
Prayook Srivilai
Department of Biology, Faculty of Science, Mahasarakham University, Khamriang Sub-District, Kuntarawichai District, Mahasarakham Province, 44150, Thailand
Wasana Chaiseana
Faculty of Science and Agricultural Technology, Rajamangala University of Technology Lanna, Phisanuklok Province, Thailand
Panida Loutchanwoot
Department of Biology, Faculty of Science, Mahasarakham University, Khamriang Sub-District, Kuntarawichai District, Mahasarakham Province, 44150, Thailand
Piyarat Dornbundit
Department of Chemistry, Faculty of Science, Srinakarintarawirot University, Sukhumvit 23, Bangkok, Thailand
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 3 | Page No.: 131-138







ABSTRACT
Cambodian Phellinus linteus is a white rot fungus that behaves as a plant pathogen. This fungus was first used for studying changes in the wood chemistry and structure of Shorea (Shorea obtusa). Cambodian P. linteus comprises two different types of mycelia, namely monokaryon (n) and dikaryon (2n). Wood blocks of Shorea were exposed to monokaryons and dikaryons under in vitro condition and biodegradation took place over a 12-week-period. Shorea wood block degradation was determined by (1) wood weight loss (2) Fourier Transform Infrared (FTIR) and (3) Scanning Electron Microscopy (SEM). Results showed that the degradation of Shorea wood blocks by monokaryons and dikaryons was 57.48 and 55.73%, respectively. The FTIR analyses showed that the Shorea wood blocks degraded by the monokaryons and dikaryons were differed in their chemical components (aromatic, C-H and C-O). The results also suggested that the lignin and carbohydrates were decayed. Meanwhile, the C-O, C-O-H and C-H groups were decreased, revealing that hemicelluloses and cellulose polymers were broken down by hydrolytic enzymes during fungal growth. SEM was used to observe the physical changes of the Shorea wood blocks and it showed that the wood cells were changed. In conclusion, Cambodian P. linteus monokaryons are more appropriated for the paper industry (biopulping and bio-breaching) and enzyme technology than dikaryons, due to monokaryons leave no dark pigments and residues from the fruiting bodies. Moreover, monokaryons have a simple genetic system (n) for genetic and biochemical analyses.
PDF Abstract XML References Citation
Received: December 20, 2012;
Accepted: February 26, 2013;
Published: May 30, 2013
How to cite this article
Prayook Srivilai, Wasana Chaiseana, Panida Loutchanwoot and Piyarat Dornbundit, 2013. Comparison of Differences Between the Wood Degradation by Monokaryons (n)
and Dikaryons (2n) of White Rot Fungus (Cambodian Phellinus linteus). Journal of Biological Sciences, 13: 131-138.
DOI: 10.3923/jbs.2013.131.138
URL: https://scialert.net/abstract/?doi=jbs.2013.131.138
DOI: 10.3923/jbs.2013.131.138
URL: https://scialert.net/abstract/?doi=jbs.2013.131.138
INTRODUCTION
In Southeast Asia, houses are mostly constructed from wood and timbers. Various types of farm equipments are also produced with wood. Wood buildings and farm equipments, including archaeological works, are mostly destroyed by fungi as Southeast Asia is located in a tropical zone and the weather is warm with high humidity, which are appropriate conditions for fungal growth. The fungal spores generate mycelium which is the initial stage of the fungal life cycle. During fungal growth mycelium releases extracellular enzymes that destroy the wood structures, causing a loss of economic value. Various items in the literatures mentioned that wood degradation is caused by white-rot, brown-rot and soft-rot fungi (Blanchette et al., 1985; Pandey and Pitman, 2003, 2004). White, brown and soft-rot fungi are major groups of wood degradation fungi. Fungi slowly destroy wood and trees, unlike wood degradation by insects. Fungal and insect degradations mainly causes death and decreased the number of living trees in forests, i.e., natural forests, public parks and commercial angiosperm