
G. Iyyapparajanarasimapallavan
Center of Advanced Study in Marine Biology, Annamalai University, Portonovo-608 502, Tamil Nadu, India
P. Sampath Kumar
Center of Advanced Study in Marine Biology, Annamalai University, Portonovo-608 502, Tamil Nadu, India
C. Prasanna Kumar
Center of Advanced Study in Marine Biology, Annamalai University, Portonovo-608 502, Tamil Nadu, India
K.C.A. Jalal
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
B.Y. Kamaruzzaman
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
B. Akbar John
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 1 | Page No.: 18-25







ABSTRACT
The distribution and abundance of the gelatinous zooplankton namely hydromedusae, scyphomedusae, siphonophore, ctenophore, salps, doliolids, appendicularians and chaetognaths were investigated in two seasons at seven stations in East Tamil Nadu coast, India. The occurrences of gelatinous zooplankton were observed to be more in summer than in winter. However, the composition of different species in each group varied from all the stations and seasons. Apart from the individual variation in seasonal distribution, most of the species appeared to be cosmopolitan in the Indian waters. A total of 34 species from summer and 17 species from winter were recorded during the present study. The hydromedusae were dominant in both the seasons of the study period. Ephyrae of the scyphomedusae were also noticed in both the seasons.
PDF Abstract XML References Citation
Received: October 01, 2012;
Accepted: October 20, 2012;
Published: February 09, 2013
How to cite this article
G. Iyyapparajanarasimapallavan, P. Sampath Kumar, C. Prasanna Kumar, K.C.A. Jalal, B.Y. Kamaruzzaman and B. Akbar John, 2013. Distribution and Abundance of Gelatinous Zooplankton along Tamil Nadu Coastal Waters. Journal of Biological Sciences, 13: 18-25.
DOI: 10.3923/jbs.2013.18.25
URL: https://scialert.net/abstract/?doi=jbs.2013.18.25
DOI: 10.3923/jbs.2013.18.25
URL: https://scialert.net/abstract/?doi=jbs.2013.18.25
INTRODUCTION
Gelatinous zooplankton are taxonomically diverse group of macroplankton playing vital role in structuring coastal and estuarine ecosystems (Mahboob and Zahid, 2002; Ali et al., 2003; Guher et al., 2004; Suchman and Brodeur, 2005), whose diversity is less explored due to their fragile nature (Raskoff et al., 2003). They include medusae, siphonophores, ctenophores, chaetognaths, pteropods, heteropods, appendicularians, salps, doliolids and pyrosomes (Ahmed et al., 2007; Oueda et al., 2007). Gelatinous zooplankton are widely distributed in all the major ocean waters. However, they are the least understood of all planktonic animal groups (Raskoff et al., 2003; Saravanakumar et al., 2007; El-Sherbiny et al., 2007; Rahimibashar et al., 2009; Kumar and Perumal, 2011). They occur in all the oceans from the surface to the seafloor. In addition of being beautiful, these drifters are capable of eating enormous amounts of prey and may compete with fishes for these natural resources. They form an important integral part of the ecosystem, the notable gap in understanding the linkages between primary and secondary productivity in the oceans being the lack of knowledge about gelatinous zooplankton (Raskoff et al., 2005). In recent years, strong and sustained increments of gelatinous organism populations have been recorded in different geographic marine areas (Mills, 2001; Brodeur et al., 2002). These proliferations have caused important changes in the pelagic community structure due to the impact of gelatinous organisms as consumers and competitors of zooplankton and fish larvae (Matsakis and Conover, 1991; Purcell, 1997). Abundance of gelatinous zooplankton has been implicated in phytoplankton blooms throughout trophic cascading (Lindahl and Hernroth, 1983; Verity and Smetacek, 1996; Schneider and Behrends, 1998). They are also able to maintain feeding rates proportional to food concentration over a wide range and allowing them to efficiently exploit high prey densities (Bamstedt, 1990, 1998; Sornes and Aksnes, 2004). These life history characteristics may at times result in a competitive edge in resource utilization with respect to each other and slower growing consumers of zooplankton e.g., many crustaceans and fishes. It also contributes to the naturally pulsed occurrence of many species of gelatinous predators in the plankton as well as the extraordinary blooms of jellies frequently observed (Arai, 1992; Boero et al., 2001). Among these groups, medusae, siphonophores and ctenophores are very abundant and dense aggregates have been recorded in various coastal settings of the world (Palma and Rosales, 1995; Palma and Apablaza, 2004). The sudden appearance of medusa with an apparently “abnormal” abundance of one or a few species (medusa blooms) is a common but as yet enigmatic characteristic of gelatinous plankton life histories. These events are usually noticed and reported when massive appearances of conspicuous stinging jellyfish occur near the coastal zone significantly and visibly affecting human activities such as tourism and fisheries (Haddad et al., 2002). The majority of the published records on gelatinous zooplankton are from the other part of the countries. Until recently, there were only few records on gelatinous zooplankton along Tamil Nadu coastal waters. The present study reports on the occurrence and abundance of gelatinous zooplankton along Tamil Nadu coastal waters.
