Pasteur rabies virus) The virus was harvested from infected cultures from third day after inoculation as viral harvest, five times at intervals of 72 h. and the virus titers was 105.2 . Furthermore the pooled SVH are concentrated by the pellicon ultrafiltration system (Jagannathan et al., 2010).
Inactivation: β propiolactone (FERAK Berlin GmbH; NMR 98.5%) is highly reactive chemical because of the moderate fire hazard when exposed to heat or flame. β propiolactone is available in a grade containing 97% minimum active ingredient (Jones and Feldman, 1981).
The βPL are hydrolyzed in the water at the concentration of 1:2000 are gradually to compare with the other ratios, after 2 h the residual βPL are more than 50% in the 1:1000 ratio and under gone minus in the 1:3000 (Table 1). The residual βPL in +4°C in all ratio are similar linear gradient.
When the single addition of the entire calculated volume of βPL gets hydrolyzed before all the viruses are inactivated. The same volume of βPL as two additions can complete its process of inactivation within the same period of time.
The 1:2000 ratio was taken for the subsequent experiment, since all the three various temperatures based inactivation experiments. The level of residual βPL was not higher then other experiments those were conducted at various temperatures. The result of 1:2000 was showed linear others are not linear (Table 2). The βPL lot. Nos 18/97 and 20/97 is also showed the repeative results in all inactivation temperatures and the purity of βPL are varied from 1% only (Table 3).
Rabies virus has been inactivated by various chemical and physical agents for vaccine production. βPL has been used extensively for the production of rabies vaccines (Peck et al., 1951).
| Table 1: | Remaining (%) of βPL in water for injection |
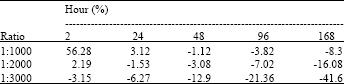 | |
| Common factors: -: Inactivation temperature-+37°C, Inactivation time: 2 h | |
| Table 2: | The inactivation of concentrated rabies viral harvests in different dilutions, temperature, time and the remaining % of βPL in different period |
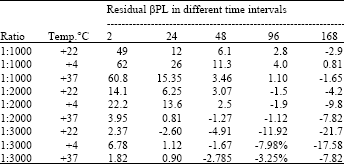 | |
| Common factors:- Inactivation time:- 2 h | |
| Table 3: | The inactivation of concentrated rabies viral harvests in 1:2000 dilution, temperature, time and the residual βPL in different interval, it containing the concentration of the βPL and their lot no |
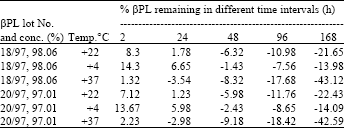 | |
| Common factors:- Inactivation ratio 1:2000, Agitation:-Yes, Inactivation time:-48 h | |
The βPL action on the cell substrate DNA has been extensively studied there has been very little analysis of its effects on chromosomal DNA and especially on the residual fraction co-purifying with virus during vaccine preparation (Morgeaux et al., 1993). The treatment with βPL of continuous cell lines can reduce the hazards associated with such containing residual cellular DNA (Perrin and Morgeaux, 1995).
Avirulance test: The entire different samples those are inoculated to intracerabrally and also different interval. Animals are not showed any symptoms; few test animals are observed upto 42 days, upto those days that animals are not showed any rabid symptom. These results are represents the titre of the virus. It states that all active virulence is inactive in 1:2000 ratios by the process of βPL inactivation.
Immunogenicity: For the analysis of neutralizing antibody the guinea pigs were immunized with vero cell derived material and the sera were analyzed by the RFFIT method. The antibody titre in International units of the serum samples for Vero cell derived material was found above required level. A two fold dose of the vaccine formulations when injected to susceptible sero negative guinea pigs did not produce any adverse reactions.
