
Ahmad Gharzi
Department of Biology, Faculty of Science, Lorestan University, Khorramabad, Iran
Mohsen Abbasi
Department of Basic Science, Faculty of Veterinary Medicine, Lorestan University, Khorramabad, Iran
Parisa Yusefi
Department of Biology, Faculty of Science, Lorestan University, Khorramabad, Iran
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 5 | Page No.: 372-378







ABSTRACT
The vomeronasal organ plays a significant role in many aspect of snakes’ life including pray detection, feeding and reproduction. In this study the anatomy and histology of the vomeronasal organ in an aglypha snake, Typhlops vermicularis, was investigated. To perform this task, 5 individuals of the snake were collected from their natural habitat and transferred to the laboratory. After classification, 5-7 micrometer histological sections were cut from their heads and the sections were stained through two different protocols. The results showed that in this taxon the vomeronasal organ has a mean length of 630 Mm and located on the baso-medial side of nasal cavity. The organ is enclosed by a bony capsule but this capsule is not complete in some regions where it is replaced by a cartilage or connective tissue. In this species, the organ has no connection to the nasal cavity but communicate with the oral cavity through two tiny openings in the anterior palate. At the connection point the lacrimal duct also joins to the organ canal. The organ’s lumen is lined by two different sensory and respiratory epitheliums. The sensory epithelium is very thick and contains three types of cells; receptor, supportive and basal. The epithelium which covers the mushroom body consists of a layer of ciliated cells. Moreover, in this taxon no PAS-positive materials were found in the lumen of the organ.
PDF Abstract XML References Citation
Received: March 09, 2013;
Accepted: April 04, 2013;
Published: July 12, 2013
How to cite this article
Ahmad Gharzi, Mohsen Abbasi and Parisa Yusefi, 2013. Histological Studies on the Vomeronasal Organ of the Worm-like Snake, Typhlops vermicularis. Journal of Biological Sciences, 13: 372-378.
DOI: 10.3923/jbs.2013.372.378
URL: https://scialert.net/abstract/?doi=jbs.2013.372.378
DOI: 10.3923/jbs.2013.372.378
URL: https://scialert.net/abstract/?doi=jbs.2013.372.378
INTRODUCTION
The sense organs in snakes are totally varies from those of other vertebrates. In contrast to mammals which are largely dependent to their sight and hearing, snakes mostly use their smell and touch senses for detection (Grzimek et al., 2003). They do not have moveable eyelids and the movement of their eye is quite restricted. Moreover, They lack some parts of ear including external ear, middle ear and tympanic membrane and as a result they sense only restricted vibrations of sound waves (Gove, 1979; Halpern, 1992). In contrast to sight and hearing, snakes are well equipped in smell sense. In addition to the main olfactory system which consists of nostrils, nasal cavities and olfactory bulb, they have an accessory chemosensory system known as the Vomeronasal (VN) system (Halpern and Martinez-Marcos, 2003; Martinez-Garcia et al., 1991). The main part of the VN system contains a pair of Jacobson’s or Vomeronasal Organs (VNO) located at the roof of the oral cavity. The flicking forked tongue of snakes is used to sample and bring minute air particles into contact with the neurosensory epithelium of this organ and the snake then perceives and identifies the smell as prey, predator, or otherwise (Doving and Trotier, 1998). In some snakes, the chemical and odorants on the lips related to their foods reach to neurosensory epithelium of the organ through diffusion (Kent and Carr, 2000).
The VNOs are found in most tetrapods lineages including amphibians (Kralovec et al., 2013), reptiles (Graves, 1993; Halpern and Borghjid, 1997) and many groups of mammals, including rodents (Bhatnagar and Smith, 2010; Lee et al., 2008; Ogura et al., 2010; Wekesa, 2012; Wynn et al., 2012) rabbits (Taniguchi et al., 2000), ungulates (Abass et al., 2012; Abbasi, 2007), monkeys (Smith et al., 2011) and even humans (Keverne, 1999) but they are not observed in fishes and birds (Doving and Trotier, 1998). These organs are highly developed in squamate reptiles such as lizards and snakes. The VN chemoreception is extremely essential for squamates’ life as mediating literally every important behavior, particularly those related to foraging, feeding and reproduction (Filoramo and Schwenk, 2009).
