
M. Baserisalehi
Department of Microbiology, Kazeroun Branch, Islamic Azad University, Kazeroun, Iran
N. Bahador
Department of Microbiology, Science and Research Branch, Islamic Azad University, Fars, Iran
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 3 | Page No.: 152-157







ABSTRACT
Nowadays, bioactive compounds produced by bacteria consider a source of new remedy to eliminate antibiotic-resistant bacteria. Therefore, the main purpose of this study was isolation and identification of soil origin Gram-negative bacteria and their evaluation for production of bioactive compounds. In general, three bioactive compounds producing bacteria were isolated from hundred and twenty five soil samples. These strains identified as Pseudomonas aeruginosa, Pseudomonas putida and Comamonas testosteroni based on phenotypic and 16 SrRNA Gene sequencing methods. The bioactive compounds produced by isolated strains were partially purified and their activities evaluated at different pHs and temperatures. Then, their antimicrobial spectrums assessed against different pathogenic microorganisms. The results obtained indicated that the best temperature and pH for activity of the bioactive compounds were 40-50°C and 6.5-7.5, respectively. In addition, bioactive compounds produced by P. aeruginosa and P. putida showed an antimicrobial property against Staphylococcus aureus, Bacillus cereus, Corynebacterium sp. Listeria monocytogens, Klebsiella pneumonia, Shigella dysentriae and Proteus mirabilis; however, E. coli was resistant to them. Although all the bacteria were resistant to the bioactive compound produced by C. testosteroni, Aspergillus niger and Alternaria sp. were sensitive. Overall bioactive compounds produced by Pseudomonas sp. showed activity against some pathogenic bacteria as well as fungi and might be considered an alternative therapeutics not only in pharmaceutical but also as a preservative in food industries.
PDF Abstract XML References Citation
Received: January 14, 2013;
Accepted: March 15, 2013;
Published: May 30, 2013
How to cite this article
M. Baserisalehi and N. Bahador, 2013. Evaluation of Soil Origin Pseudomonas sp. for Production of Bioactive Compounds. Journal of Biological Sciences, 13: 152-157.
DOI: 10.3923/jbs.2013.152.157
URL: https://scialert.net/abstract/?doi=jbs.2013.152.157
DOI: 10.3923/jbs.2013.152.157
URL: https://scialert.net/abstract/?doi=jbs.2013.152.157
INTRODUCTION
Bioactive compounds are peptides produced by various bacteria with antimicrobial property against different microorganisms (Motta and Brandelli, 2008). These compounds with use of hydrophobic/amphiphilic properties penetrate into the cell and consequently the bacterial compositions release from the cell (Oscariz and Pisabarro, 2001). However, production of bioactive compounds such as bacteriocins is similar to antibiotics but several advantages could be considered in compared to the antibiotics. For example, toxicity of bacteriocins on animal tissues is relatively nil. In addition, proteinaceous property of these compounds resulted in their degradation by protease enzymes; hence could not accumulate in the human body (Hechard and Sahl, 2002). In general, Bacteriocins are divided into four groups based on their structures. Class I bacteriocins are small peptides (<5 kDa) with unusual amino acids. Class II are thermo stable peptides with size of <10 kDa. Class III are heat-sensitive with high molecular weight (<30 kDa) and Class IV are relatively complex with glycoprotein, lipoprotein and non-protein moiety (Ennahar et al., 2000). It is confirmed that non-protein moieties of bacteriocins require for stability and antimicrobial activity (Garneau et al., 2002).
Nowadays, application of bacteriocins considers as a way to reduce the frequency of occurrence of antibiotic-resistant bacteria. For examples, alternative drugs as well as safe food preservatives could eliminate antibiotic-resistant strains of bacteria and increase shelf life of foods (Settanni and Corsetti, 2008). Therefore, based on foregoing evidence present study was conducted to isolate and identify bacteriocin-producing bacteria from soil and assessed their antimicrobial effect against some pathogenic bacteria.
MATERIALS AND METHODS
Isolation of gram negative bacteria from soil samples: One hundred and twenty five soil samples were collected from different areas of kazeroun city, Iran. Isolation of potent bioactive producing bacteria was carried out by serial dilution of soil sample (10-1, 10-2, 10-3 and 10-6) using sterile distilled water. Then, 0.1 mL of each dilution was transferred onto MacConkey agar and the plates were incubated at 35°C for 24 h. After this period the growth colony was picked up and subjected for assay the bioactive compound production.