However, inactivation with βPL is considered to be the result of alkylation of imidazole functional groups in the viral ribonucleic acid The total number of hours required for inactivation also decreases with an increase in the use of biological (therapeutic proteins or vaccines) produced from Continuous Cell Lines (CCLs) has raised problems of safety. In particular, the presence of residual cell nucleic acid (rcDNA) is associated with hypothetical risk of oncogenicity. The fundamental reason for viral infectivity inactivation by propiolactone is carboxyethylation of nucleic bases within the viral genome (Chen et al., 1981). βPL treatment reduced the heamagglutination titer to one-half of the control value after 4 h of treatment; however, no further loss in hemagglutinin titer was detected during the subsequent 20 h of treatment. Complement fixation by the rabies virus concentrate remained at 100% of the control value, even after exposure to βPL for 24 h (Wiktor et al., 1972).
Base carboxyethylation reactions are kinetically single-step and irreversible under storage conditions for inactivation antiviral vaccines. Therefore the infectivity of the inactivated virus cannot be restored by genome demodification during storage of vaccines prepared with the aid of propiolactone, which ensures the stable safety of the preparation. The kinetics of inactivation of RNA-containing viruses reflects the kinetics formation of the first inactivating lesion in the genome. (Budowsky et al., 1991).
Those samples were inactivated by 1:2000, 48 h with agitation are gave the better result, to compare with the other test samples. The residual βPL also decreased after 48 hrs that are very less to compare with other samples. This experiment was done only in the ratio of 1:2000 and the time of the inactivation also very less to compare with the other different samples. The results also indicate that the rate of inactivation is directly proportional to the concentration of βPL used in the process (Aarthi et al., 2004); the total number of hours required for inactivation also decreases with an increase in the concentration of βPL. All of the inactivating experiments are in which the effect is time-dependent. As per the regulatory norms the ratio of the βPL and viral harvest ought to not above the limit.
The titer of the virus was decreased at the inactivation temperatures like +22 and 37°C, the hydrolysis of βPL also occurring at 37°C temperature. Exposure at the 37°C helps in hydrolyzing the βPL which helps in avoiding the step of addition of thiosulphate for neutralization. And also the βPL does not require any neutralizing agent to stop its reaction since it is completely hydrolyzed. It is more quickly degraded in the presence of cellular debris and cell culture media. The rate of βPL hydrolysis increases in the presence of proteins. These decreasing viral titers directly reflect the yield that seems in the final potency of the vaccine. During the experiments the pH of the samples are not increased drastically but slightly and maintained as the neutral pH, the inactivation rate decreased linearly with increase in pH, whereas the 99% viral inactivation time increased proportionately with increase in pH. the rate of the viral inactivation is pH dependent (faster in acid than in alkaline medium), having a fractional order with respect to both [H+] and [OH¯]; When BPL was used as the inactivating agent, infectivity was exponentially reduced 100,000-fold during the first 6 hr, whereas complement-fixing activity remained similar to that of the untreated vaccine (Wiktor et al., 1972). All the inactivated antigens are formulated with 1% human albumin and 5% maltose. The proteins present in the liquid rabies vaccine are not an influential factor for the efficacy of vaccines. Only its immunogenicity of inactivated, purified rabies viral proteins plays the main role in the efficacy (Jagannathan et al., 2011). The immunogenicity of the vaccines is above the prescribed limit. And also the experimented vaccines are not produced any adverse reaction in guineas pigs it seem the safe of the vaccine.
CONCLUSION
In the vaccine manufacturing industry the time is a crucial factor because to get more inactivated viral harvest are yield in a short period and they are processed to further purification and downstream processing, this findings are to reduce the time frame and the residual βPL in various level are influenced the tissue culture derived anti rabies vaccine. Since inactivated vaccines are more stable and easier than other attenuated virus vaccines. These results are reflecting the safety, potency and efficacy of the Vero cell culture derived rabies vaccine. These results are also encouraging and more studies are needed for the shortening the time duration during inactivation.
ACKNOWLEDGMENT
The author’s sincere thanks to the Director, Pasteur Institute of India, Coonoor, for providing facilities to carry out the study and acknowledged.
REFERENCES
- Aarthi, D., K. Ananda Rao, R. Robinson and V.A. Srinivasan, 2004. Validation of binary ethyleneimine (BEI) used as an inactivant for foot and mouth disease tissue culture vaccine. Biologicals, 32: 153-156.
