
Kimberly S. Beltran-Benjamin
Department of Biology, College of Arts and Sciences, University of the Philippines Manila, Padre Faura St. Taft Avenue, Ermita, Manila, Philippines
Elisa L. Co
Department of Biology, College of Arts and Sciences, University of the Philippines Manila, Padre Faura St. Taft Avenue, Ermita, Manila, Philippines
Sally Andrea D. Gaspi
Department of Biology, College of Arts and Sciences, University of the Philippines Manila, Padre Faura St. Taft Avenue, Ermita, Manila, Philippines
Jessica Leika R. Matibag
Department of Biology, College of Arts and Sciences, University of the Philippines Manila, Padre Faura St. Taft Avenue, Ermita, Manila, Philippines
Glenn L. Sia Su
Department of Biology, College of Arts and Sciences, University of the Philippines Manila, Padre Faura St. Taft Avenue, Ermita, Manila, Philippines
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 4 | Page No.: 183-195







ABSTRACT
The present study was designed to test the effect of the crude methanolic extract of Peperomia pellucida (L.) HBK on the antioxidant and redox enzymes. Twenty-five rats were divided into five groups that received different treatments. These rats were sacrificed and the liver was obtained to estimate the antioxidant superoxide dismutase (SOD) and catalase (CAT) and redox thioltransferase (TT) and thioredoxin reductase (TrxR) enzyme activities by spectrophotometry and to examine the physiological state of the liver by histopathological analysis. The crude methanolic extract of P. pellucida increased the activities of antioxidant enzymes (SOD and CAT) while redox enzyme (TT and TrxR) activities were greatly decreased. It was found out that 400 mg kg-1 b.wt. of P. pellucida extract was the most effective concentration and this finding was further supported by lesser incidence of fibrosis and necro-inflammation in the histopathological analysis of liver subjected to such dosage. Based on this result, it can be concluded that the crude methanolic extract of P. pellucida (400 mg kg-1 b.wt.) has a potent antioxidant activity because of its ability to elevate SOD and CAT and lower TT and TrxR activities under oxidative stress.
PDF Abstract XML References Citation
Received: March 14, 2013;
Accepted: April 04, 2013;
Published: July 10, 2013
How to cite this article
Kimberly S. Beltran-Benjamin, Elisa L. Co, Sally Andrea D. Gaspi, Jessica Leika R. Matibag and Glenn L. Sia Su, 2013. Enzyme Activity and Histopathology of Rat Liver Treated with Crude Methanolic Extract of Pepperomia pellucida (L.) HBK. Journal of Biological Sciences, 13: 183-195.
DOI: 10.3923/jbs.2013.183.195
URL: https://scialert.net/abstract/?doi=jbs.2013.183.195
DOI: 10.3923/jbs.2013.183.195
URL: https://scialert.net/abstract/?doi=jbs.2013.183.195
INTRODUCTION
The problem of providing the basic health care for individuals has constantly been a main issue for most developing countries. The World Health Organization (WHO, 2010) estimated that more than half of the world’s population does not have access to sufficient health care services. This may be due to the fact that compliance to present health care services is very expensive. Hence, the World Health Organization promotes the use of traditional health care to ensure the use of safe, effective and quality products and to guarantee patient safety by upgrading skills and knowledge in the use of traditional medicine (WHO, 2010).
In the Philippines, it is estimated that 1,500 plant species have medicinal value. One of these medicinal plants is a short-lived fleshy herb called Peperomia pellucida (L.) HBK and is more commonly known as pansit-pansitan@ or ulasimang bato@. In folk medicine, infusions of this plant are usually used to treat abdominal pain, rheumatic pain, gout and fatigue (Bojo et al., 1994). Several investigative researches have also validated the antimicrobial, anti-inflammatory, analgesic and antihyperuricemic activities of the plant (Bojo et al., 1994; Sio et al., 2001; Arrigoni-Blank et al., 2004). However, the effect of P. pellucida on antioxidant and redox enzymes has not yet been fully explored.
A recent study by Khan et al. (2010) showed that P. pellucida contained xanthones, particularly xanthone glycoside which exhibit inhibitory property towards lipid peroxidation and free radical scavenging ability. Thus, this study aims to test the antioxidative property of P. pellucida in rat liver undergoing oxidative stress by measuring the activities of selected antioxidant and redox enzymes and verifying findings through histopathological analysis.
MATERIALS AND METHODS
Preparation of the crude methanolic extract: Aerial parts, stem, leaves and flowers, of P. pellucida were collected from Matibag Farm in Brgy. Palsahingin, San Pascual, Batangas and were brought to the Botany Division, National Museum of the Philippines for identification and authentication.
The aerial parts of the plant were sun-dried for three days and oven-dried at 60°C. The dried plant materials were grounded into powder using mortar and pestle. The powdered materials (450 g) were soaked in 1000 mL 95% methanol for 48 h. The solution was filtered and evaporated using a rotary evaporator to obtain the crude extract. Phosphate buffer saline was added in increment amounts to the extract before administration to the test specimens.
Acclimatization of rats: Twenty five male Sprague-Dawley rats aged between 6-8 weeks and weighing 80-100 g were obtained from the department of Pharmacology and Toxicology, College of Medicine, University of the Philippines Manila. The rats were housed individually in metal cages in the animal house of College of Arts and Sciences of the same university. The rats were acclimatized for one week at standard conditions for temperature (28±2°C), relative humidity (50±5%) and light intensity (12 h light and dark cycles) (Avti et al., 2006). Water and food pellets were fed ad libitum throughout the experiment.
Induction of oxidative stress: Oxidative stress was induced using trichloroethylene (TCE) which was given orally through a syringe. The rats were treated with 0.75 mL kg-1 b.wt. of TCE daily for two weeks except for group 2. After which, treatment with P. pellucida extract followed.
