
Farah F. Haron
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Kamaruzaman Sijam
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Dzolkhifli Omar
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Mawardi Rahmani
Department of Chemistry, Faculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 3 | Page No.: 158-162







ABSTRACT
Anthracnose is an economically important disease that can cause 10 to 80% yield reduction in the market with its typical symptoms of dark spots, sunken necrotic tissues and concentric rings of acervuli on fruits. One of the important methods to control the disease is by using antifungal agents derived from plant. Preliminary study indicated that the extracts of Allamanda species have the potential to be developed as the target agent. Five Allamanda species were extracted and screened for antifungal activity against plant pathogenic fungus Colletotrichum gloeosporioides by using poison agar technique. The three chloroform extracts of A. blanchetti, A. cathartica ‘Alba’ and A. cathartica ‘Jamaican Sunset’ exhibited potent inhibitory effects and suppressed the mycelial growth of C. gloeosporioides by up to about 70%. Further study by bioautography-guided fractionation of the extracts led to the isolation and identification of tetracyclic sesquiterpene plumericin as the antifungal agent. The structural determination of the compound was carried out by interpreting the IR, MS, 1D-NMR and 2D-NMR spectral data.
PDF Abstract XML References Citation
Received: July 27, 2012;
Accepted: March 22, 2013;
Published: May 30, 2013
How to cite this article
Farah F. Haron, Kamaruzaman Sijam, Dzolkhifli Omar and Mawardi Rahmani, 2013. Bioassay-guided Isolation of Antifungal Plumericin from Allamanda
Species (Apocynaceae). Journal of Biological Sciences, 13: 158-162.
DOI: 10.3923/jbs.2013.158.162
URL: https://scialert.net/abstract/?doi=jbs.2013.158.162
DOI: 10.3923/jbs.2013.158.162
URL: https://scialert.net/abstract/?doi=jbs.2013.158.162
INTRODUCTION
Genus Allamanda from Apocynaceae family is a popular warm climate plant with trumpet-shaped flowers covering the whole plant throughout the year, easily cultivated, fast growing and gives beauty to any landscape. It is woody evergreen tree with vigorous growth. Allamandas are different in shapes and sizes from small shrubs, big shrubs to vines that can cover the whole fence with beautiful flowers when bloom. The genus is native to South and Central America and had become neutralized throughout the tropical areas (Min et al., 2006). In traditional herbal medicine, leaves and roots extracts of Allamanda cathartica are used as a strong purgative, treating malaria, jaundice and enlarged spleen (Liogier, 1995; Nayak et al., 2006). The extracts were also reported to possess remarkable wound healing property for treating various types of wounds in human besides its antibiotic action against Staphylococcus (Nayak et al., 2006). Allamanda species has been demonstrated as antifungal, antileukemic, anti-HIV (Kardono et al., 1990; Tan et al., 1991), anticancer (Dobhal et al., 2004), cytotoxic activity against Madison lung carcinoma (Abdel-Kader et al., 1997) and strong fungitoxicity against some dermatophytes causing dermatomycosis in animals and humans (Tiwari et al., 2002).
The major postharvest disease of tropical fruits is anthracnose caused by Colletotrichum gloeosporioides and the infections can occur on immature and ripe fruits. To reduce this problem, fungicides were applied either at preharvest or postharvest period (Dodd et al., 1989; Muirhead and Gratitude, 1986). However, the damage it causes is more important in the postharvest period. Due to public concerned about toxic effect of fungicides, their uses had been restricted. Moreover, few farmers in developing countries can afford to purchase these expensive chemicals. Biocontrol agents are economically cheaper and suitable for small scale farmers in developing countries and offer an attractive alternative to overcome this problem. Only a few biocontrol agents have been reported for postharvest protection of tropical fruits against diseases (Govender and Korsten, 2006; Jager et al., 2001; Koomen and Jeffries, 1993). The primary move in finding new biocontrol agents and agrochemicals is bioassay-directed screening of plant extracts and compounds obtained from them. Since, there has been no report on the major active antifungal substances in Allamanda species extracts responsible for controlling C. gloeosporioides, causal agent of papaya anthracnose, the aim of the present study is to evaluate the antifungal activity of Allamanda species and bioassay-guided separation of the active components.