In squamates, the VN chemosensory system is anatomically and functionally distinct from the main olfactory system. Unlike other vertebrates, in Squamata the paired VNOs has no direct connection to the main olfactory system (Schwenk, 1995). In addition, the VN nerves project to a separate part of the olfactory bulb (accessory olfactory bulb) and remain separate within the central nervous system until at least the amygdaloid nuclei (Martinez-Marcos et al., 1999). A significant functional discrepancy between the two chemosensory systems is that the VN system can be stimulated by large, nonvolatile molecules that would not typically reach the main olfactory system during inhalation. Thus, the VN system is frequently viewed as specialized for the reception of high molecular weight, nonvolatile chemicals. On the contrary, the main olfactory system is well developed for sensing volatile chemicals that distribute into the air very easier and move better to farther distances (Martinez-Garcia et al., 1991).
Several researches have been executed on the VNO of snakes (Miller and Guzke, 1999; Takami and Hirosawa, 1990; Young, 1993) but most of these studies are related to Garter snake or venomous snakes like vipers. To illustrate, in Garter snakes, the VNO is involved in location and trailing of pray prior to strike, in courtship, in aggression behavior and in movement within home ranges (Alving and Kardong, 1996; Halpern and Borghjid, 1997). In rattles snakes this organ plays a role in regulating pre and post strike behaviors and the blockage of VN duct results to complete elimination of these behaviors (Graves and Duvall, 1985).
Some biological aspects of the aglypha snake, Typhlops vermicularis have been studied (Amr and Disi, 2011). However, there are no published reports on the VNO of this taxon. The aim of the present study was to examine the anatomical and histological structure of the VNO in order to shed light on this snake which has good potential for use as animal research model. The worm-like snake, T. vermicularis displays a maximum length of 350 mm with a cylindrical body covered with smooth scales of even size. Eyes in this snake are degenerated and snout is spherical. The color of this species is light pink-gray, although on the ventral side it is slightly lighter (Amr and Disi, 2011). This species is distributed in southeast Europe, southwest Asia and north Africa. In Iran this snake is found in east Azerbaijan, Central, Fars, Guilan, Mazandaran, Kerman, Khozestan Provinces (Latifi, 1991).
MATERIALS AND METHODS
Five adults (3 males and 2 females) of worm-like snakes were collected from their natural habitat at the vicinity of Siahkaleh village, Dorood, in Lorestan Province and transferred to the laboratory. The specimens were then killed by chloroform. Following decapitation, the heads were washed with normal saline and then fixed in 4% formalin saline for at least a week. The specimens were later decalcified using formic acid-sodium citrate solution for 3-4 days according to Smith et al. (1997). The tissues were dehydrated in graded series of ethanol, cleared in xylene and embedded in paraffin wax. The blocks were sectioned in transverse plane at 7 μm thickness. The sections were mounted on glass slides and stained with Haematoxylin-Eosin or Alcian blue-Periodic Acid Schif (PAS). Then stained sections were scrutinized in details by the aid of a light microscope. Ocular graded graticule was used to measure specific histological parameters in the organs including the lumen’s width and distance between the two capsules enclosed the organs.
RESULTS
In the Typhlops vermicularis, the VNO consists of a paired bilateral structure located at the baso-medial side of the nasal cavity, with direct relationship with the vomer bone and palatine process of maxillary bone. In general, each organ appeared as a hollow spindle with an average length of 630 μm. Each organ begins at rostral one-third of the nasal cavity. Anteriorly, the organ is narrow but gradually widens. This increase in diameter continues to two-third of the organ while in its caudal one-third it undergoes a reverse process so that it again becomes tapered (Table 1).
In transversal sections the organ is observed as a dome-shaped structure which is enclosed by an osseous capsule composed of medial palatine and lateral vomer bones. The capsule is incomplete as in some regions the bone is replaced by connective or cartilage tissues. The sites and locations of these replacing tissues varies along the organ’ length but mostly is seen in dorsolateral and ventromedial regions. The lumen of organ has become crescent-shaped as the result of Mushroom Body (MB) which is a bulge-like evagination protrudes from ventral or ventrolateral side into the lumen (Fig. 1a).