Screening of bioactive compound producing bacteria: The bacterial colonies obtained from soil samples separately were inoculated into hundred milliliters of trypticase soy broth in the conical flask (250 mL) and incubated at 35°C, 200 rpm for 24 h. Then, 5 mL of each culture broth was centrifuged at 12000 rpm for 30 min and the supernatant was assessed for antimicrobial property against bioassay strains viz., E. coli PTCC 1330, Staphylococcus aureus, PTCC 1337 Bacillus cereus PTCC 1137, Pseudomonas aeruginosa PTCC1556 and Candida albicans PTCC 5027. Screening of bioactive compound producing strains in this study was done by the Well Diffusion Agar (WDA) method. To perform the test, bioassay strain was cultivated on Mueller Hinton agar and wells (5 mm in diameter) were made in plate agar using sterile sharp borer. Then, 100 μL of each supernatant was added into each well and the plates were incubated at 35°C. After 24 h, observation of a clear zone of growth inhibition considered bioactive compound activity (Ten Brink et al., 1994).
Phenotypic identification of bioactive compound producing strains: Promising bacteria for production of bioactive compounds were presumptive identified by Gram stain, growing on MacConkey agar, catalase and oxidase tests and then identification was confirmed using O/F, Nitrate, TSI, Urease, Lysin decarboxylase and Phenylalanine tests.
Authentication of antimicrobial producing strains: Verification of presumptive identification was carried out by Gene sequencing of 16SrRNA.
To perform the test DNA was extracted from the isolated strains using High Pure Template DNA PCR kit (Roche-Germany) according to the supplier's instructions. The purity of the extracted DNA was assessed based on absorbance of the extracted DNA at 260 and 280 nm wavelengths by Biophotometer (Eppendrof-Germany) and then the purity was calculated based on absorbance ratio 260/280 nm. The extracted DNA with ratio (260/280 nm) 1.9≤corresponding to 121 μg DNA mL-1 was used for amplification of 16S rRNA by PCR. Amplification of 16S rRNA was performed using universal primers zroduced by TAG K openhagen (Denmark). The sequence of Forward and Reverse primers were 5'-AGGAGGTGATCCAACCGCA-3' and 5'-AACTGGAGGAAGGTGGGGA-3', respectively. Each reaction was carried out in a total volume of 25.5 μL contained 14.5 μL of molecular biology-grade water (Sigma Aldrich Company Ltd.), 2.5 μL of 10xPCR buffer (Cinagen-Iran), 1 μL of each forward and reverse PCR primers, 1 μL of a 10 mM dNTPs (Cinagen-Iran), 0.5 μL of Smar taq polymerase (cinagen-Iran), 1 μL of 50 mM MgCl2 (cinagen-Iran) and 5 μL of DNA template.
PCR amplification conditions on an Eppendrof thermocycler were as follows: 95°C for 4 min, followed by 35 cycles of 95°C for 40 sec, 56°C for 30 sec and 72°C for 40 sec, with a final extension at 72°C for 5 min and storage at 4°C. All PCR products obtained were run on a 1.5% (w/v) agarose gel with a 100 bp DNA ladder (Fermntas-Russia). PCR products were electrophoresed at 75V for 20 min; DNA was visualized using ethidium bromide and photographed by Uvidoc (England). After visualization of pure DNA bands, the PCR products have been sent to Macrogen in South Korea (http://www.macrogen.com/) for DNA sequencing. The 16S rRNA sequenced data for bioinformatic applications were subjected to BLAST analysis (http://www.ncbi.nlm.nih.gov/BLAST/) in order to identify each respective 16S rRNA gene amplicon.
Assay of bioactive compound production during bacterial growth phases: To assess antimicrobial property of the metabolite during the bacterial growth phases, 5 mL of each bacterial culture was withdrawn at time intervals (5, 10, 15, 20….45 h) and the cells counted by optical density at 600 nm. Then the bacterial culture centrifuged at 12000 rpm for 30 min. Afterward, the antimicrobial property of the supernatant was assessed against bioassay strains (Van Reenen et al., 1998).