Treatment protocol: Twenty five acclimatized rats were divided into 5 groups with 5 individuals each as follows: group 1 was the positive control group treated with vitamin E at 25 IU kg-1 b.wt. or 0.07 mL kg-1 b.wt.; group 2 was treated with P. pellucida extract only at 400 mg kg-1 b.wt., its effective dose as studied by Arrigoni-Blank et al. (2004), groups 3 and 4 were treated with 400 and 800 mg of P. pellucida (L.) HBK extract kg-1 b.wt., respectively and group 5 was the negative control group treated with 1 mL physiologic saline as stated by Pavlovic et al. (2009). Vitamin E, P. pellucida extract and placebo were injected intraperitoneally for 14 days daily. After 14 days, the rats were restrained and anesthesized with Zoletil 50 at 30 mg kg-1 b.wt. All methods were carried out following the guidelines set by the IACUC and Ethics Review Board of the National Institutes of Health, UP Manila.
Enzymatic assays of antioxidant and redox enzymes in rat liver: Rats were sacrificed via., exsanguinations. The liver was excised and perfused with an ice-cold perfusion solution (0.15 M KCl, 2 mM ethylenediaminetetraacetic acid [EDTA], pH 7.4). Portion of the liver (right, median and caudate lobes) obtained was subjected to homogenization in Tris-HCl buffer (50 mM, pH 7.4) and centrifugation at 10 000 Hg at 4°C for 30 min to get the Post-Mitochondrial Supernatant (PMS) (Avti et al., 2006). The PMS was used for the estimation of the different antioxidant enzyme concentrations. Protein concentrations were determined by Bradford Assay using bovine serum as standard.
Estimation of superoxide dismutase (SOD) activity: Superoxide dismutase activity was determined following the procedure in the study of Kim et al. (2003). The reaction mixture, measuring 0.2 mL, contained 0.01 U mL-1 of xanthine oxidase, 50 mM phosphate buffer (pH 7.4), 0.1 mM EDTA, 1 FM catalase, 0.05 mM xanthine, 20 FM cytochrome C and PMS. The change in absorption at 550 nm was monitored for 3 min and performed in triplicate. Superoxide dismutase activity was presented as change in absorbance of 550 nm/min/mg protein.
Estimation of catalase (CAT) activity: Catalase estimation was assayed by monitoring the decrease in absorbance at 240 nm due to hydrogen peroxide consumption Kim et al. (2003) and was done in triplicate. Three milliliter reaction mixture contained 100 mM phosphate buffer (pH 7.0) and 10 mM H2O2. It was incubated for 2.5 min at 30°C and was initiated by adding 0.05 mL of PMS. The change in absorbance over 3 min was recorded and performed in triplicate.
Estimation of thioltransferase (TT) activity: With the use of Glutathione Reductase (GR) as a coupling enzyme, thioltransferase was first measured spectrophotometrically at 340 nm as standard. The total volume must be 0.2 mL and the reaction mixture contained 1 mg mL-1 Bovine Serum Albumin (BSA), 10 mM reduced glutathione (GSH), 60 μg mL-1 yeast Glutathione Reductase (GR), 15 mM 2-hydroxyethyl disulfide (HED), 4 mM nicotinamide adenine dinucleotide phosphate (NADPH) and 1 M Tris-HCl-20 mM EDTA at pH 8.0. The change in absorbance for 3 min was recorded and performed in triplicate (Kim et al., 2003).
Estimation of thioredoxin reductase (TrxR) activity: Thioredoxin reductase activity was measured as the reduction of 5, 5'-dithio-bis(2-nitrobenzoic acid) (DNTB) in the presence of NADPH as cited by Hong-Gyum Kim et al. (2003). The assay mixture contained 0.2 M phosphate buffer with pH 7.6, 1 mM EDTA, 0.25 mM NADPH and 1 mM DNTB. The increase in absorbance was measured at 412 nm for 3 min at 25°C and performed in triplicate.
Histopathology of rat liver: The largest left lobe of the excised liver was placed in a cooler filled with ice. It was immediately fixed in 10% formalin for 24 h, dehydrated in graded ethanol concentrations and embedded in paraffin wax. Sagittal sections (5 μm of thickness were cut and mounted on glass slides. Sections were deparaffinized in xylene, hydrated in ethanol and stained with Hematoxylin-Eosin (HE) method and the prepared slides were viewed under light microscopy with 400x magnifications and evaluated for histopathological changes on the effect of P. pellucida crude methanolic extract in rat liver. The hepatic tissue changes were graded using the modified Knodell’s Scoring System as cited in Brunt (2000).
Statistical analysis: Data were analysed using SPSS 17.0 for Windows (SPSS Inc., Chicago, USA). Values were expressed as mean of enzymatic activities±SEM. Analysis of variance was performed to analyze if there is any difference between the means (p<0.05). Tukey multiple comparison test was used to determine which of the means are different. The p<0.05 will be considered significant.
Values, depending on the scoring, were expressed as mean. Kruskal-Walis one way analysis of variance by ranks was used to analyze if there is any difference between the means of the groups, where p<0.05 was considered significant. Mann-Whitney test was used to determine which of the means are different, where U<0.05 was considered significant.
RESULTS
Enzymatic analysis: Both antioxidant and redox enzymes in the rat liver were measured in the different treatment groups. Figure 1 shows a summary of the different enzymes measured in the rats under different treatments. Results show that antioxidant and redox enzymes exhibit contrasting trends in the different groups.
In Fig. 2a, the estimated SOD activity in the rat liver of those treated with crude methanolic extract of P. pellucida (800 and 400 mg kg-1 b.wt.; 0.106±0.013 and 0.150±0.008/min/mg protein, respectively) showed significant increase when compared with the negative control group (0.067±0.019/min/mg protein shown in Fig. 2). However, SOD activity in group 4 is significantly lower than that of group 3. Furthermore, the SOD activities of both group 3 and the positive control group (0.07 mL kg-1 b.wt. of vitamin E; 0.146±0.026/min/mg prot) are not significantly different.
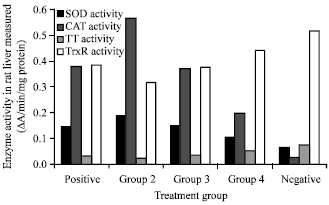 | |
| Fig. 1: | Summary of the estimation of both antioxidant and redox enzymes measured in the rat liver after different treatments. Enzyme activities were expressed as mean |
Group 2 which received 400 mg kg-1 b.wt. of the crude methanolic extract of P. pellucida without prior induction of oxidative stress via TCE administration, showed the highest SOD activity among the treatment groups (0.187±0.033/min/mg prot).