forests (Breen and Singleton, 1999). Decay causing fungal species, such as Phanerochaete chrysosporium, Trametes versicolor, Ganoderma spp. Ceriporiopsis subvermispora, Lentinus endodes, Schizophylum commune, Phellinus flavomarginatus and Phellinus igniarius are well known as causes of biodegradation due to the production of hydrolytic enzymes during growth (Blanchette et al., 1985; Ferraz et al., 2000; Pandey and Pitman, 2003, 2004). Most basidiomycetes (mushrooms) consist of two different types of mycelia in a life cycle. There is a sterile primary mycelium or monokaryon/vegetative mycelium (n), containing a nucleus in a hyphal cell and a set of genes and another fertile secondary mycelium or dikaryon (2n), containing two different nuclei per a hyphal cell. The two sets of genes result in the dikaryons being able to form the fruiting body (Kües, 2000; Kothe, 2001). In general, white rotting fungi degrade lignin by secreting extracellular oxidative enzymes that diffuse and immediately attack the wood. Lignin is present in high concentrations in wood cells and their wall components. When lignin is degraded by white rot fungi, defibrillation and dissolution of the middle lamella were occurred resulting in white rot of the wood. In contrast, brown rot fungi specifically decay carbohydrates with limited lignin degradation, causing brown rot of the wood (Blanchette et al., 1985). Phellinus flavomarginatus is a white rot fungus commonly studied in the decomposition of Eucalyptus grandis and found that litter wood biomass was lost (Fischer and Binder, 2004). The effects of white rot fungi on wood components are usually studied by Fourier Transform Infrared Spectroscopy (FTIR), near infrared spectroscopy and Scanning Electron Microscopy (SEM) (Pandey 1999; Pandey and Pitman, 2003, 2004; Worrall et al., 1997; Akio et al., 1988; Fackler et al., 2007). FTIR is a useful technique for investigating wood decay even though sample preparation is limited (Faix 1992; Pandey 1999; Pasikatan et al., 2001; Pandey and Pitman, 2003; Schwanninger et al., 2004; Rana et al., 2010). FTIR has also been used for component analysis of wood including changes in the chemistry of the wood, such as in lignin, cellulose, hemicelluloses and other wood chemistry (Schultz and Glasser, 1986; Rodrigues et al., 1998). Light and electron microscopy have been primarily used to analyse fungal attacks (Raberg and Daniel, 2009). The behaviour of hyphal penetration was studied by SEM (Worrall et al., 1997; Fischer and Binder, 2004; Genestar and Palou, 2006).
Cambodian P. linteus is a plant pathogen that is commonly found in the tropical forests of Cambodia. The fruiting body of this fungus whose weight is approximately 5.5 kg (Natural Medicinal Mushrooms Museum of Mahasarakham University) is bigger than those of other P. linteus specimens. Cambodian P. linteus has two different types of mycelium, i.e., monokaryon and dikaryon. Its mating type system is heterothallic and functions as bipolar. Shorea obtusa is a hardwood tree and a famous commercial wood that has been used for houses and other forms of construction. This tree species is in the Dipterocarpaceae family. It is commonly found in Myanmar, Cambodia, Laos, Vietnam and Thailand. Moreover, S. obtusa is a suitable host tree for P. linteus, which can reach 10-30 m tall (http://en.wikipedia.org/wiki/ Shorea-obtusa).
Most wood degradation studies either used monokaryons or dikaryons, while the comparative data of wood degradation between monokaryons and dikaryons is on the litter. Therefore, the objective of this study was to determine the differences between the wood degradation by monokaryons (n) and dikaryons (2n) of Cambodian P. linteus, focusing on the changes in the structure and chemical composition of Shorea wood. Biodegradation time was 12 weeks and monitoring was accomplished by wood weight loss and FTIR, whereas the behaviour of the hyphal growth in the wood samples was investigated by SEM.