Along the length of the organ the lumen is lined with three types of epithelia. The dorsal or concave side of the VNO lumen is lined with a sensory epithelium whose main role is to detect chemical and elicit impulses to be conducted into the central nervous system. This is a pseudo stratified epithelium in which three types of cells can be recognized. Just beside the lumen there is a layer of supportive cells which have elongated nuclei. At the top of supportive cells a several layers of sensory cells (on average 10 layers) with rounded nuclei are observed. The receptor cells are indeed bipolar neurons whose dendrites terminate as microvilli at the luminal surface.
| Table 1: | Measurements of the nasal cavity and VNO parameters in μm |
 | |
| NCD: Diameter of nasal cavity, DVL: Diameter of VNO lumen | |
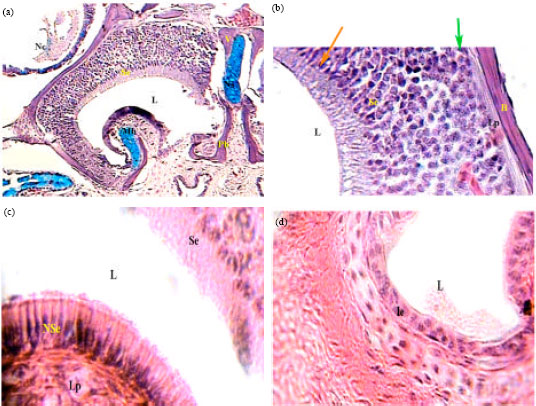 | |
| Fig. 1(a-d): | Histology of the VNO in T. vermicularis, (a) A cross section through the medial segment of the VNO which shows sensory epithelium at the top and the mushroom body (Mb) at the bottom. The VNO is enclosed by an incomplete capsule, (b) Higher magnification of the sensory epithelium composed of three types of cells: supportive (red arrow), sensory (Se) and basal (green arrow). This epithelium is laterally covered by a connective tissue (LP), (c) The non-sensory (NSe) or respiratory epithelium contains a simple layer of ciliated cells and surrounds the mushroom body, (d) A stratified (2 rows) cuboidal epithelium is seen on the lumen wall between the sensory and non-sensory epithelia. Lumen (L), Nasal cavity (NC), Nasal septum (NS), Mucous glands (Mg) at the roof of mouth, Palatine bone (Pb), Oral cavity (Oc), Vomer bone (V). PAS-Alcian blue staining in A and B, H and E in C and D. A= 100x, B= 600x and C-D = 400x |
The microvilli themselves could not be visualized with the optics used for this experiment due to their small diameter, 100 nm. Finally, around these sensory cells a layer of basal cells is observed. This sensory epithelium itself is laterally covered by a thin loose connective tissue (Fig. 1b).
The ventral or convex side of the VNO lumen is lined with a non-sensory but respiratory epithelium which covers the MB (Fig. 1c). This epithelium is made of simple layer of ciliated columnar cells. Beneath the MB there is a thick loose connective tissue which displays profuse vascularization. This represents venous sinuses with a prominent large vein amongst smaller arteries, capillaries and venules. Between the sensory and non-sensory epithelia the wall of lumen is lined with a stratified (2 rows) cuboidal epithelium (Fig. 1d).
In cross sections, three segments with different anatomical and histological features were observed between the rostral and caudal ends of the VNO. At the rostral segment the VNO in much smaller than the nasal cavity as its different regions can not be clearly distinguished. Here the organ’s capsule is oval and made of different tissues at various sides (Fig. 2a). At dorsolateral and ventral sides the capsule’s wall is bony whereas at medial side the bone is replaced by the cartilage of nasal septum (Fig. 2a). As we move backward the shape of the capsule changes and becomes dome-shaped in the medial segment. In this part the extension of the vomer has been disappeared in basal side and the palatine turned into two L-like bone pieces which make the basomedial wall of the capsule (Fig. 1a). In the middle segment, in addition to medial side, the replacing tissue which here is a connective tissue covers ventral and dorsal walls.
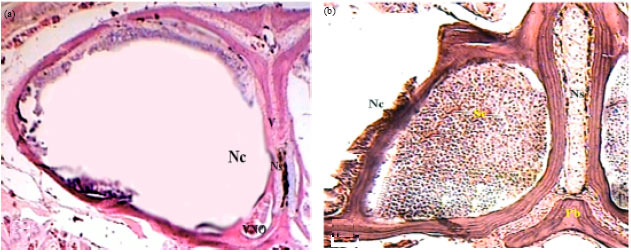 | |
| Fig. 2(a-b): | Regional changes in the VNO structure, (a) In rostral segment the organ (VNO) lies at the basomedial side of nasal cavity (Nc), (b) In caudal segment the lumen disappears and is taken up by sensory epithelium (Se). At this point the organ’s capsule is complete. Ns: Nasal sepyum, V: Vomer bone, Bs: Blood sinuses, Ct: Connective tissue, H and E, A= 40x, B = 100x |
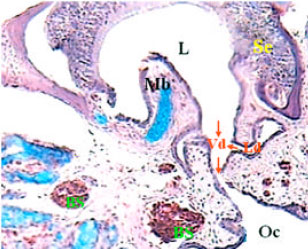 | |
| Fig. 3: | Connection of the VNO lumen with the oral cavity. A cross section through rostral segment of the organ which shows the linkage canal between the VNO and oral cavity (Oc). Arrows display the movement direction of secretory materials into the mouth. Within the connective tissue located beneath the Mushroom body (Mb) blood sinuses(BS) are visible. Vomeronasal duct (Vd), Sensory epithelium (Se). H and E, 100x |
At this point a wide lumen is seen in the organ (Fig. 1a) and the MB displays a cartilaginous supportive tissue. The type of this cartilage is hyaline and many mitotic figures are observed in its constituent cells.