In Fig. 2b, it can be seen that CAT activity in the rat liver of those treated with crude methanolic extract of P. pellucida (800 and 400 mg kg-1 b.wt.; 0.197±0.019 and 0.369±0.030/min/mg prot, respectively) also showed significant increase when compared with the negative control group (0.026±0.048/min/mg prot). Group 2 showed the highest enzymatic activity of CAT (0.563±0.050/min/mg prot). It is also presented that CAT activity in group 3 is higher than that of group 4. In addition, group 3 showed no significant difference in the activities of both antioxidant enzymes with that of the positive control group (0.377±0.069/min/mg prot).
In the estimation of thioltransferase activity, one of the redox enzymes, the groups treated with P. pellucida extract (800 and 400 mg kg-1 b.wt.; 0.053±0.039 and 0.039±0.025/min/mg prot, respectively) showed significant decrease when compared with the negative control group (0.074±0.068/min/mg prot; shown in Fig. 2c). It was also presented that all the treated groups are significantly different from each other. TT activity is highest in the negative control group which was treated with 1 mL physiologic saline after administration of TCE. Group 4 showed the second highest enzyme activity followed by group 3. TT activity in the positive control group (0.034±0.031/min/mg prot) is significantly higher than that of Group 2 (0.024±0.015/min/mg prot).
Figure 2d shows that the negative control group (0.516±0.409/min/mg prot) has the highest activity of TrxR among the treatment groups followed by group 4 (0.440±0.265/min/mg prot). However, it is also presented that there is no significant difference between the positive control group (0.384±0.391/min/mg prot) and that of group 3 (0.374±0.152/min/mg prot).
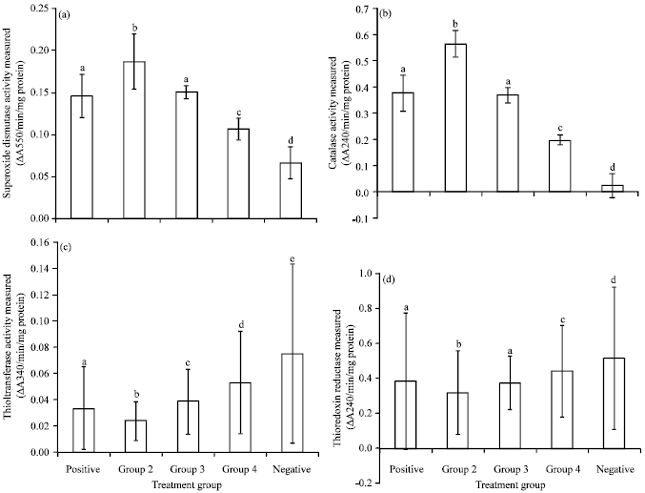 | |
| Fig. 2(a-d): | Estimation of (a) SOD, (b) CAT, (c) TT and (d) TrxR activity in the liver of sprague-dawley male rats under oxidative stress by trichloroethylene for two weeks. Values were expressed as Mean±SEM. Means with different letters on the bar indicate significant difference in enzyme activity based on Tukey’s test at p<0.05 |
Furthermore, group 2 (0.316±0.244/min/mg prot) showed the lowest enzymatic activity of TrxR among the treatment groups.
Histopathological examination: Figure 3 shows the histopathological results of group 1, the positive control group which received 0.07 mL kg-1 b.wt. vitamin E after TCE administration. In the liver samples, the lining of the central vein remained intact and well-defined. The hepatocytes have euchromatic nuclei and were organized into hepatic plates and the sinusoids were not enlarged and devoid of blood (Fig. 3a). Moreover, the hepatic portal triad appeared to be normal (Fig. 3b).
Group 2 as in seen in Fig. 4 was treated with P. pellucida extract at 400 mg kg-1 b.wt. without the administration of TCE. The lining of the central vein is intact but mild fibrosis can be seen around it (Fig. 4a). Moreover, centrolobular hepatocytes displayed ballooning and microvesicularsteatosis (Fig. 4b). A widespread infiltration of blood cells and Kupffer cells was most remarkable in the hepatic sinusoids near the central vein. However, hepatocytes exhibited normal morphology as characterized by euchromaticnuclei and one to two prominent nucleoli (Fig. 4c). In some of the portal veins, the lumen were partially filled with blood cells (Fig. 4d).
Group 3 was treated with 400 mg kg-1 b.wt. P. pellucida extract after the administration of TCE. In this treatment group, the central vein which has collapsed and the hepatic sinusoids which were enlarged, exhibited intense blood congestion (Fig. 5a). In the hepatic portal triad, the portal artery was congested with blood. The hepatic sinusoids were indistinct and filled with erythrocytes (Fig. 5b). Closer analysis revealed that the lining of the central vein has been partially damaged and the hepatocytes have lost their integrity. The cell membranes were torn and the nuclei either fused or overlapped (Fig. 5c).