MATERIALS AND METHODS
Sampling sites: The Bay of Bengal is located in the eastern side of India which is a tropical basin and experiences monsoonal wind force that reverses its direction semi-annually. A large number of rivers such as the Irrawaddy, Brahmaputra, Ganges, Godavari, Krishna and Cauvery discharge into the Bay of Bengal. The present study has been carried out in seven different coastal environmental stations along Tamil Nadu coastal waters from station 1 (8°7'N, 77°33'E) to station 7 (13°06'N, 80°17'E) (Table 1) and the map showing the study area is depicted in Fig. 1. The investigations were carried out on two different seasons during summer and winter 2008.
Sampling procedure: The samples were collected using plankton net with a mesh size of 200 μm and 50 cm diameter. After the net operations, the gelatinous organisms were fixed in 5% formalin solution in seawater for subsequent laboratory analysis.
| Table 1: | GPS locations of the sampling stations |
 | |
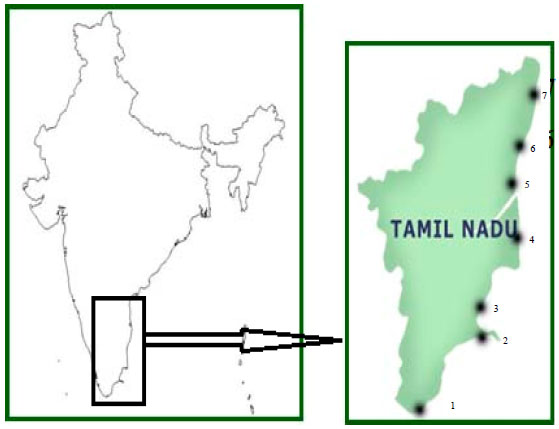 | |
| Fig. 1: | Map of the study area, 1: Kanyakumari, 2: Mandapam, 3: Thondi, 4: Nagapattinam, 5: Parangipettai, 6: Puducherry, 7: Chennai |
The gelatinous zooplankton collection were made in different ways, not only by the plankton net and also by using scoop net, hand pick, because of their size variation from 2 mm to 20 cm, especially Scyphomedusae group.
Identification key: The Identification of gelatinous zooplankton was based on the taxonomic keys described by various authors (Kirkpatrick and Pugh, 1984; Kramp, 1959, 1961; Russell, 1953, 1970).
Data analysis: Biodiversity indices were calculated by following the standard formula of Shannon and Weaver (1949) diversity index (H’), richness (D); Pielou (1966) evenness. Pearson correlation co-efficient (r) values were calculated to understand the relationships between distribution, diversity of Gelatinous zooplankton and hydrographical features.