Along the length of caudal segment, the MB gradually disappears and the lumen starts becoming narrower. At the caudal end, the lumen is completely eliminated and occupied by a thick stratified epithelium (Fig. 2b). In this region the osseous capsule is oval in shape, its walls are complete and the vomer returns to its initial shape. Throughout the length of the paired organs the space between two left and right capsules remains unchanged (Table 1).
All over the organ’s length no connection is found between the organ and nasal cavity. However, there is a linkage canal between the VNO and oral cavity where through which the mouth connects to the VNO lumen. These openings are located on the anterior part of palate. At the connection point a lacrimal duct also joins to the canal (Fig. 3). The lacrimal and VNO ducts both are lined with simple squamous epithelium.
DISCUSSION
The anatomy and histology of the VNO in snakes, particularly venomous species, has been established by some workers (Takami and Hirosawa, 1990; Taniguchi et al., 2000; Young, 1993). The comparison of their findings with the results presented here clarifies that the VNO structure in T. vermicularis in general is similar to that observed in venomous snakes (Alving and Kardong, 1996; Taniguchi et al., 2000). To illustrate, in present study it was shown that the VNO contains sensory epithelium for chemoreception and non-sensory or respiratory epithelium which is in agreement to reports performed on venomous snakes (Alving and Kardong, 1996). Moreover, we here demonstrated that like other squamates, in this taxon the VNO has no connection to nasal cavity and moves at the roof of the mouth independently but communicates solely with the oral cavity through two tiny openings (vomeronasal fenestrae) in the anterior palate (Filoramo and Schwenk, 2009). Another comparable point in venomous and non-venomous snakes is that the sensory epithelium in both snakes is made of three types of cells; supportive, sensory and basal, arranged in similar way around the organ’ lumen (Alving and Kardong, 1996). These observation concerning the respiratory covering of the MB is supported by similar findings in Garter snake, rattlesnake and other studied squamates (Halpern, 1992; Schwenk, 1995). It is also demonstrated here that in contrast to the non-sensory, the sensory epithelium is made of about 15 cell layers and it confirms previous studies carried out on reptiles and mammals. Owing to several investigations on different groups of vertebrates it has been proved that the thickness of sensory tissue on reptiles is generally higher when compared to that of mammals. In the same way, it is believed that compared to lizards, this epithelium in thicker in snakes. A reason for these differences is that sight and hearing senses in snakes and lizards are very weaker than those in mammals and similarly these senses in snakes are again much weaker than that of lizards (Gove, 1979, Halpern, 1992; Kratzing, 1975; Rehorek, 1997; Schwenk, 1993).
In present study, it is demonstrated that the total length of the VNO is 630 μm. In addition, it is showed that the maximum diameter is seen in the middle segment and as it moves forward and backward this parameter reduces. Comparison of these measurements with those from the snake, Eirenis collaris (Abbasi et al., 2011) suggests that the length of the VNO in snakes is correlated to the snout length so that the snakes with longer snout have longer VNO.
In present study, it was shown that at the connection point of the VNO duct and oral cavity a lacrimal duct also joins to the duct and this against observations made in mammals whose lacrimal duct opens into nasal cavity and has no connection with the mouth (Doving and Trotier, 1998). Previous studies performed on reptiles have shown a connection between the VNO and lacrimal ducts but the location of this joining point varies among different orders. In lizards the lacrimal duct opens into the VNO lumen via an opening observed just beside the MB whereas, in snakes the opening lies on the concave wall, opposite to the MB (Bellairs and Boyd, 1947). In the studied taxon, like other snakes this opening is on the opposite side of the MB (Fig. 3).