Partial purification of the bioactive compounds: The bacterial cultures containing bioactive compound (as described above) were centrifuged at 12000 rpm, 30 min, 4°C. Then, the pellets were removed and ammonium sulfate was added into the supernatants (up to 40%) with stirring. After 12 h mixtures were centrifuged at 12000 rpm, 30 min, 4°C and surface pellicle from each suspension was recovered. The pellicle containing bacteriocin was resuspended in 50 mM sodium phosphate buffer (pH 6.5). Then 25 mL of mixture was treated with chloroform-methanol (2:1, v/v) for 1 h and centrifuged at 12000 rpm, 30 min, 4°C. The precipitate was air dried and dissolved in 10 mL of 10% 2-propanol/0.1% tri fluroaceticacid (TFA).
Effect of different pHs on activity of bioactive compounds: One milliliter of each partial purified bioactive compound was adjusted to pH 5.0, 5.5, 6.0, 6.5, 7.0, 7.5 and 8.0 by diluted NaOH and HCl for 1 h and readjusted to pH 7.0. To assess bacteriocin activity, 100 μL of each compound was added into the well on Mueller Hinton agar seeded by bioassay strain (Sharma et al., 2009). After 24 h, observation of a zone of inhibition was considered bioactive compound activity.
Effect of different temperatures on activity of bioactive compounds: Stability of partial purified bioactive compounds at different temperatures was tested by overlaid of 10 mL of aliquot compounds using paraffin oil. The tubes were incubated at 40, 50, 60 and 70°C for 30 min then, 100 μL of each bioactive compound was added into the well of Mueller Hinton agar that previously seeded by bioassay strain (Sharma et al., 2009). After 24 h, observation of a zone of inhibition was considered bioactive compound activity.
Antimicrobial spectrum of the bioactive compounds: Antimicrobial activity of the compounds against various microorganisms viz., Listeria monocytogens PTCC 1163, Klebsiella pneumonia PTCC 1053, Salmonella typhi PTCC 1609, Shigella dysentriae PTCC 1183 and Proteus mirabilis PTCC 1076, Aspergillus niger PTCC 5011 and Alternaria PTCC 5224 was assessed by Well Diffusion agar method. To perform the test, each bioassay strain was full cultivated on Mueller Hinton agar and three wells were made in plate agar using sterile sharp borer. Then, 100 μL of purified bacteriocin of BC10, Bd9 and BSI were separately added into each well and the plate was incubated at 35°C. After 24 h, clear zone of growth inhibition was measured and considered antimicrobial activity. All experiments were carried out in three replicates.
RESULTS
Isolation of bioactive compound producing strains from soil samples: The results obtained from screening of bioactive compound producing bacteria from soil samples indicated that three strains of Gram-negative bacteria could produce bioactive compounds. Two bioactive compounds (BC10 and Bd9) showed an antimicrobial property against Bacillus cereus and staphylococcus aureus with zone of inhibition of 22 and 20 mm; however, Escherichia coli was resistant to them. Pseudomonas aeruginosa was sensitive (with zone of inhibition of 15 mm) and resistant to BC10 and Bd9, respectively. Furthermore, of all antagonistic microorganisms only Candida albicans was sensitive to bioactive compound BS1.
Phenotypic and molecular identification of bacteriocin producing strains: The results obtained from phenotypic identification of the isolates illustrated that bacteriocin producing bacteria were Pseudomonas sp. However, alignment analysis of 16SrRNA genes of bacteriocins BC10, Bd9 and BSI producing strains exhibited 100, 99 and 99% identical to Pseudomonas aeruginosa strain GDC10 with accession number gb|JN594664.1|, Pseudomonas putida strain CDd9 with accession number gb|GU248219.1| and Comamonas testosteroni with accession number gb|HQ200412.1, respectively (Fig. 1).
Determination of the growth phase of bioactive compounds production: The results obtained from production of bioactive compounds during the bacterial growth phases indicated that all bacteriocins were produced after 10 h and reach to maximum level at 20 h of bacterial growth. It means all the bioactive compounds were primary metabolites. Therefore, they must be noted as bacteriocins.
Effect of various pHs and temperatures on activity of bioactive compounds: As shown in Table 1 activity of three bioactive compounds maintains at pHs 6-7.5 however, at pHs 5, 5.5 and 8 was nil. In addition, BC10 and Bd9 showed weak and strong activity at pHs 6 and 6.5-7.5, respectively. The results obtained from activity of bioactive compounds at different temperatures indicated that the best temperatures for activity of the bioactive compounds were 40 and 50°C. In addition, BC10 and BS1 showed stable and unstable bioactive compounds against different pHs and temperatures, respectively.