RESULTS
Species composition and abundance of gelatinous zooplankton: A total of eight groups of gelatinous zooplankton were observed during the study period. A total of 14 species of hydromedusae in summer and 5 species in winter were recorded, among this, the maximum number of species were recorded from station 1 whereas, the minimum were from stations such as station 2, 3 and 7. In winter, the maximum numbers of species were recorded from station 5 whereas, the minimum were from station 6 and 7. A total of 5 species of Scyphomedusae in summer and 3 species from winter were recorded. Among this the maximum numbers of species were recorded from station 5 whereas, the minimum were from station 3 and 6. The maximum numbers of species were recorded from station 5, whereas the minimum were from station 2. A total of 5 species of Siphonophores in summer and 2 species from winter were recorded. The maximum numbers of species were recorded from station 5 whereas the minimum were from station 1 and 7. In winter, the maximum numbers of species were recorded from station 5 whereas the minimum were from station 3. Only one species of Ctenophores in both the seasons was recorded, the Pleurobrachia sp. was observed in this group. This was recorded from station 1 and 5 whereas no species of this group were recorded from station 2, 3, 4, 6 and 7. A total of 3 species from the group of salps and doliolids in summer and 3 species from winter were recorded. The maximum numbers of species of this group were recorded from station 5 whereas the minimum was from station 3. In winter, the maximum number of species of this group was recorded from station 3 whereas the minimum was from station 2. A total of 2 species of Appendicularians in summer and 2 species from winter were recorded. The Oikopleura species were observed in both the seasons. In summer the maximum numbers of species of this group were recorded from station 5 whereas the minimum was from station 6. In winter, the maximum number of species of this group was recorded from station 6 whereas the minimum was from station 2 and 3. A total of 3 species of Chaetognaths from summer and 2 species from winter were recorded. The maximum numbers of species were recorded from station 5 whereas the minimum was from station 2 and station 7. In winter, the maximum number of species were recorded from station 3 where as the minimum were from station 2 and station 7. The details of species composition and abundance of gelatinous zooplankton are depicted Table 2-5, respectively.
Diversity indices of gelatinous zooplankton: The detailed values of the different diversity indices are depicted in the Table 4 and 5. The species diversity ranged from 2.174 to 2.742 during summer and 1.274 to 2.277 during winter. The minimum species diversity was recorded during winter at station 3 and the maximum species diversity was recorded during summer at station 1. The species richness ranged from 0.8193 to 0.9105 during summer and 0.6786 to 0.8889 during winter. The minimum species diversity was recorded during winter at station 3 and the maximum species diversity was recorded during summer at station 2. The species richness ranged from 0.7843 to 0.9249 during summer and 0.7913 to 0.9496 during winter. The minimum species diversity was recorded during summer at station 4 and the maximum species diversity was recorded during winter at station 5.
Correlation between different physicochemical parameters and gelatinous zooplankton diversity: The present study found that the diversity and abundance of the gelatinous zooplankton are highly influenced by the various physicochemical parameters prevailing in the study area. There was positive correlation between the abundance of gelatinous zooplankton and salinity, temperature and pH of the study area, whereas there was a negative correlation between dissolved oxygen and abundance of gelatinous zooplankton. This trend was similar in both the summer and winter seasons (Table 6).