MATERIAL AND METHODS
Wood preparation, fungal strain, inoculation and test conditions: The experiment was performed on October 2011 to January 2012. S. obtusa wood blocks (approximately 1x1x2 cm) were washed with sterile water and oven dried at 60°C, until a constant weight was achieved. The Shorea wood block samples were packed in autoclaved plastic bags and sterilized by autoclaving at 121°C for 15 min and then the moisture was removed by oven drying at 60°C. Then 3 sterilized Shorea wood blocks were placed on stainless steel net supporters closed to the edge of a 250 mL Erlenmeyer flask, containing 100 mL of Malt Extracted Agar (MEA; 10% w/v of malt extract; 12% w/v agar). The wood blocks were then inoculated with a 1.5x1.5 cm agar plug of active monokaryonotic, or dikaryotic mycelium and loosely closed by a sterilized cotton plug and kept at room temperature (30-32°C, 50-60% humidity) for 90 days. The experiment was designed as three replicates (2 monokaryonsx3 = 6 Erlenmeyer flasks and 18 wood blocks; 2 dikaryonsx3 = 6 Erlenmeyer flasks and 18 wood blocks).
Wood weight loss determination: Residual mycelium from the surface of Shorea wood blocks was removed and the wood block samples were oven-dried to a constant weight at 60°C and then weighed. Then the wood weight loss was calculated.
Fourier transforms infrared (FTIR) examination: FTIR spectra of un-decayed and decayed wood block samples were measured by direct transmittance using the KBr pellet technique. Spectra were recorded using a Spectrum GX (FT-IR system 49931) by Perkin Elmer. Small pieces of wood block samples were ground using a mortar. The wood powder was mixed with KBr powder at a concentration of 0.5-1%. Pelleted samples were detected by IR spectroscopy. All spectra were measured at a spectral resolution of 400-4000 cm-1.
Scanning electron microscopy observation: Pieces of Shorea wood block samples were observed by SEM. For SEM performance, Shorea wood samples were fixed in 2.5% glutaradehyde in 0.1 M phosphate buffer (pH 7.2) overnight at 4°C and then washed with the same buffer. The wood samples were then fixed in 1.7 Osmiumtetraoxide for 2 h before being washed with distilled water. Shorea wood samples were fixed and dehydrated in an acetone series (20, 40, 60, 80 and 100%) and then left to air dry. Gold coated sample films were determined using a SEM (JSM 6460 LV).
Statistical analysis: Data is presented as Mean±Standard Deviation (SD). Significant differences among all treatment groups were analysed by one-way ANOVA with the Tukey post-hoc test followed by the Student’s t-test (PrismTM; GraphPad, San Diego, CA, USA). The p values<0.05 were considered significant.
RESULTS AND DISCUSSION
Monokarotic and dikaryotic growth: Active monokaryotic and dikaryotic mycelium were inoculated in 250 mL Erlenmeyer flasks containing 100 mL MEA. Three wood blocks of S. obtusa were placed on the medium surface. Three days after mycelial inoculation, monokaryotic mycelium directly grew on the wood blocks and fully covered the medium surface within 6 days. The growth rate of monokaryotic mycelium was 0.7±0.15 cm per day. At 21 days, monokaryotic mycelium was tightly packed without fruiting body formation. At 90 days, mycelium was white, dense and flat with no fruiting body. Dikaryotic mycelium was exposed to the wood blocks that were placed on the surface of the MEA. Dikaryotic mycelium grew fast, more protuberant and fully covered the medium surface within 5 days. The growth rate of the dikaryontic mycelium was 0.9±0.1 cm day. At 21 days, dikaryotic mycelia developed and formed the initial fruiting bodies (size = 2-5 mm) on both inoculums and wood blocks. The morphological features of the initial fruiting bodies resembled the sponge. At 90 days, mycelium was light brown and the size of the initial fruiting bodies was a bit bigger. The results suggested that dikaryotic mycelium grew more actively than monokaryotic mycelium, which is a comparable growth rate to the dikaryotic mycelium of Coprinopsis cinerea and S. commune and also similar to the mycelial growth rate of the famous white rotting fungus (T. versicolor) (Addleman and Archibald, 1993). Dikaryotic mycelium tended to grow faster with denser and more vigorous propagating characters (Kües, 2000; Kothe, 2001).