Many capillaries, several arterioles and large veins were observed in connective tissue lies beneath the MB (Fig. 3) which is in accordance with the findings in others animals (Altner et al., 1970). This makes the VON a highly vascularized structure. Such an irrigated structure as the VNO with similarities to the neighboring nasal mucosa suggests that the VNO in T. vermicularis can be an erectile tissue. In current study it could not recognized any blood vessels within the epithelia of the VNO. However, such intra-epithelial blood vessels have been described in mammals. It has been proposed that the presence of intraepithelial blood vessels is to provide nourishment for the thick epithelium and permit exchange of metabolites (Igbokwe and Nwaogu, 2009). According to Bellairs and Boyd (1947) the lumen of VNO in snake, Pseudonaja textiles is filled with PAS-positive materials possibly derived from Harder’s gland (an accessory lacrimal gland). In case of T. vermicularis, the VNOs stained with PAS method display no such materials in the lumen. In mammals excretory ducts of many glands dorsally or ventrally open into the VNO lumen but in the studied species like other snakes these glands are not present.
Another noticeable observation made in this study was that the osseous capsule which encases the organ was not bony in all regions but replaced by a connective or cartilaginous tissue, a remark has been made in some reptiles and mammals (Rehorek, 1992; Slaby, 1984). In the studied snake, this replacing tissue is observed clearly on medial segment of the organ and in this respect it resembles Coluber natrix (Doving and Trotier, 1998) and Eirenis collaris (Abbasi et al., 2011). However, in contrast to Eirenis collaris which the nasal septum cartilage fills the gap between bony pieces, here in T. vrmicularis, like Coluber natrix a connective tissue occupies this space.
CONCLUSION
Collectively, in the species examined here, it is showed that the sensory epithelium which contains three cell types is very thick and this can be related to life’s style of this species. This taxon normally lives underground and in dark environment where for feeding it mainly relies to its chemoreceptory system rather than the eyes. Unlike the sensory, the respiratory epithelium observed at the top of MB consists of just a single layer of cells. Moreover, an intermediary stratified epithelium covers the interval between the MB and lateral walls. All these epithelia are encased by an incomplete osseous capsule. In general, it became clear that in T. vermicularis, despite some differences, the VNO displays the same anatomy and histology as found in other snakes.
ACKNOWLEDGMENT
This study was financially supported by Lorestan University which is deeply appreciated.
REFERENCES
- Abass, T.A., M.S. Al-Mayahi and B.F. Al-Hussany, 2012. Anatomical and histological investigate of Vomeronasal Organ (VNO) in Iraqi sheep Alawasi. Kufa J. Vet. Med. Sci., 3: 98-112.
Direct Link - Abbasi, M., 2007. The vomeronasal organ in buffalo. Ital. J. Anim. Sci., 6: 991-994.
CrossRefDirect Link - Abbasi, M., A. Gharzi and P. Usefi, 2011. Histomorphometry of vomeronasal organ in the snake, Eirenis collaris. Res. J. Biol. Sci., 6: 432-435.
Direct Link - Alving, W.R and K.V. Kardong, 1996. The role of the vomeronasal organ in rattlesnake (Crotalus viridis oreganus) predatory behavior. Brain Behav. Evol., 48: 165-172.
PubMedDirect Link - Amr, Z.S. and A.M. Disi, 2011. Systematics, distribution and ecology of the snakes of Jordan. Vertebrate Zool., 61: 179-266.
Direct Link - Bellairs, A.D.A. and J.D. Boyd, 1947. The lachrymal apparatus in lizards and snakes.-I. The brille, the orbital glands, lachrymal canaliculi and origin of the lachrymal duct. Proc. Zool. Soc. London, 117: 81-108.
CrossRef - Bhatnagar, K.P. and T.D. Smith, 2010. The human vomeronasal organ. Part VI: A nonchemosensory vestige in the context of major variations of the mammalian vomeronasal organ. Curr. Neurobiol., 1: 1-9.
Direct Link - Doving, K. and D. Trotier, 1998. Structure and function of the vomeronasal organ. J. Exp. Biol., 201: 2913-2925.
Direct Link - Filoramo, N.I. and K. Schwenk, 2009. The mechanism of chemical delivery to the vomeronasal organs in squamate reptiles: A comparative morphological approach. J. Exp. Zool. Part A: Ecol. Genet. Physiol., 311: 20-34.
CrossRefDirect Link - Gove, D., 1979. A comparative study of snake and lizard tongue-flicking, with an evolutionary hypothesis. Zeitschrift fur Tierpsychologie, 51: 58-76.