| Table 2: | Distribution and abundance of gelatinous zooplankton in summer |
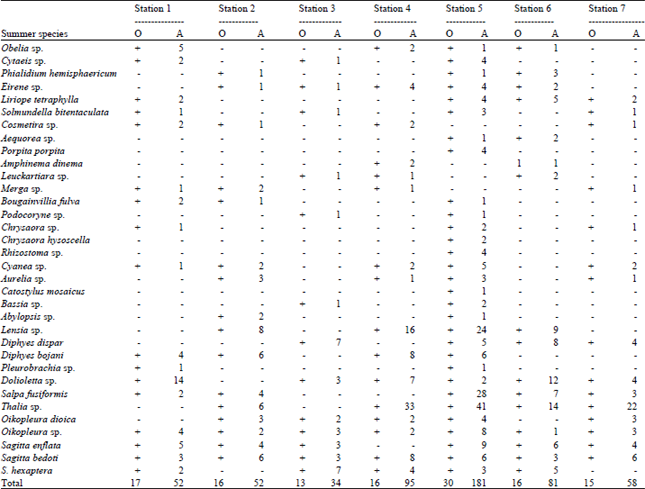 | |
| ES: Eudoxid stage, EL: Ephyra larvae, O: Occurrence, A: Abundance (ind L-1), Stations 1: Kanyakumari, 2: Mandapam, 3: Thondi, 4: Nagapattinam, 5: Parangipettai, 6: Puducherry, 7: Chennai | |
| Table 3: | Distribution and abundance of gelatinous zooplankton in winter |
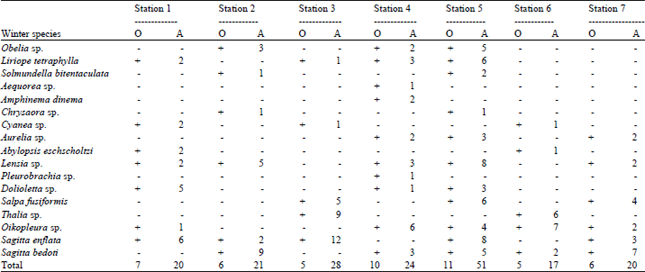 | |
| ES: Eudoxid stage, EL: Ephyra larvae, O: Occurrence, A: Abundance (ind L-1), 1: Kanyakumari, 2: Mandapam, 3: Thondi, 4: Nagapattinam, 5: Parangipettai, 6: Puducherry, 7: Chennai | |
| Table 4: | Diversity indices of gelatinous zooplankton during summer |
 | |
| 1: Kanyakumari, 2: Mandapam, 3: Thondi, 4: Nagapattinam, 5: Parangipettai, 6: Puducherry, 7: Chennai | |
| Table 5: | Diversity indices of gelatinous zooplankton during winter |
 | |
| 1: Kanyakumari, 2: Mandapam, 3: Thondi, 4: Nagapattinam, 5: Parangipettai, 6: Puducherry, 7: Chennai | |
| Table 6: | Correlation matrix between various parameters and diversity and abundance of zooplankton |
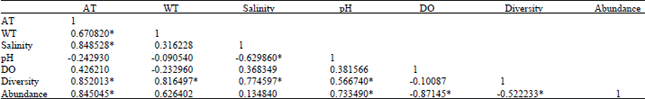 | |
| The results are pooled values of the total data obtained, n = 10, *Significant at p<0.05, AT: Atmospheric temperature, WT: Water temperature, DO: Dissolved oxygen | |
DISCUSSION
There were few records on whole gelatinous zooplankton from the study area, in this connection the present study were conducted and a total of eight groups of gelatinous zooplankton were observed during the study period. They are hydromedusae, scyphomedusae, siphonophores, ctenophores, salps, doliolids, appendicularians and chaetognaths in both the seasons. However, the composition of different species in each group varied from all the stations and seasons. The seasonal distribution and abundance of planktonic cnidarians is primarily governed by factors controlling the release of the larvae. Temperature, food abundance, salinity and the ratio of light to dark are known to affect the liberation of medusae from hydroids (Arai, 1992). Apart from the individual variation in seasonal distribution, most of the species appeared to be cosmopolitan in the Indian waters. Hydromedusae are important in the sense that they are exclusively carnivores and moreover, they are predators and hence they compete with other predators like fish larvae (Santhakumari and Nair, 1999). This group often serves as an index to industrial pollution (Santhakumari et al., 1999). Among the group hydromedusae, Liriope tetraphylla occupied the dominant position and more in summer. The similar observations were noticed by Zakaria (2004), the density of hydromedusae was high in the inshore waters during summer and in the east coast of India by Santhakumari and Nair (1999). In the group of scyphomedusae, minimum species were got and those were picked by hand, because they are maximum in size when compare to that of the other gelatinous zooplankton and the samples are collected from the shorewards and some larvae stages were also noticed from the present collection from the sampling areas. Suchman and Brodeur (2005) observed that the genus Chrysaora was always most dense at the most shoreward. The cosmopolitan scyphomedusa Aurelia aurita is particularly abundant during summer and it is recognized as an important predator in plankton communities (Hay et al., 1990). In the present investigation the Aurelia sp. were observed as dominant during the summer. The same report has pointed out that scyphomedusae A. aurita was the most abundant and commonly occurred during the summer months (Ballard and Myers, 2000). Siphonophora are abundant in the Indian seas and constitute an important part of the marine plankton (Venkataraman, 2005). The dominant species from siphonophores are the Lensia species. In