MATERIALS AND METHODS
Instrumentation: This research work was conducted between 2009 and 2012. The IR spectra were recorded on a Perkin Elmer FTIR 1650 spectrophotometer using KBr discs measured in cm-1; MS were recorded on a SHIMADZU GC-MS QP2010 Plus with EI electron impact ion source of 70eV; 1H-NMR 13C-NMR, DEPT-135, HMQC, HMBC, COSY NMR spectra were obtained on a Varian AS 400 spectrometer using CDCl3 as solvent; High Performance Flash Chromatography (HPFC) was performed using a Biotage Inc. Horizon pump (Charlottesville, VA) equipped with a Horizon flash collector and a fixed wavelength (254 nm) detector (system from Biotage Inc. Dyax Corp.). Biotage columns (surface area 500 m2 g-1, porosity 60 Å, particle size 40-63 μm) used were made up of FLASH 12+M, KP-SiL (12x150 mm) and FLASH 25+M, KP-SiL (25x150 mm); TLC was carried out on commercial TLC plates with fluorescent indicator (250 μm, silica gel GF Uniplate, Analtech Inc., Newark DE), developing with solvent mixture made up of EtOAc and hexane, visualization either under UV light at 254 or 365 nm and/or by spraying with anisaldehyde and heating at 110°C.
Statistical analysis: The treatments were arranged in a completely randomized design in four replicates with Allamanda spp., types of extracts and concentration levels as factors. All data were subjected to analysis of variance (ANOVA) where significant (p<0.05) differences between means were determined by Tukey’s standardized range test (HSD). Relationships (quadratic) between colony diameter and extract concentration were determined by regression analysis of data obtained for the different Allamanda species and types of extracts. The SAS (version 9.2) software was used to perform all analyses.
Plant materials: Allamanda blanchetti, Allamanda cathartica, Allamanda cathartica ‘Alba’, Allamanda cathartica ‘Jamaican Sunset’ and Allamanda oenotheraefolia were obtained from nurseries in Sungai Buloh, Selangor and University Agriculture Park, UPM Serdang, Malaysia. The leaves were washed, air-dried at room temperature, ground into powder and stored until required for extraction.
Plant extraction and bioassay: The dried fine powdered leaf of each plant (1 kg) was separately and sequentially extracted with petroleum ether, chloroform and methanol at room temperature. The extracts were filtered through Whatman No. 1 filter paper and concentrated by rotary evaporator to give fifteen dark sticky semisolid extracts. The stock solutions (100 mg mL-1) were serially diluted with acetone to obtain the desired concentration for test solutions of 1, 3, 5 and 7 mg mL-1. Effect of Allamanda extracts on mycelia growth of C. gloeosporioides was measured in vitro according to the method described by Bautista-Banos et al. (2003) using poison agar technique. PDA was amended with 4 concentrations of each extract (1, 3, 5 or 7 mg mL-1). Fifteen mL of the amended PDA was poured into petri dishes (9 cm) and a fungal plug (4 mm) from pure culture of C. gloeosporioides was placed in the centre of the dishes. Petri dishes were then incubated at room temperature and radial measurements of growth were recorded daily until the fungus reached the edge of the plate. Petri dishes containing PDA and acetone served as control. Percentage Inhibition of Radial Growth (PIRG) was calculated after 8 days of incubation using a standard formula:
where, R1 is the radial growth of fungus in the control plate (mm), while R2 is the radial growth of the fungus containing extracts (mm) (Sivakumar et al., 2000).