Antimicrobial spectrum of the bioactive compounds: The result obtained from the antimicrobial spectrum of the bioactive compounds against some pathogenic microorganisms illustrated that the bioactive compounds BC10 and Bd9 exhibited an antimicrobial property to all bacteria tested with varied responses; however, their antimicrobial effects against fungi were nil.
| Table 1: | Effect of different pHs and temperatures on bioactive compounds |
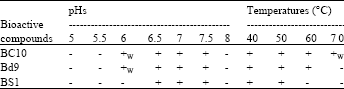 | |
| -: Negative in bacteriocin activity, +: Positive in bacteriocin activity and +W: Weak in bacteriocin activity | |
 | |
 | |
| Fig. 1: | Alignment analysis of 16SrRNA genes of bacteriocin producing strains |
| Table 2: | Antimicrobial spectrum of bioactive compounds against pathogenic microorganisms |
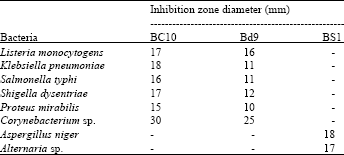 | |
| Numbers represent average of three replicate (mm), R: Resistance | |
In addition, Corynebacterium sp. and Proteus mirabilis exhibited high and less sensitive character. Bioactive compound BS1 showed antimicrobial activity against fungi (Aspergillus niger and Alternaria sp.) (Table 2).
DISCUSSION
Several reports verified the antimicrobial effect of bioactive compounds against pathogenic microorganisms. For instance, bacteriocin as bioactive compound of Paenibacillus polymyxa strongly inhibited growth of Xanthomonas campestris strains CP-1-1 and M-5 and bacteriocin of Bacillus subtilis MTCC-8114 inhibited growth of Microsporum fulvum and Trichophyton species (Hammami et al., 2009; Kumar et al., 2009). Gupta and Garg (2009) reported that protection of urogenital treat of human against pathogenic bacteria is possible by using two strains of bioactive producing bacteria viz., Lactobacillus GG and Lactobacillus rhamnosus GR-1 and therefore, antimicrobial substances produced by these organisms can be considered new antimcrobial agents. Recently application of bacteriocins has focused on their antimicrobial effect because of specific target of these compounds and less frequency of developing bacterial resistance (Ketla et al., 2003). In the present study bacteriocin producing bacteria were detected and identified from soil samples. These isolates were Pseudomonas aeruginosa and Pseudomonas putida and Comamonas testosteroni. Pseudomonas aeruginosa produced bacteriocin BC10, whereas Pseudomonas putida and Comamonas testosteroni produced bacteriocins Bd9 and BS1 respectively. The best pH for activity of bacterocins was 6-7.5 however, the optimum temperature for their activity was 40-50°C. Although, bacteriocins BC10 and Bd9 showed antimicrobial effect against pathogenic bacteria, BS1 exhibited an antimicrobial effect against pathogenic fungi. Concerning to this result it must be noted that Comamonas testosteroni known as Pseudomonas testosteroni before its reclassification by molecular genetic techniques, metabolize testosterone (Gilligan et al., 2003). Although, the pathogenic potential of C. testosteroni has not been well studied, it is usually considered a commensal. C. testosteroni has been implicated in bacteremia-related septic shock, endocarditis and meningitis (Abraham and Simon, 2007). Our results showed antifungal activity of Comamonas testosteroni isolates. Parallel to our report El-Banna (2007) reported an antifungal activity of this bacterium. In addition, several reports confirmed capability of Comamonas testosteroni in degradation of steroids and non-steroid compounds (Arda et al., 2003). In addition several reports affirmed capability of Comamonas testosteroni in degradation of steroids and non-steroid compounds.
CONCLUSION
In general, natural habitat of bacteriocin producing bacteria is soil. In addition, bacteriocins produced by Gram negative bacteria have antimicrobial property against pathogenic microorganisms. Hence, in order to eliminate frequency of antibiotic-resistant bacteria and increase antimicrobial spectrum of the new drugs, production of bacteriocins from Gram-negative bacteria must be considered an especial area of investigation.