CrossRef - Budowsky, E.I. and M.A. Zalesskaya, 1991. Principles of selective inactivation of viral genome. V. Rational selection of conditions for inactivation of the viral suspension infectivity to a given extent by the action of β-propiolactone. Vaccine, 9: 319-325.
CrossRef - Budowsky, E.I., E.A. Friedman, N.V. Zheleznova and F.S. Noskov, 1991. Principles of selective inactivation of viral genome. VI. Inactivation of the infectivity of the influenza virus by the action of β-propiolactone. Vaccine, 9: 398-402.
CrossRef - Chen, R.F., J.J. Mieyal and D.A. Goldthwait, 1981. The reaction of β-propiolactone with derivatives of adenine and with DNA. Carcinogenesis, 2: 73-80.
CrossRefDirect Link - Hiatt, C.W., 1964. Kinetics of the inactivation of viruses. Bacteriol. Rev., 28: 150-163.
Direct Link - Hassanzadeh, S.M., A. Zavareh, M.A. Shokrgozar, A. Ramezani and A. Fayaz, 2011. High vero cell density and rabies virus proliferation on fibracel disks versus cytodex-1 in spinner flask. Pak. J. Biol. Sci., 14: 441-448.
CrossRefPubMedDirect Link - Jagannathan, S., S. Chaansha, K. Rajesh, T. Santhiya, C. Charles and K.N. Venkataramana, 2009. Standardization and assessment of cell culture media quantities in roller poly ethylene terephthalate bottles employed in the industrial rabies viral vaccine production. Pak. J. Biol. Sci., 12: 1246-1252.
CrossRefDirect Link - Jagannathan, S., R. Vijayakumar, P.R. Gandhi, M. Ananthi, C. Charles, A. Premkumar and K.N. Venkataramana, 2010. Analysis of carbohydrates in newly developed liquid state rabies vaccine. J. Microb. Biochem. Technol., 2: 147-151.
Direct Link - Jagannathan, S., K.R. Mani, R. Vijayakumar, P.R. Gandhi and K.N. Venkataramana, 2011. Comparative analysis of magnesium chloride in recently developed liquid State Rabies vaccine. Int. J. Pharma Bio Sci., 2: 473-479.
Direct Link - Morgeaux, S., N. Tordo, C. Gontier and P. Perrin, 1993. β-propiolactone treatment impairs the biological activity of residual DNA from BHK-21 cells infected with rabies virus. Vaccine, 11: 82-90.
CrossRef - LoGrippo, G.A., 1960. Investigations of the use of beta-propiolactone in virus inactivation. Ann. N. Y. Acad. Sci., 83: 578-594.
CrossRef - Sheng-Fowler, L., A.M. Lewis Jr. and K. Peden, 2009. Quantitative determination of the infectivity of the proviral DNA of a retrovirus In vitro: Evaluation of methods for DNA inactivation. Biologicals, 37: 259-269.
CrossRef - Perrin, P. and S. Morgeaux, 1995. Inactivation of DNA by β-propiolactone. Biologicals, 23: 207-211.
CrossRef - Reed, L.J. and H. Muench, 1938. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol., 27: 493-497.
CrossRefDirect Link - Sudarshan, M.K., S.N. Madhusudana, B.J. Mahendra, N.S.N. Rao and D.H. Ashwath Narayana et al., 2007. Assessing the burden of human rabies in India: Results of a national multi-center epidemiological survey. Int. J. Infect. Dis., 11: 29-35.
Direct Link - Trabelsi, K., S. Rourou, H. Loukil, S. Majoul and H. Kallel, 2005. Comparison of virous culture modes for the production of rabies virus by Vero cells grown on microcarriers in a 2-l bioreactor. Enzyme Microb. Technol., 36: 514-519.
CrossRef - Tyler, W.P. and D.W. Beesing, 1952. Chemical and cryoscopic analysis of β-propiolactone. Anal. Chem., 24: 1511-1513.
CrossRef - Wiktor, T.J., H.G. Aaslestad and M.M. Kaplan, 1972. Immunogenicity of rabies virus inactivated by β-propiolactone, acetylethyleneimine and ionizing irradiation. Applied Microbiol., 23: 914-918.
Direct Link