Bioautography-guided fractionation and isolation of plumericin: The chloroform extract of A. cathartica ‘Jamaican Sunset’ (5 g) was absorbed into silica gel Biotage samplet and applied to a Biotage column FLASH 25+M, KP-SiL (150x25 mm i.d.; 40 mL min-1). Elution of the column was performed by using mixtures of EtOAc:hexane (1:9, 3:7; 1:1; 7:3; 9:1) to give 240 fractions of 50 mL each. Based on TLC analysis, similar fractions were combined and their activity was monitored by bioautography. The active combined fractions 86-112 were further separated by preparative TLC (20x20 cm) and eluted twice with ether:hexane (1:1) and the two active bands were scraped off separately. The scraped bands were dissolved in dichloromethane, filtered through cotton wool to give two white solids. Due to insufficient amount of one of the samples, only the major and active component was submitted for further spectroscopic analysis and identified as plumericin. IR vmax cm-1 (CHCl3): 2921, 2854 (CH), 1759 (C = O), 1696 (C = O), 1448 (C = C); 1H-NMR (CDCl3, 400 MHZ)-see Table 1; 13C-NMR (CDCl3, 400 MHZ)-see Table 1; EIMS m/z (%): 290 (M+) (35), 272 (30), 258 (60), 230 (100), 212 (20), 193 (60), 173 (45), 160 (70), 145 (60), 139 (85), 115 (55), 105 (40).
| Table 1: | 1H-NMR (400 MHZ, CDCl3) and 13C-NMR (100 MHZ, CDCl3) spectral data of plumericin |
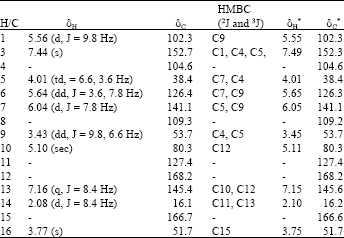 | |
| *Brigitta et al. (2005) | |
Bioautography: TLC plates for bioautography assays were spotted with 80 μg of crude plant extracts and selected fractions and then chromatographed in duplicate (Fig. 1). Fungicide technical grade standards benomyl, cyprodinil, captan (Chem Service, Inc., West Chester, PA) and azoxystrobin (Syngentia, Greensboro, NC) were used as controls at 2 μg in 4 μL of acetone. TLC plates were sprayed with spore suspension adjusted to a final concentration of 1.0x06 conidia mL-1 with liquid potato dextrose broth (PDB, Difco) and 0.1% Tween-80 in order to detect biological activity directly on the TLC plate. Using a 50 mL chromatographic sprayer, each TLC plate with a fluorescent indicator was sprayed lightly with the conidial suspension. Inoculated plates were then placed in a 30x13x7.5 cm moisture chamber (25°C, 100% relative humidity, Pioneer Plastics, Inc., Dixon, KY) and incubated in a growth chamber at 24±1°C. Inhibition of fungal growth was measured 4 days after treatment. The sensitivity of the fungus to each test sample was indicated by the presence of clear zones of fungal growth inhibition directly on TLC plates.
RESULTS AND DISCUSSION
The petroleum ether, chloroform and methanol extracts of the five plants were monitored for antifungal activity by poison agar technique and the three chloroform extracts of A. blanchetti (CEB), A. cathartica ‘Alba’ (CEA) and A. cathartica ‘Jamaican Sunset’ (CEJS) with strong antifungal activity were selected for further investigation. TLC development of these extracts eluted with ethyl acetate:hexane (3:7) revealed the presence of several prominent spots in each plate with the same Rf values range from 0.17-0.77 (Fig. 1a). The plates were then sprayed with C. gloeosporioides and incubated for 4 days at 24±1°C.
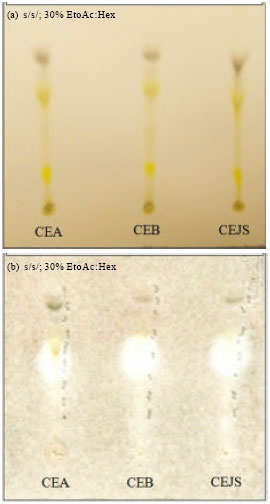 | |
| Fig. 1(a-b): | (a) TLC chromatograms and (b) Bioautograms of A. cathartica ‘Alba’ (CEA), A. blanchetti (CEB) and A. cathartica ‘Jamaican Sunset’ (CEJS) chloroform extracts against C. gloeosporioides in 30% ethyl acetate:hexane as solvent system. (Clear zones indicate inhibition of fungal growth by compounds in the extracts after 4 days of incubation at 24±1°C |
Very clear and prominent zones were visible on three of the active spots with the same Rf values (0.3-0.55) on the bioautograms (Fig. 1b). This clearly indicated the presence of phytochemicals which inhibited the growth of C. gloeosporioides. The chloroform extract of A. cathartica ‘Jamaican Sunset’ (CEJS) was selected for larger scale separation by using High Performance Flash Chromatography (HPFC) with Biotage column FLASH 12+M, KP-SiL (12x150 mm) and eluted with various mixtures of solvents to give 240 fractions of 50 mL each. All fractions collected were monitored by bioautography and the active fractions (86-112) with prominent clear zones were combined and further purified by preparative TLC.