Weight loss data: The average initial weight of the wood block samples, i.e., before degradation by monokaryotic mycelium, was 2.083±0.282 g. After 90 days incubation, the average final weight of the wood block samples was 0.886±0.085 g, contributing to the net weight loss of 57.48%. Meanwhile, the average weight of the wood block samples before incubation with dikaryotic mycelium was 2.1140±0.3630 g. After 90 days incubation, the average weight of wood block samples was 0.9358±0.0461 g; the wood weight loss was 55.73% (Fig. 1).
Wood weight loss when decayed by monokaryons was more than those of dikaryons by approximately 1.75%. However, the efficiencies of the wood degradation by monokaryons and dikaryons were not statistically significant difference (Fig. 1). It was indicated that Cambodian P. linteus is highly efficient for wood degradation (approximately 56%) as the other wood decaying fungi such as T. versiolor and P. chrysosporium. Faix et al. (1993) studied the chemical changes in beech wood decayed by the white rot fungi T. versicolor, Pleurotus ostreatus and L. edodes and found that after 14 weeks of incubation, the average wood weight loss were 51, 27 and 24%, respectively (Faix et al., 1993).
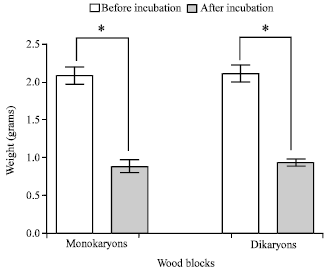 | |
| Fig. 1: | Comparison of the weight of Shorea wood blocks after incubation with monokaryons and dikaryons. *P<0.05 means a statistically significant difference was found in the monokaryotic and dikaryotic degradation |
It was also found that the mean weight losses of Beech and Scots pine decayed by Coriolus (T. versicolor) were 45.5 and 38.8%, respectively, after 12 weeks incubation (Pandey and Pitman, 2003). Weight loss of a soft wood (Pinus radiate) decayed by T. versicolor was 22% and the biodegradation time was 77 days (Ferraz et al., 2000). The results indicated that dikaryons have greater cellular development than monokaryons due to dikaryons require more nutrients than monokaryons and therefore, dikaryons possibly secreted high amounts of extracellular enzymes during their fungal growth to produce much more glucose molecules from the wood components that are need for cell growth. From the principle of genetic systems, dikaryons contain two copies of enzyme genes (homologous), when these genes are expressed the amount of enzymes produced must be twice as much, more than the amount of enzymes produced by monokaryons. Hence, dikaryons would have greater efficiency for wood degradation than monokaryons. In this study, the results found that monokaryons were more efficient for wood degradation than dikaryons, but the difference was not statistically significant. Addleman and Archibald (1993) studied the delignification by monokaryons and dikaryons of T. versicolor and found that delignification by monokaryons was similar to dikaryons. Monokaryons and dikaryons of basidiomycetes, i.e., C. cinerea and P. igniarius, mostly produced equally amounts of laccase enzymes (Srivilai, 2006), these results showed that mycelial growth rate, pigments of mycelium, fruiting body and genetic system would be important information for consideration of mycelium types for various fungal applications.
Undecayed wood and wood decayed by monokaryons and dikaryons: FTIR was used to characterize the composition of the wood, both undecayed and decayed by monokaryons and dikaryons, as shown in Fig. 2a-c. Several studies have done on band assignment for wood samples. The broader signal at ~3500 cm-1 indicated the presence of water vapor in the wood sample powder, a band was observed at 3420 cm-1 (O-H stretching absorption), a prominent C-H stretching vibration was around 2880 cm-1. The region around 1800-600 cm-1 is known as the fingerprint region for wood. Moreover, well defined peaks were assigned as follows: 1730 cm-1 (unconjugated C = O in hemicelluloses), 1645 cm-1 (absorbed O-H in cellulose), 1500-1550 cm-1 (aromatic rings of lignin), 1458 and 1428 cm-1 (C-H deformation in cellulose and hemicelluloses), 1265 cm-1 (C-H stretching in lignin), 1117 and 1060 cm-1 (C-O stretching in cellulose and hemicelluloses) and 877 cm-1 (C-H deformation in cellulose) (Faix, 1992; Pandey, 1999; Pandey and Pitman, 2003, 2004; Schwanninger et al., 2004; Genestar and Palou, 2006; Rana et al., 2010).