contrast to just one predominantly epipelagic species, Solmundella bitentaculata, these high abundances observed in the upper 100 m were mostly due to large numbers of Lensia sp. (Hosia et al., 2008). In the present investigation, the Solmundella bitentaculata were present from the sample collected from seven stations and (Zakaria, 2004), observed that the siphonophore density was high in summer and in the present observation also the average density in summer was high, among this the Lensia sp. were observed as dominant in both the seasons. Comb jellies comprise a diverse phylum (ctenophore) of delicate, gelatinous species living throughout the world’s oceans (Purcell et al., 2001). The phylum Ctenophora (known as comb jellies) consists of gelatinous marine carnivores found from the surface to several thousand meters depth. Ctenophores are exclusively carnivorous, their principal feeding task is the capture of prey; there are no herbivorous ctenophores and only one genus can sometimes be parasitic (Haddock, 2007). Ctenophores have been known to occur throughout the year along the east coast of India. Pleurobrachia sp. occurred during the summer and reached a maximum density (Ballard and Myers, 2000). In the present study also state the same as above and only one species of Pleurobrachia sp. were identified from this group. Tunicates were subdivided into two groups: appendicularians encompassing fritillarians and oikopleurids and thaliaceans which included both doliolids and salps, salps being numerically dominant (Stemmann et al., 2008). Salpa fusiformis is usually the predominant gelatinous organism in the group of salps, Occurrence of high density of the alternation of generation of Salpa fusiformis in this study suggested that the animals could attain rapid population increase through the ontogenic cycle (Chae et al., 2008). This species has the widest world distribution of all salps (Pages et al., 2001). In the present study the Salpa fusiformis was also dominantly represented from this group. The blooming also has been usually observed during spring and summer when the phytoplankton is productive (Harris et al., 1991; Menard et al., 1994). Generation of salps may be achieved when enough food sources are provided to these animals and the growth rates are extremely high (Laval, 1997). They can utilize a broad prey size spectrum and are therefore assumed to be of special importance in oligotrophic environments as the most competitive metazooplankton group to survive and live at low food concentrations (Fenaux et al., 1998; Acuna, 2001). Larvaceans can contribute significantly to planktonic biomass (Clarke and Roff, 1990; Uye and Ichino, 1995; Hopcroft et al., 1998) but because they rarely dominate numerically, they are generally believed to be minor components of the zooplankton community (Hopcroft et al., 1998). It is interesting to note that the genus Oikopleura which contains the heaviest species showed significant relationships to both small phytoplankton and Larvacean biomass (Hopcroft et al., 1998). In the present study the Oikopleura species were present throughout the collection and the summer distribution is higher than in the winter, the same report were noticed by Santhanam and Perumal (2003). The abundance and community structure of chaetognaths in the northern Indian Ocean was investigated based on zooplankton samples (Nair et al., 2002). Sagitta enflata or Flacisagitta enflata, is a cosmopolitan species in temperate and warm waters and occurs mainly in the upper 300 m (Pierrot-Bults and Nair, 1991). It is the dominant chaetognatha species by number in the Indian Ocean (Andreu et al., 1989; Michel, 1995). In the present investigation the most dominant species in the chaetognatha is the Sagitta enflata as in the case of earlier studies.