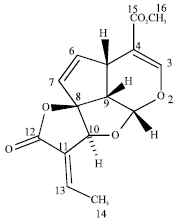 | |
| Fig. 2: | Structure of plumericin |
The major and prominent active spot was scraped, dissolved in dichloromethane, filtered and on solvent removal white solid was obtained. Since the Rf values of the active spots for the two other chloroform extracts of A. blanchetti and A. cathartica ‘Alba’ were identical to the isolated compound, no further separation work was carried on them.
Confirmation of the structure of the isolated compound (Fig. 2) was carried out by comparing the spectral data obtained with literature values. The IR spectrum revealed existence of two carbonyl groups as indicated by the occurrence of prominent absorptions at 1696 and 1759 cm-1. The mass spectrum showed a molecular ion peak at m/z 290 which corresponded to the molecular formula C15H14O6 and the base peak occurred at m/z 230 due the cleavage of -CO2CH3. The integration of 1H-NMR indicated the presence of fourteen protons which further supported the suggested molecular formula (Table 1). The two methyl groups observed at δ 2.08 (day) and 3.77 (sec) are assigned to C-14 and C-16, respectively. The rest of the proton signals are due to the occurrence of eight methine groups and they are assigned based on coupling constants, COSY and HMBC correlations spectra. Two of them are observed as singlets at δ 7.44 (H-3) and 5.10 (H-10). Another two occurred as doublets at δ 5.56 and 6.04 assigned for H-1 and H-7. A quartet observed at δ 7.16 which exhibited 2J correlation with the carbonyl group (C-12) is assigned to H-13. The 2J and 3J correlations between protons and carbons observed in the HMBC spectrum further supported the assignments of other methine groups. The attachment of the carboxylate group to the pyran ring is rationalized by the correlation of H-3 to the carboxyl carbon. The existence of fifteen carbon atoms displayed by the 13C-NMR and DEPT spectra are made up of two methyls, eight methines, three quarternary carbons and two carbonyl groups. The assignments of various protons to their respective carbon atoms could be clearly seen in the HMQC spectrum. Based on these detailed spectral analysis and comparison with literature report, the compound is identified plumericin previously isolated and identified from Plumeria rubra (Brigitta et al., 2005).
Plumericin is an important member of tetracyclic sesequiterpene commonly found in various plant families including from different genera of Apocynaceae such as Plumeria, Allamanda and Himatanthus (Silva et al., 2010; Sharma et al., 2011). The antifungal, anticancerous, antiviral and antibacterial actions of plumericin have been reported previously (Singh et al., 2011; Sticher, 1977). The compound isolated from another species within the Apocynaceae (Himatanthus sucuuba) has been demonstrated to have antiparasitic activity against Leishmanial amazonnensis, responsible for cutaneous leishmaniasis (Castillo et al., 2007). It is believed that the strong activity of the tetracyclic plumericin is due the presence of an á-methylene-γ-lactone moiety which is susceptible to undergo a Michael-type addition with biological nucleophiles (Kupchan et al., 1970). Knobloch et al. (1986) reported that sesquiterpenes affect the bacterial processes that include the inhibition of electron transport, protein translocation, phosphorylation steps and other enzyme-dependent reactions. Most probably similar mechanisms of action were responsible for the antifungal activity of the compound under study. Structural characteristics and chemical properties of terpenes and fatty acids played major roles in signifying antifungal activity for compositions containing these compounds with the presence of oxygenated substituent such hydroxyl and carboxyl groups (Glinski and Branly, 2002).