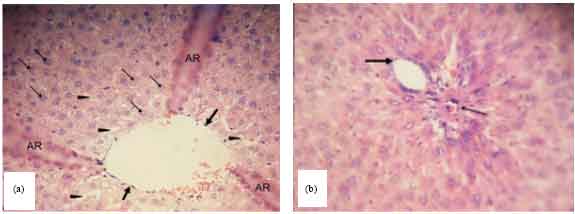 | |
| Fig. 3(a-b): | Transverse section of the rat liver of positive control group, (a) The central vein lining (thick arrow) is well-defined and intact, hepatocytes arranged in hepatic plates showed euchromatic nuclei (thin arrow), and hepatic sinusoids (arrowhead) are not enlarged and devoid of blood. Artifacts (AR). (400x) and (b) The hepatic portal triad appears normal without blood congestion. Portal venule (thick arrow) and arteriole (thin arrow) (1000x) |
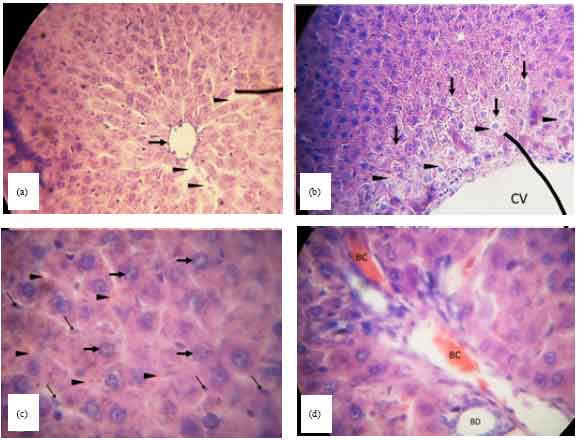 | |
| Fig. 4(a-d): | Light micrograph of a rat liver in group 2 treated with 400 mg kg 1 b.wt .P. pellucida extract, (a) The lining of the central vein is intact but with mild fibrosis (arrow). Some hepatic sinusoids (arrowhead) appear to be slightly enlarged (400x), (b) Ballooning of the centrolobular hepatocytes (arrow). Note that the cells are bigger and lighter due to the presence of microvesicular steatosis (arrowhead). Central vein (CV) (100x), (c) There is widespread appearance of erythrocytes (arrowhead) and Kupffer cells (thin arrow) in the hepatic sinusoids. The nuclei (thick arrow) of the hepatocytes showed normal morphology which is euchromatic and with one or two prominent nucleoli (1000x) and (d) The hepatic portal vein exhibits blood congestion (BC) and bile duct (BD) appears normal (1000x) |
Severe pyknosis and karyorrhexis can also be observed in the said region. Enlarged hepatic sinusoids contained many erythrocytes (Fig. 5d). Hepatocytes surrounding the central vein exhibited an increase in size and foamy appearance which are distinct features of ballooning. Incipient microvesicularsteatosis is also evident as evidenced by the granulated appearance of the cytoplasm. In addition, some centrolobular sinusoids were indistinct and also contained blood cells (Fig. 5e).
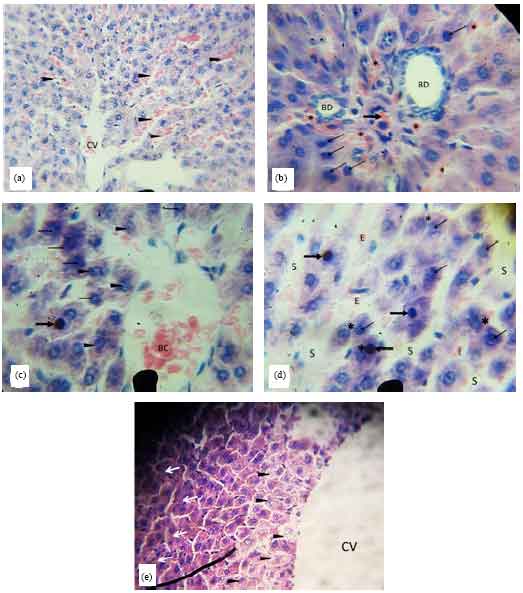 | |
| Fig. 5(a-e): | Photomicrograph of rat liver of group 3 treated with 400 mg kg-1 b.wt. P. pellucida extract, (a) The central vein (CV) appears collapsed and the hepatic sinusoids are enlarged mainly due to blood congestion (arrowhead) (400x), (b) The hepatic portal triad appears normal but the lumen of the hepatic portal artery exhibits blood congestion (thick arrow). Some periportal hepatocytes exhibit mild pyknosis (thin arrow) and the hepatic sinusoids are indistinct and filled with erythrocytes (asterisk) (1000x), (c) The central vein has mild blood congestion (BC) and centrolobular hepatocytes have indistinct cell membrane, irregularly-shaped nucleus (arrowhead) and some exhibits severe pyknosis (thick arrow) and severe karyorrhexis (thin arrow) (1000x), (d) Adjacent centrolobular hepatocytes appear to be fused due to disintegration of cell membrane (asterisk) and some exhibits severe pyknosis (thick arrow) and severe karyorrhexis (thin arrow). Hepatic sinusoids (S) are enlarged and contains some erythrocytes (E) (1000x) and (e) Centrolobular hepatocytes immediately surrounding the central vein exhibit ballooning and microvesicular steatosis (arrowhead). Clusters of hepatocytes distal to the ballooned cells seem to aggregate (arrow) and consequently, the usual hepatic sinusoidal pattern was in disarray. Central vein (CV) (400x) |
Group 4 was treated with 800 mg kg-1 b.wt. P. pellucida extract after the administration of TCE. Liver samples from this treatment group demonstrated massive blood congestion in the central vein and moderate collagen deposition (Fig. 6a). In some areas of the liver, the central vein has collapsed and the surrounding hepatic sinusoids were enlarged due to blood congestion (Fig. 6b). The hepatic portal triad appeared normal but a small portion of the portal vein endothelial lining has been destroyed. Moreover, the portal artery showed slight collagen deposition in its lumen (Fig. 6c).