CONCLUSION
There was significant variation in atmospheric temperature, water temperature, salinity, pH and dissolved oxygen between different sampling stations during summer and winter (p<0.05). A total of eight groups of gelatinous zooplankton were observed during the present study. They are hydromedusae, scyphomedusae, siphonophores, ctenophores, salps and doliolids, appendicularians and chaetognaths in both the seasons. However, the composition of different species in each group varied from all the stations and seasons. The present study found that the diversity and abundance of the gelatinous zooplankton are highly influenced by the various physicochemical parameters prevailing in the study area. There was positive correlation between the abundance of gelatinous zooplankton and salinity, temperature and pH of the study area, whereas there were a negative correlation between dissolved oxygen and abundance of gelatinous zooplankton.
ACKNOWLEDGMENT
Authors would like to thank the funding agency of the project from the Ministry of Earth Science and the Director, Faculty of Science, Annamalai University, Parangipettai, for the facility given throughout the study period.
REFERENCES
- Ahmed, M.S., M. Raknuzzamman, H. Akther and S. Ahmed, 2007. The role of cyanobacteria blooms in cholera epidemic in Bangladesh. J. Applied Sci., 7: 1785-1789.
CrossRefDirect Link - Ali, M., Abdus Salam, S. Jamshaid and T. Zahra, 2003. Studies on biodiversity in relation to seasonal variations in water of river Indus at ghazi Ghatt, Punjab, Pakistan. Pak. J. Biol. Sci., 6: 1840-1844.
CrossRefDirect Link - Andreu, P., C. Marrase and E. Berdalet, 1989. Distribution of epiplanktonic Chaetognatha along a transect in the Indian Ocean. J. PIankton Res., 11: 185-192.
Direct Link - Ballard, L. and A. Myers, 2000. Observations on the seasonal occurrence and abundance of gelatinous zooplankton in Lough Hyne, co. Cork, South-west Ireland. Biol. Environ.: Proc. Royal Irish Acad., 100B: 75-83.
Direct Link - Bamstedt, U., 1990. Trophodynamics of the scyphomedusae Aurelia aurita, predation rate in relation to abundance, size and type of prey organism. J. Plankton Res., 12: 215-229.
Direct Link - Brodeur, R.D., H. Sugisaki and G.L. Hunt, 2002. Increases in jellyfish biomass in the Bering Sea: Implications for the ecosystem. Mar. Ecol. Prog. Ser., 233: 89-103.
Direct Link - Chae, J., H.W. Choi, W.J. Lee, D. Kim and J.H. Lee, 2008. Distribution of a pelagic tunicate, Salpa fusiformis in warm surface current of the eastern Korean waters and its impingement on cooling water intakes of Uljin nuclear power plant. J. Environ. Biol., 29: 585-590.
PubMed - Clarke, C. and J.C. Roff, 1990. Abundance and biomass of herbivorous zooplankton off Kingston, Jamaica, with estimates of their annual production. Estuarine Coastal Shelf Sci., 31: 423-437.
Direct Link - El-Sherbiny, M.M., M.H. Hanafy and M.A. Aamer, 2007. Monthly variations in abundance and species composition of the epipelagic zooplankton off Sharm El-Sheikh, Northern Red Sea. Res. J. Environ. Sci., 1: 200-210.
CrossRefDirect Link - Guher, H., T. Kirgiz, B. Camur and U. Guner, 2004. A study on zooplankton organisms community structures of Lake Terkos (Istanbul-Turkey). Pak. J. Biol. Sci., 7: 566-570.
CrossRefDirect Link - Haddad, Jr. V., F.L. da Silveira, J.L.C. Cardoso and A.C. Morandini, 2002. A report of 49 cases of cnidarians envenoming from southeastern Brazilian coastal waters. Toxicon, 40: 1445-1450.
Direct Link - Haddock, S.H.D., 2007. Comparative feeding behavior of planktonic ctenophores. Integr. Comp. Biol., 47: 847-853.