REFERENCES
- Abraham, J.M. and G.L. Simon, 2007. Comamonas testosteroni bacteremia: A case report and review of the literature. Infect. Dis. Clin. Pract., 15: 272-273.
CrossRefDirect Link - Arda, B., S. Aydemir, T. Yamazhan, A. Hassan, A. Tunger and D. Serter, 2003. Comamonas testosteroni meningitis in a patient with recurrent cholesteatoma. Acta Pathol. Microbiol. Immunol. Scand, 11: 474-476.
CrossRefDirect Link - Brink, B.T., M. Minekus, J.M.B.M. van der Vossen, R.J. Leer and J.H.J.H. Intveld, 1994. Antimicrobial activity of lactobacilli: Preliminary characterization and optimization of production of acidocin B, a novel bacteriocin produced by Lactobacillus acidophilus M46. J. Applied Bacteriol., 77: 140-148.
CrossRefPubMedDirect Link - Ennahar, S., T. Sashihara, K. Sonomoto and A. Ishizaki, 2000. Class IIa bacteriocins: Biosynthesis, structure and activity. FEMS Microbiol. Rev., 24: 85-106.
CrossRefDirect Link - Garneau, S., N.I. Martin and J.C. Vederas, 2002. Two-peptide bacteriocins produced by lactic acid bacteria. Biochimie, 84: 577-592.
CrossRefDirect Link - Gilligan, P.H., G. Lum, P.A.R. Vandamme and S. Whittier, 2003. Burkholderia, Stenotrophomonas, Ralstonia, Brevundimonas, Comamonas, Delftia, Pandoraea and Acidovorax. In: Manual of Clinical Microbiology, Murray, P.R., E.J. Baron, J.H. Jorgensen, M.A. Pfaller and R.H. Yolken (Eds.). 8th Edn., ASM Press, Washington, DC.
- Gupta, V. and R. Garg, 2009. Probiotics. Indian. J. Med. Microbiol., 27: 202-209.
CrossRefDirect Link - Hammami, I., A. Rhouma, B. Jaouadi, A. Rebai and X. Nesme, 2009. Optimization and biochemical characterization of a bacteriocin from a newly isolated Bacillus subtilis strain 14B for bio-control of Agrobacterium sp. strains. Lett. Applied Microbiol., 48: 253-260.
CrossRefDirect Link - Hechard, Y. and H.G. Sahl, 2002. Mode of action of modified and unmodified bacteriocins from gram-positive bacteria. Biochimiestry, 84: 545-557.
CrossRefDirect Link - Ketla, T., K. Naterstad, M. Vancanneyt, J. Swings and L. Axelsson, 2003. Differences in susceptibility of Listeria monocytogenes strains to Sakacin P, Sakacin A, Pediocin PA-1 and Nisin. Applied Environ. Microbiol., 69: 4431-4437.
CrossRefDirect Link - Kumar, A., P. Saini and J.N. Shrivastava, 2009. Production of peptide antifungal antibiotic and biocontrol activity of Bacillus subtilis. Indian J. Exp. Biol., 47: 57-62.
Direct Link - Motta, A.S. and A. Brandelli, 2008. Evaluation of environmental conditions for production of bacteriocin-like substance by Bacillus sp. strain P34. World J. Microbiol. Biotechnol., 24: 641-646.
CrossRefDirect Link - El-Banna, N.M., 2007. Antifungal activity of Comamonas acidovorans isolated from water pond in south Jordan. Afr. J. Biotechnol., 6: 2216-2219.
Direct Link - Oscariz, J.C. and A.G. Pisabarro, 2001. Classification and mode of action of membrane-active bacteriocins produced by gram-positive bacteria. Int. Microbiol., 4: 13-19.
PubMed - Settanni, L. and A. Corsetti, 2008. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol., 121: 123-138.
CrossRefDirect Link - Sharma, N., G. Kapoor, N. Gautam and B. Neopaney, 2009. Charactrization of a partially purified bacteriocin of Bacillus sp. MTCC 43 isolated from Rhizosphere of radish (Raphanus sativus) & its application as a potential food biopreservative. J. Scient. Indust. Res., 68: 881-886.
Direct Link - Van Reenen, C.A., L.M. Dicks and M.L. Chikindas, 1998. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum. J. Applied Microbiol., 84: 1131-1137.
CrossRefPubMedDirect Link