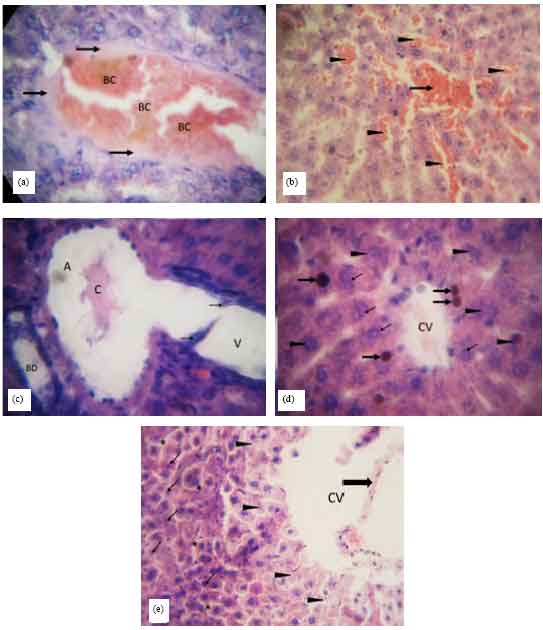 | |
| Fig. 6(a-e): | Photomicrograph of rat liver of group 4 treated with 800 mg kg-1 b.wt. P. pellucida extract, (a) The central vein exhibits massive blood congestion (BC) and moderate amount of collagen (arrow) surround it (1000x), (b) Portion of liver showing a collapsed central vein with massive blood congestion (arrow). All the hepatic sinusoids are markedly enlarged due to blood congestion (arrowhead) (400x), (c) The hepatic portal triad appears normal but small portion of the endothelial lining (arrow) of the portal vein (V) is destroyed. The portal artery (A) shows slight deposition of collagen (C). Bile duct (BD) (1000x), (d) Portion of centrolobular parenchyma showing mild pyknosis (arrow) and karyorrhexis (arrowhead). Adjacent hepatocytes appear to have double or fused nuclei (thin arrow) due to disintegration of cell membranes (1000x) and (e) Marked disintegration of the central vein (CV) as evidenced by the sloughed off lining (thick arrow) in the lumen. Hepatic plates nearest the central vein have undergone ballooning and microvesicular steatosis (arrowhead). Hepatic sinusoids (asterisk) are indistinct and the nuclei of hepatocytes diminished in size (thin arrow) (400x) |
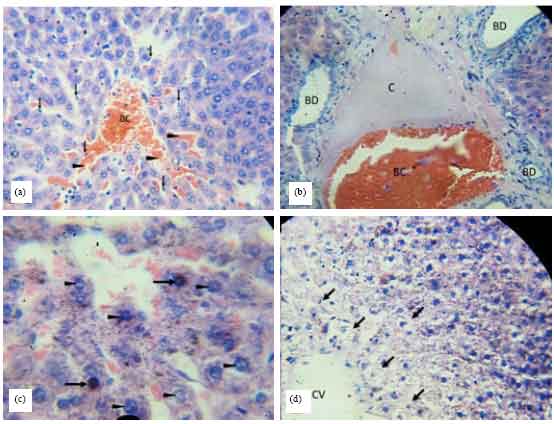 | |
| Fig. 7(a-d): | Photomicrograph of rat liver of the negative control group treated with 1 mL physiologic saline, (a) Central vein appears collapsed, massively congested with blood, and its lining destroyed (double arrow), hepatic sinusoids are enlarged and clogged by blood (arrowhead) with numerous Kupffer cells present in it (arrow) (400x), (b) A hepatic portal triad exhibiting massive blood congestion (BC) and fibrosis (C) in its periphery (1000x), (c) Pyknosis(arrow) in some cells of the liver parenchyma is observable and others with karyorrhexis (arrowhead) (1000x) and (d) Centrolobular hepatocytes exhibit ballooning and microvesicular steatosis (arrow) and extend to regions more distal to the central vein (400x) |
A few hepatocytes were found to have pyknotic nuclei and karyorrhexis was most prevalent in the hepatic plates of the centrolobular area. Most striking is the presence of double or fused nuclei in adjacent hepatocytes with indistinct cellular membranes (Fig. 6d). A marked disintegration of the central vein was noted with the sloughed off lining lying in the lumen. Hepatic plates near the central vein have also undergone ballooning and microvesicularsteatosis. These cells appeared bigger, lighter and frothy in appearance. The hepatic sinusoids seemed to have disappeared yet there are still definite intercellular spaces between adjacent hepatocytes. In effect, no clear cut hepatic plates can be discerned (Fig. 6e).
Group 5 is the negative control group which received physiologic saline treatment after TCE administration. There was a massive congestion of blood in the central vein as their lumen was occluded with erythrocytes. The hepatic sinusoids were enlarged and exhibit mild blood congestion. There was also presence of numerous Kupffer cells in the enlarged sinusoids (Fig. 7a). Likewise, in the hepatic portal triad region, the portal vein exhibited severe blood congestion and intense collagen deposition or fibrosis (Fig. 7b). Pyknosis and karryorhexis of nuclei in hepatocytes were also observed in some regions of the liver (Fig. 7c). Moreover, there was alteration in the hepatocytes even to the most distal areas of the central vein which exhibited high degree of ballooning and microvesicularsteatosis (Fig. 7d).
DISCUSSION
Enzymatic analysis: Oxidative stress is associated with numerous deleterious consequences for the cell, such as lipid peroxidation and even cell death (Wu and Cederbaum, 2003). Various substances such as trichloroethylene can cause formation of free radicals in the form of Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) which will induce oxidative stress to cells (Babre et al., 2010). Trichloroethylene is mainly metabolized in the liver and causes extensive damage to the organ by oxidative stress. It causes increased in liver weights and serum levels of liver enzymes such as SOD and CAT, tissue necrosis and even cell hypertrophy (DEFRAEA, 2004). To be protected from ROS and RNS, living systems are equipped with antioxidant enzymes (SOD, CAT, GPx, LPO etc.) and redox enzymes (TT, TrxR). There are also endogenous antioxidant sources such as alpha-tocopherol and exogenous antioxidant sources which includes flavonoids and xanthones from plants (Babre et al., 2010). Xanthones, specifically xanthone glycosides, have found to have antioxidative property by having inhibitory properties toward lipid peroxidation and radical scavenging properties (Potterat, 1997). In one study, it has been found out that P. pellucida contains a xanthone glycoside called Patuloside A (Khan et al., 2010) which may suggest antioxidative property of the plant.
Administration of the crude extract of P. pellucida increases the activities of both SOD and CAT in TCE-induced liver damage rats to prevent the accumulation of excessive free radicals and protects the liver from TCE intoxication (Kumar et al., 2010). The crude methanolic extract of the plant (400 mg kg-1 b.wt.) also enhanced the activities of SOD and CAT in the liver even without the induction of oxidative stress of trichloroethylene, suggesting that P. pellucida extract does not have deleterious effect on the liver enzymes of the animal. Furthermore, the crude methanolic extract of P. pellucida (800 mg kg-1 b.wt.) showed lower antioxidant enzyme activities than that of group 3 which is in contrast with the study of Mutee et al. (2010), where it was found out that there is increasing antioxidant activity with increasing concentration of P. pellucida. In the study of Mutee et al. (2010), the highest concentration of the methanolic extract of P. pellucida used was 4 mg mL-1 since the determination of DPPH radical scavenging activity is only applicable for low concentrations of a certain plant extract (Sochor et al., 2010). A probable explanation in the decrease of the antioxidant enzyme activities observed in the group treated with 800 mg kg-1 of P. pellucida after TCE induction is the presence of apiol, mainly dill-apiol, in the P. pellucida extract (Xu et al., 2006). Apiols are present in some plants of the families Piperaceae and Apiaceae and several studies have shown that these substances may cause DNA binding activity in rats and mice which eventually leads to severe damage in liver and presence of liver tumors (Phillips et al., 1984; Miller et al., 1983). In a study of Bayma et al. (2000), dill-apiol compound was isolated from the aerial parts of P. pellucida, however, the percentage composition of this compound in P. pellucida extract and have not yet been studied. Likewise, mutagenicity and toxicity studies of this compound have not yet been reported. In a study made by Vora et al. (2009), Petroselinum crispum extract (2, 5 and 10 mg kg-1 b.wt.) which contained apiols, showed protective activities in the brain of mice. However, Buchanan (1978) has reported that 50 and 200 mg kg-1 body weight of P. crispum cause severe liver damage and even death to mice. These studies may be indicative of the lowered antioxidant enzyme activities of a higher concentration of P. pellucida.