CONCLUSION
Antifungal-guided isolation of the chloroform extract of A. cathartica ‘Jamaican Sunset’ has led to the identification of plumericin as the compound responsible for the potent antifungal activity exhibited by the plant. The structure of the compound was determined by detailed spectroscopic analysis and comparison with previous reports and has the potential to control anthracnose disease caused by pathogenic fungus Colletotrichum gloeosporioides, an economically important disease in postharvest papaya cultivation. This study adds another important development of this interesting compound which has been the target of various chemical syntheses.
ACKNOWLEDGMENT
We gratefully acknowledge Dr. S.O. Duke, Dr. K.M. Meepagala, Mr. J. Martin and Ms. L. Robertson from USDA-ARS-NPURU for technical assistance. We also would like to acknowledge Universiti Putra Malaysia for providing facilities to carry out this project.
REFERENCES
- Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Elsasser, B., K. Krohn, M.N. Akhtar, U. Florke and S.F. Kouam et al., 2005. Revision of the absolute configuration of plumericin and isoplumericin from Plumeria rubra. Chem. Biodiver., 2: 799-808.
CrossRefDirect Link - Castillo, D., J. Arevalo, F. Herrera, C. Ruiz and R. Rojas et al., 2007. Spirolactone iridoids might be responsible for the antileishmanial activity of a Peruvian traditional remedy made with Himatanthus sucuuba (Apocynaceae). J. Ethnopharmacol., 112: 410-414.
CrossRefDirect Link - Dodd, J.C., P. Jeffries and M.J. Jeger, 1989. Management strategies to control latent infectionin tropical fruits. Asp. Applied Biol., 20: 49-56.
Direct Link - Govender, V. and L. Korsten, 2006. Evaluation of different formulations of Bacillus licheniformis in mango pack house trials. Biological Control, 37: 237-242.
CrossRefDirect Link - Jager, E.S., F.C. Wehner and L. Karsten, 2001. Microbial ecology of the mango phylloplane. Microbial. Ecol., 42: 201-207.
CrossRef - Kardono, L.B., S. Tsauri, K. Padmawinata, J.M. Pezzuto and A.D. Kinghorn, 1990. Cytotoxic constituents of the bark of Plumeria rubra collected in Indonesia. J. Nat. Prod., 53: 1447-1455.
PubMedDirect Link - Koomen, I. and P. Jeffries, 1993. Effects of antagonistic micro organisms on the post harvest development of Colletotrichum gloeosporioides on mango. Plant Pathol., 42: 230-237.
CrossRef - Nayak, S., P. Nalabothu, S. Sandiford, V. Bhogadi and A. Adogwa, 2006. Evaluation of wound healing activity of Allamanda cathartica L. and Laurus nobilis L. extracts on rats. BMC Complement. Altern. Med., Vol. 6.
CrossRefDirect Link - Sharma, U., D. Singh, P. Kumar, M.P. Dobhal and S. Singh, 2011. Antiparasitic activity of plumericin and isoplumericin isolated from Plumeria bicolor against Leishmania donovani. Indian J. Med. Res., 134: 709-716.
CrossRef - Silva, J.R.A., C.M. Rezende, A.C. Pinto and A.C.F. Amaral, 2010. Cytotoxicity and antibacterial studies of iridoids and phenolic compounds isolated from the latex of Himatanthus sucuuba. Afr. J. Biotechnol., 9: 7357-7360.
Direct Link - Singh, D., U. Sharma, P. Kumar, Y.K. Gupta, M.P. Dobhal and S. Singh, 2011. Antifungal activity of plumericin and isoplumericin. Nat. Prod. Commun., 6: 1567-1568.
PubMed - Sivakumar, D., R.S.W. Wijeratnam, R.L.C. Wijesundera, F.M.T. Marikar and M. Abeyesekere, 2000. Antagonistic effect of Trichoderma harzianum on postharvest pathogen of rambutan (Nephelium lappaceum). Phytoparasitica, 28: 240-247.
CrossRef - Tan, G.T., J.M. Pezzuto, A.D. Kinghorn and S.H. Hughes, 1991. Evaluation of natural products as inhibitors of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase. J. Nat. Prod., 54: 143-154.
PubMedDirect Link