Direct Link - Harris, G.R., F.B. Griffiths, L.A. Clementson and H. Van der Doe 1991. Seasonal and interannual variability in physical process, nutrient cycling and the structure of the food chain in Tasmanian shelf waters. J. Plankton Res., 13: 109-131.
Direct Link - Hay, S.J., J.R.G. Hislop and A.M. Shanks, 1990. North Sea scyphomedusae: Summer distribution, estimated biomass and significance particularly for 0-group gadoid fish. Netherland J. Sea Res., 25: 113-130.
Direct Link - Hopcroft R.R., J.C. Roff and H.A. Bouman, 1998. Zooplankton growth rates: The larvaceans Appendicularia, Fritillaria and Oikopleura in tropical waters. J. Plankton Res., 20: 539-555.
Direct Link - Hosia, A., L. Stemmann and M. Youngbluth, 2008. Distribution of net-collected planktonic cnidarians along the northern mid-atlantic ridge and their associations with the main water masses. Deep-Sea Res. Part II: Topical Stud. Oceanography, 55: 106-118.
Direct Link - Kumar, C.S. and P. Perumal, 2011. Hydrobiological investigations in ayyampattinam coast (Southeast coast of India) with special reference to zooplankton. Asian J. Biol. Sci., 4: 25-34.
CrossRefDirect Link - Laval, P., 1997. A virtual mesocosm with artificial salps for exploring the conditions of swarm development in the pelagic tunicate Salpa fusiformis. Mar. Ecol. Prog. Ser., 154: 1-16.
Direct Link - Lindahl, O. and L. Hernroth, 1983. Phyto-zooplankton community in coastal waters of western Sweden-An ecosystem off balance?. Mar. Ecol. Prog. Ser., 10: 119-126.
Direct Link - Mahboob, S. and M. Zahid, 2002. Seasonal limnological variation in Rakh branch canal, Samundri road, Faisalabad. J. Biological Sci., 2: 217-219.
CrossRefDirect Link - Matsakis, S. and R.J. Conover, 1991. Abundance and feeding of medusae and their potential impact as predators on other zooplankton in Bedford Basin (Nova Scotia, Canada) during spring. Can. J. Fish. Aquat. Sci., 48: 1419-1430.
Direct Link - Menard, F., S. Dallot, G. Thomas and J.C. Branconnot, 1994. Temporal fluctuations of two mediterranean salp populations from 1967 to 1990. Analysis of the influence of environmental variables using a Markov chain model. Mar. Ecol. Prog. Ser., 104: 139-152.
Direct Link - Michel, H.B., 1995. The chaetognath Aidanosagitta bedfordii (Doncaster 1903) in the Persian gulf. Bull. Mar. Sci., 56: 490-494.
Direct Link - Mills, C.E., 2001. Jellyfish blooms: Are populations increasing globally in response to changing ocean conditions. Hydrobiologia, 451: 55-68.
Direct Link - Nair, V.R., M. Terazaki and K.V. Jayalakshmy, 2002. Abundance and community structure of chaetognaths in the northern Indian Ocean. Plankton Biol. Ecol., 49: 27-37.
Direct Link - Oueda, A., W. Guenda, A.T. Kabre, F. Zongo and G.B. Kabre, 2007. Diversity, abundance and seasonal dynamic of zooplankton community in a South-Saharan reservoir (Burkina Faso). J. Boil. Sci., 7: 1-9.
CrossRefDirect Link - Pages, F., H.E. Gonzales, M. Ramon, M. Sobarzo and J.M. Gili, 2001. Gelatinous zooplankton assemblages associated with water masses in the humboldt current system and potential predatory impact by Bassia bassensis (Siphonophora: Calycophorae). Mar. Ecol. Prog. Ser., 210: 13-24.
Direct Link - Palma, S. and P. Apablaza, 2004. Seasonal abundance and vertical distribution of the carnivorous gelatinous zooplankton in an upwelling area of the northern humboldt current system. Invest. Mar. Valparaiso, 32: 49-70.