Redox enzymes which include TT and TrxR, play an important role at diverse levels of cellular function, including the stress response and cell growth. Both redox enzymes in study (TrxR and TT) are small disulfide reducing enzymes that have conserved consensus sequence-CXXC- at their active sites (Kim et al., 2003). In both enzymes, the negative control group which were administered with trichloroethylene showed the highest enzyme activities of both TT and TrxR, suggesting increased oxidative stress state in the tissue. The crude methanolic extract of the plant (800 and 400 mg kg-1 b.wt.) lowered the enzyme activities of both redox enzymes compared to that of the negative control group. Group 3 which received 400 mg kg-1 b.wt. of crude methanolic extract of P. pellucida after administration of trichloroethylene, lowered the enzyme activity levels of both TT and TrxR as compared with that of the negative control group and the group 4 (800 mg kg-1 b.wt.). This is also in conjunction with the results of the antioxidant enzymes, where 800 mg kg-1 b.wt. of P. pellucida extract presented lower antioxidant activity than that of a lower concentration. Again, the presence of apiols in the extract (Xu et al., 2006; Bayma et al., 2000), may account for the lesser protective effect of a higher concentration of P. pellucid extract (Buchanan, 1978), since the said compound has been proven to mediate DNA binding activity in rats and mice which eventually leads to severe damage in liver and presence of liver tumors (Phillips et al., 1984; Miller et al., 1983). In addition, group 2 which only received 400 mg kg-1 b.wt. of P. pellucida without any administration of trichloroethylene, showed the least activity levels for both redox enzymes, suggesting that P. pellucida extract did not cause any harmful effects to the liver enzymes measured.
Histopathological examination: Normal liver architecture is characterized by the distinct lobulation of certain hepatic areas. The hepatic sinusoids radiate out from an intact and well-defined central vein. Hepatocytes are polygonal, compact and arranged in plates. They are distinct, well-stained and with a prominent single nuclei or polynuclei. The hepatic portal triad consisting of the small bile duct, hepatic artery and hepatic vein occupies a small triangular area surrounded by thin connective tissues and confluent with the hepatic sinusoids (Atangwho et al., 2010; Fawcett and Raviola, 1994). However, the liver is prone to oxidative injury because of its central role in combatting free radicals and other reactive species and its portal location within the circulatory system. TCE, a known inducer of oxidative stress, will generate free radicals upon absorption in the body and will react with the components of the cells (Aslan et al., 2000; Gharib, 2009).
Group 1 exhibited absence of or minimal damage in the liver samples. The said condition can be attributed to the oral administration of vitamin E. According to Wazir et al. (2008), necroinflammation of the liver is significantly decreased as opposed to fibrosis which can only be prevented by the action of vitamin E. In a study conducted by Yazar et al. (2004), it was found out that treatment with tocopherols hindered the occurrence of oxidative stress. Vitamin E directly reduced membrane peroxides by promoting the termination phase during radical reactions and eventually facilitating the local action of membrane glutathione. Because ROS were rapidly and efficiently reduced or less produced, they were not able to accumulate and cause a chain of deleterious events for the cell. Basically, antioxidants reduce the hepatic injury caused by oxidative stress by scavenging free radicals responsible for cell damage, enhancing enzymes responsible for antioxidant activity and inducing the regeneration of liver cells (Ao et al., 2009). The aforementioned mechanism of action of Vitamin E most likely provided protection to the hepatic tissue and its individual constituents from the damage caused by free radicals during oxidative stress induction.
Group 2 was treated with the plant extract without prior administration of TCE. The liver samples from this group accumulated slight hepatic injuries which include mild fibrosis, minor blood congestion in the sinusoids and portal vein, ballooning and microvesicularsteatosis in 2-3 layers of centrolobular hepatocytes. The said damages may have been caused by the innate sensitivity of liver from foreign compounds introduced into the system (Foxal, 2008). This may also be supported by the infiltration of numerous Kupffer cells in the hepatic sinusoids. Kupffer cells are the resident specialized macrophages of the liver and are responsible in breaking down aged erythrocytes and eliminating foreign materials in the said organ (Fawcett and Raviola, 1994; Mescher, 2010). Their main role is to remove bacteria and foreign proteins from the blood to ensure normal functioning of the liver in removing toxic substances. Generally, Kupffer cells are activated by numerous molecules including chemical substances and bacterial endotoxins (Wheeler, 2003). The Kupffer cells from this group most probably detected TCE, thus activating them and promoting their rapid migration and proliferation near the central vein to eliminate the foreign substance. This phenomenon was also observed in previous studies regarding Kupffer cell mobilization in mice. Mouse Kuppfer cells prelabeled with carbon or iron were found out to migrate to the necrotic centrilobular zone following a carbon tetrachloride-induced necrosis in the liver after 48 h of administration (Parry, 1978). Moreover, in another study, rats intoxicated with carbon tetrachloride showed Kupffer cell hyperplasia or abnormal cell proliferation. Kupffer cell number per mm3 of normal rat liver is about 16,500. In the investigation, when the hepatocytes started to exhibit necrosis 48 h after the administration of the chemical, Kupffer cells proliferated 5.2 times in the central zone and 1.7 times in the periportal zone (Kiyotoshi, 1982).