CrossRef - Graves, B.M., 1993. Chemical delivery to the vomeronasal organs and functional domain of squamate chemoreception. Brain Behav. Evol., 41: 198-202.
CrossRef - Halpern, M. and S. Borghjid, 1997. Sublingual plicae (anterior processes) are not necessary for garter snake vomeronasal function. J. Comp. Psychol., 111: 302-306.
PubMedDirect Link - Halpern, M. and A. Martinez-Marcos, 2003. Structure and function of the vomeronasal system: An update. Prog. Neurobiol., 70: 245-318.
PubMedDirect Link - Igbokwe, C.O. and I.C. Nwaogu, 2009. Histological studies of the vomeronasal organ of African giant rat (Cricetomys gambianus, Waterhouse). Anim. Res. Int., 6: 1003-1008.
Direct Link - Kralovec, K., P. Zakova and V. Muzakova, 2013. Development of the olfactory and vomeronasal organs in Discoglossus pictus (Discoglossidae, Anura). J. Morphol., 274: 24-34.
CrossRef - Kratzing, J.E., 1975. The fine structure of the olfactory and vomeronasal organs of a lizard (Tiliqua scincoides scincoides). Cell Tissue Res., 156: 239-252.
CrossRef - Lee, S.J., A. Mammen, E.J. Kim, A.Y. Kim and Y.J. Park et al., 2008. The vomeronasal organ and adjacent glands express components of signaling cascades found in sensory neurons in the main olfactory system. Mol. Cells, 26: 503-513.
PubMedDirect Link - Martinez-Garcia, F., F.E. Olucha, V. Teruel, M.J. Lorente and W.K. Schwerdtfeger, 1991. Afferent and efferent connections of the olfactory bulbs in the lizard Podarcis hispanica. J. Comp. Neurol., 305: 337-347.
CrossRef - Martinez-Marcos, A., E. Lanuza and M. Halpern, 1999. Organization of the ophidian amygdala: Chemosensory pathways to the hypothalamus. J. Comp. Neurol., 412: 51-68.
CrossRef - Miller, L.R. and W.H.N. Guzke, 1999. The role of the vomeronasal organ of crotalines (Reptilia: Serpentes: Viperidae) in predator detection. Anim. Behav., 58: 53-57.
CrossRef - Ogura, T., K. Krosnowski, L. Zhang, M. Bekkerman and W. Lin, 2010. Chemoreception regulates chemical access to mouse vomeronasal organ: Role of solitary chemosensory cells. PLoS ONE, Vol. 5.
CrossRefDirect Link - Schwenk, K., 1993. The evolution of chemoreception in squamate reptiles: A phylogenetic approach. Brain Behav. Evol., 41: 124-137.
CrossRef - Schwenk, K., 1995. Of tongues and noses: Chemoreception in lizards and snakes. Trends Ecol. Evol., 10: 7-12.
CrossRef - Slaby, O., 1984. Morphogenesis of the nasal apparatus in a member of the genus Chamaeleon L. (morphogenesis of the nasal capsule, the epithelial nasal tube and the organ of Jacobson in Sauropsida. VIII). Folia Morphol., 32: 225-246.
PubMedDirect Link - Smith, T.D., E.C. Garrett, K.P. Bhatnagar, C.J. Bonar and A.E. Bruening et al., 2011. The vomeronasal organ of New World monkeys (Platyrrhini). Anatomical Rec., 294: 2158-2178.
CrossRef - Takami, S. and K. Hirosawa, 1990. Electron microscopic observations on the vomeronasal sensory epithelium of a crotaline snake, Trimeresurus flavoviridis. J. Morphol., 205: 45-61.
CrossRef - Taniguchi, M., D. Wang and M. Halpern, 2000. Chemosensitive conductance and inositol 1,4,5-trisphosphate-induced conductance in snake vomeronasal receptor neurons. Chem. Senses, 25: 67-76.
Direct Link - Wekesa, K.S., 2012. Signaling in the mammalian vomeronasal organ. Biochem. Physiol., Vol. 1.
CrossRefDirect Link - Wynn, E.H., G. Sanchez-Andrade, K.J. Carss and D.W. Logan, 2012. Genomic variation in the vomeronasal receptor gene repertoires of inbred mice. BMC Genomics, Vol. 13.
CrossRef - Young, B.A., 1993. Evaluating hypotheses for the transfer of stimulus particles to Jacobson's organ in snakes. Brain Behav. Evol., 41: 203-209.
CrossRef