Direct Link - Palma, S. and S. Rosales, 1995. Composition, distribution and seasonal abundance of macroplankton in Valparaiso Bay. Invest. Mar. Valparaiso, 23: 49-66.
Direct Link - Pielou, E.C., 1966. The measurement of diversity in different types of biological collections. J. Theor. Biol., 13: 131-144.
CrossRefDirect Link - Purcell, J.E., 1997. Pelagic cnidarians and ctenophores as predators: Selective predation, feeding rates and effects on prey populations. Ann. Inst. Oceanographique, 73: 125-137.
Direct Link - Purcell, J.E., T.A. Shiganova, M.B. Decker and E.D. Houde, 2001. The ctenophore Mnemiopsis in native and exotic habitats: U.S. estuaries versus the black Sea basin. Hydrobiologia, 451: 145-176.
Direct Link - Rahimibashar, M.R., A. Esmaeili-Sary, S.A. Nezami, A. Javanshir, S.M.R. Fatemi and S. Jamili, 2009. The planktonic community structure and fluxes nutrients in the sefid-rood river estuary (South Caspian Sea). Res. J. Environ. Sci., 3: 149-162.
CrossRefDirect Link - Raskoff, A.K., F.A. Sommer, W.M. Hamner and K.M. Cross, 2003. Collection and culture techniques for gelatinous zooplankton. Biol. Bull., 204: 68-80.
Direct Link - Raskoff, K.A., J.E. Purcell and R.R. Hopcroft, 2005. Gelatinous zooplankton of the Arctic Ocean: In situ observations under the ice. Polar Biol., 28: 207-217.
Direct Link - Santhakumari, V.L., R. Tiwari and V.R. Nair, 1999. Species composition, abundance and distribution of hydromedusae from Dharamtar estuarine system, adjoining Bombay Harbor. Indian J. Mar. Sci., 28: 158-162.
Direct Link - Saravanakumar, A., M. Rajkumar, J. Sesh Serebiah and G.A. Thivakaran, 2007. Abundance and seasonal variations of zooplankton in the arid zone mangroves of gulf of Kachchh-Gujarat, Westcoast of India. Pak. J. Biol. Sci., 10: 3525-3532.
CrossRefPubMedDirect Link - Shannon, C.E. and W. Weaver, 1949. The Mathematical Theory of Communication. 1st Edn., University of Illinois Press, Urbana, IL., ISBN-10: 0252725484, Pages: 117.
Direct Link - Sornes, T.A. and D.L. Aksnes, 2004. Predation efficiency in visual and tactile zooplanktivores. Limnol. Oceanogr., 49: 69-75.
Direct Link - Stemmann, L., A. Hosia, M.J. Youngbluth, H. Soiland, M. Picheral and G. Gorsky, 2008. Vertical distribution (0–1000 m) of macrozooplankton, estimated using the underwater video profiler, in different hydrographic regimes along the northern portion of the mid-atlantic ridge. Deep-Sea Res. Part II: Topical Stud. Oceanogr., 55: 94-105.
Direct Link - Suchman, C.L. and R.D. Brodeur, 2005. Abundance and distribution of large medusae in surface waters of the northern California current. Deep-Sea Res. Part II, 52: 51-72.
Direct Link - Uye, S. and S. Ichino, 1995. Seasonal variations in abundance, size composition, biomass and production rate of Oikopleura dioica (Fol) (Tunicata: Appendicularia) in a temperate eutrophic inlet. J. Exp. Mar. Biol. Ecol., 189: 1-11.
Direct Link - Venkataraman, K., 2005. Coastal marine biodiversity of India. Indian J. Mar. Sci., 34: 57-75.
Direct Link - Zakaria, H.Y., 2004. Pelagic coelenterates in the waters of the western part of the Egyptian Mediterranean coast during summer and winter. Oceanologia, 46: 253-268.
Direct Link