For group 3, liver samples had a more extensive degree of inflammation. Massive blood congestion was observed in the central and some portal veins which may be attributed to the destruction of the valves. The flow of blood may have been disrupted and consequently, aggregation of erythrocytes occurred. Fibrosis which was present in moderate amount, may also have played a factor in the congestion of blood in this group. Fibrosis is the accumulation of excessive connective tissue or overproduction of extracellular matrix as a mechanism for tissue healing. This process is usually triggered if there is a chronic injury caused by an inflammatory factor such as TCE. Moreover, fibrosis can lead to portal hypertension and the scarring may have distorted blood flow in the liver (Shaffer, 2007). The hepatic sinusoids were enlarged and filled with blood. This event possibly occurred due to the extravasation or overflow of erythrocytes from the occluded central vein. Due to blood congestion, an increase in venous pressure happens in the central vein and that pressure is transmitted to the sinusoids which subsequently dilate (Rubin and Reisner, 2009). As a result, blood cells from the central vein were able to leak to the enlarged hepatic sinusoids. Hepatocyte ballooning and microvesicularsteatosis were also evident in the centrolobular region of the central veins. Hepatocyte ballooning is a structural manifestation of severe cell injury and is likely demonstated by cells undergoing lytic necrosis. It is characterized by the swelling or enlargement of the hepatocytes categorized as a rarefaction of the cytoplasm. Ballooned hepatocytes are located most often in the centrilobular parenchyma and interspersed with regions exhibiting steatosis (Schattner and Knobler, 2008). On the other hand, microvesicularsteatosis is the abnormal accumulation of fat wherein the nucleus has not been fully displaced and reflects an impaired hepatocellular metabolism (Goldfrank, 2006). It can also be deduced that hepatocytes from this group are in the process of or a visible manifestation of necrosis due to pyknosis and karyorrhexis. Pyknosis is a characteristic of a dying cell’s nucleus which exhibits a shrivelled and intensely basophilic appearance while karyorrhexis is the disintegration of the nucleus into small, darkly-staining fragments which may occur de novo or succeed pyknosis (Jones et al., 1997; Cormack, 2001). The appearance of pyknosis in the liver of chemically-treated rats was also found in studies made by Chanchal and Das (2010) and Kanchana and Sadiq (2011). Paracetamol and carbon tetrachloride were the substances utilized in the experiements, respectively. Histological analysis of their liver samples displayed pyknosis and necrosis in groups treated with the particular chemical they used, suggesting that such compounds are capable of damaging hepatic tissues.
The said damages on group 3 samples could be attributed to the initial reaction of the liver upon the introduction of an oxidative stress inducer such as TCE. In a case study in humans by McCunney (1988), liver function abnormalities such as reactive hepatitis and fatty liver which developed insidiously were observed from a group of chemical workers exposed to TCE. It was suggested that the hepatic injury was mostly due to the direct toxic effect of the chemical substance or the hypersensitivity of the liver. Moreover, trichloroethylene exposure by ingestion of laboratory animals caused notable hepatic effects such as increase in serum levels of liver enzymes, necrosis and cell hypertrophy (Foxal, 2008). Similar to Group 3 and 4 liver samples also exhibited blood congestions but was only intense in some central veins. Their linings were partially damaged as opposed to group 4 wherein there was total destruction of the central vein lining. The hepatic plates from this group were still arranged in cords while those from group 4 were in disarray. Ballooning and microvesicularsteatosis were present in both groups but the extent of these damages was confined to only a few layers of the centrolobular hepatocytes. Although both groups received treatment of the plant extract, group 4 exhibited less hepatic damage than group 4. Observations from these two groups are supported by scientific studies involving apiols which is a compound of P. pellucida (Xu et al., 2006). Apiols given in high frequent doses can be toxic not only in the liver, but in kidneys as well. Lesions found on the affected organs are lethal but variations in susceptibility are widely demonstrated (Patoir and Patoir, 1936). Generally, a direct relationship is associated with the concentration of the plant extract and the hepatic damages which could occur. When higher concentration of P. pellucida extract was used, a more injured liver was observed. Since, group 3 received a lower dosage than group 4, a lesser extent of damage was noted.
Group 5, the negative control group, exhibited the most severe damage in all the liver samples. Chronic exposure to TCE resulted to several hepatic injuries that led to the impairment of normal liver functions (Zimmerman, 1999; Provaznik and Kizek, 2001; Kuntz and Kuntz, 2008). Auspiciously, harmful damages of the resulting free radicals can be hindered or reversed with the administration of an antioxidant. The antioxidants render the free radicals harmless and stop the chain reaction formation of new free radicals (Barron, 2008; Blake, 2008; Rolfes et al., 2009). However, in group 5, P. pellucida extract or vitamin E exhibiting antioxidative property were not given to the test subjects, thus a fatty liver due to microvesicularsteatosis and a high probability of necrosis.
A comparison between group 4 and 5 showed that the latter has a higher degree of damage than the former by taking into consideration the injuries observed in the liver samples. The blood congestion in the central vein in group 5 was more widespread than those in group 4. Ballooning and microvesicularsteatosis were observed in both groups but these conditions are even more pronounced in group 5 since regions distal the centrolobular hepatocytes were also affected. Moreover, pyknosis and karyorrhexis were more apparent in group 5 than in 4. Less hepatic injuries observed in group 3 can be attributed to the treatment of the P. pellucida extract which may have hindered or reversed the deleterious effects caused by ROS from TCE administration. This finding can be supported by a study made by Mutee et al. (2010) wherein the methanol extract of P. pellucida has been proven to demonstrate DPPH free radical scavenging activity. With that, less damage is definitely expected from group 4.
Based on the statistical analysis for necroinflammatory grade, groups 1-3 have been categorized into one group by not exhibiting a significantly different p-value. This suggests that administration of the plant extract at 400 mg kg-1 b.wt. has a similar effect to that of the established antioxidant, Vitamin E, on rat liver with or without oxidative stress. For the fibrosis stage of the liver, groups 1-4 showed statistically insignificant results suggesting that P. pellucida renders no toxic effect on biological system of the test animal and has the potential to boost antioxidant and redox enzymes in liver.
It is recommended that future studies may be directed in extracting the active compound of the plant extract and its corresponding physical effects on organisms. In addition, it is suggested that further studies utilizing more concentrations of the plant extract in testing its effects on the antioxidant and redox enzymes. Moreover, the extract could also be used to test its effects on other organs of the body such as kidneys, brain, lungs, heart and gonads. The behavioural adaptations of the rats subjected to oxidative stress could also be dealt with in future investigations.
REFERENCES
- Ao, Z.H., Z.H. Xu, Z.M. Lu, H.Y. Xu, X.M. Zhang and W.F. Dou, 2009. Niuchangchih (Antrodia camphorate) and its potential in treating liver diseases. J. Ethnopharmacol., 121: 194-212.
CrossRefDirect Link - Arrigoni-Blank, D.F.M., E.G. Dmitrieva, E.M. Franzotti, A.R. Antoniolli, M.R. Andrade and M. Marchioro, 2004. Anti-inflammatory and analgesic activity of Peperomia pellucida (L.) HBK (Piperaceae). J. Ethnopharmacol., 91: 215-218.
PubMed - Aslan, R., M. Tutuncu, S. Aslan and Y. Dundar, 2000. Acute phase effect of trichloroethylene ingestion on some biological markers in dogs. Turk J. Vet. Anim. Sci., 24: 109-112.
Direct Link - Avti, P.K., S. Kumar, C.M. Pathak, K. Vaiphei and K.L. Khanduja, 2006. Smokeless tobacco impairs the antioxidant defense in liver, lung and kidney of rats. Toxicol. Sci., 89: 547-553.
CrossRefDirect Link - Babre, N., S. Debnath, Y.S. Manjunath, P. Parameshwar, S.V. Wankhede and K. Hariprasath, 2010. Antioxidant potential of hydroalcoholic extract of Bariingtoniaacutangula Linn roots on streptozotocin-induced diabetic rats. Int. J. Pharm. Pharm. Sci., 2: 201-203.
Direct Link - Bayma, J.D.C., M.S.P. Arruda, A.H. Muller, A.C. Arruda and W.C. Canto, 2000. A dimeric ArC2 compound from Peperomia pellucida. Phytochemistry, 55: 779-782.
CrossRefDirect Link - Brunt, E.M., 2000. Grading and staging the histopathological lesions of chronic hepatitis: The knodell histology index and beyond. Hepatology, 31: 241-246.
CrossRefDirect Link - Buchanan, R.L., 1978. Toxicity of spices containing methylenedioxybenzene derivatives: A review. J. Food Safety, 1: 275-293.
CrossRefDirect Link - Gharib, O.A., 2009. Effects of Kombucha on oxidative stress induced nephrotoxicity in rats. Chin. Med., Vol. 4.
CrossRef - Kim, H.G., S.M. Hong, S.J. Kim, H.J. Park and H.I. Jung et al., 2003. Age-related changes in the activity of antioxidant and redox enzymes in rats. Mol. Cells, 16: 278-284.
PubMedDirect Link - Kanchana, N. and A.M. Sadiq, 2011. Hepatoprotective effect of Plumbagozeylanica on paracetamol-induced liver toxicity in rats. Int. J. Pharm. Pharm. Sci., 3: 151-154.
Direct Link - Kiyotoshi, S., 1982. Kupffer cell hyperplasia in rats intoxicated by carbon tetrachloride as demonstrated by scanning electron microscopy. J. Gastroenterol., 17: 422-429.
Direct Link - Kumar, R.S., K.A. Kumar and N.V. Murthy, 2010. Hepatoprotective and antioxidant effects of caesalpiniabonducella on carbon tetrachloride-induced liver injury in rats. Int. Res. J. Plant Sci., 1: 62-68.
Direct Link - McCunney, R.J., 1988. Diverse manifestations of trichloroethylene. Br. J. Ind. Med., 45: 122-126.
CrossRefDirect Link - Miller, E.C., A.B. Swanson, D.H. Phillips, L. Fletcher, A. Liem and J.A. Miller, 1983. Structure-activity studies of the carcinogenicities in the mouse and rat of some naturally occurring and synthetic alkenylbenzene derivatives related to safrole and estragole. Cancer Res., 43: 1124-1134.
Direct Link - Mutee, A.F., S.M. Salhimi, M.F. Yam, C.P. Lim and G.Z. Abdullah et al., 2010. In vivo anti-inflammatory and in vitro antioxidant activities of Peperomia pellucida. Int. J. Pharmacol., 6: 686-690.
CrossRefDirect Link - Pavlovic, V., D. Pavlovic, G. Kocic, D. Sokolovic, M. Sarac and Z. Jovic, 2009. Ascorbic acid modulates monosodium glutamate induced cytotoxicity in rat thymus. Bratisl Lek Listy, 110: 205-209.
PubMedDirect Link - Parry, E.W., 1978. Studies on mobilization of Kupffer cells in mice: I. The effect of carbon tetrachloride-induced liver necrosis. J. Comp. Pathol., 88: 481-487.
CrossRefDirect Link - Sochor, J., O. Zitka, H. Shutkova, D. Pavlik and P. Babula et al., 2010. Content of phenolic compounds and antioxidant capacity in fruits of apricot genotypes. Molecules, 15: 6285-6305.
CrossRefDirect Link - Vora, S.R., R.B. Patil and M.M. Pillai, 2009. Protective effects of Petroselinum crispum (Mill) Nyman ex A. W. Hill leaf extract on D-galactose-induced oxidative stress in mouse brain. Indian J. Exp. Biol., 47: 338-342.
Direct Link - Wazir, I., F. Wazir, M. Javed, M. Saeed, N. Haq and H. Khan, 2008. Effect of vitamin E supplements in therapy of chronic hepatitis C: A histological study. Gomal J. Med. Sci., 6: 81-86.
Direct Link - Wheeler, M.D., 2003. Endotoxin and Kupffer cell activation in alcoholic liver disease. Alcohol Res. Health, 27: 300-306.
Direct Link - Wu, D. and A.I. Cederbaum, 2003. Alcohol, oxidative stress and free radical damage. Alcohol Res. Health, 27: 277-284.
Direct Link - Xu, S., N. Li, M.M. Ning, C.H. Zhou, Q.R. Yang and M.W. Wang, 2006. Bioactive compounds from peperomia pellucida. J. Nat. Prod., 69: 247-250.
CrossRefDirect Link - Yazar, E., S. Konyalioglu, R. Col, Y.O. Birdane, A.L. Bas and M. Elmas, 2004. Effects of vitamin E and prednisolone on some oxidative stress markers in endotoxemic rabbits. Revue Med. Vet., 155: 538-542.
Direct Link