For Shorea wood decayed by monokaryons (Fig. 2b), a peak showed the broader signal at~3500 cm-1 (peak 1), indicating that the sample wood powder had water vapour. It is generally known that the region between 1800 and 600 cm-1 is a fingerprint region for wood. The fingerprint region 1 corresponding signal at 1710-1740 cm-1 (peak number 2) indicated for the unconjugated C = O in hemicelluloses. The fingerprint in region 2 signals at 1500-1600 cm-1 corresponding to peak numbers 3 and 4 were changed when compared to those of the undecayed Shorea wood. The peak numbers 3 and 4 showed that lignin was degraded, particularly at a peak around 1510 cm-1 that is frequently used as a reference band of lignin changes (Berben et al., 1987; Friese and Banerjee, 1992). The signals in fingerprint region 3 at 1400-1060 cm-1 corresponding to peak numbers 5, 6 and 7, fingerprint region 4 at 600-850 cm-1 corresponding to peak numbers 8, 9, 10 and 11 were found to be different from those of undecayed Shorea wood. These results revealed that cellulose and hemicelluloses polymers of wood cell wall were degraded.
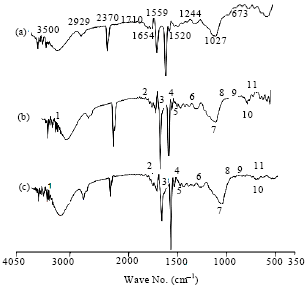 | |
| Fig. 2: | FTIR spectra of (a) Undecayed Shorea wood blocks (b) Wood blocks decayed by monokaryon and (c) Wood blocks decayed by dikaryon, after 12 weeks of incubation |
The contents of the hemicelluloses and cellulose polymers could be calculated from the carbohydrate composition of the wood by assuming that the glucose/mannose ratio was 1:2 for hardwood glucomannan (1 mol of Glc/2 mol of Man).
Wave numbers at 814 cm-1, 870 cm-1, 872 cm-1 and 898 cm-1 are correlated with glucomannan, mannose, glucomannan and cellulose, respectively (Kacurakova et al., 2000; Bjarmestad and Dahlman, 2002). The results showed that signals at 800 to 900 cm-1 were slightly changed and it was possible that hemicellulose and cellulose polymers were partially degraded even though Cambodian P. linteus is a white rot fungus.
For Shorea wood decayed by dikaryons (Fig. 2c) the fingerprint regions 1, 2, 3 and 4 were changed. Many peaks (numbers 1 to 11) in each fingerprint region decreased, but the intensity of some peaks increased. The results indicated that the lignin in Shorea wood, which is a natural phenolic compound was majorly degraded. Lignin polymers were broken down due to the influence of the oxidative enzymes, such as manganese-peroxidase, lignin peroxidase and laccase, which are produced by dikaryons during their growth and formation of initial fruiting bodies. In previous study, Cambodian P. linteus also produced a high amount of laccase, stimulated by copper (Cu2) (Srivilai, 2006).
Interestingly, the fingerprint regions 1, 2, 3 and 4 of the Shorea wood decayed by dikaryons were similar to those of Shorea wood decayed by monokaryons. This result suggested that monokaryons and dikaryons may secrete the same set of hydrolytic and oxidative enzymes during fungal incubation. These enzymes caused the depolymerisation of lignin, cellulose and hemicelluloses polymers, therefore simple sugar molecules were produced. However, some peaks in fingerprint region 4 contained low signals, compared to the same peaks in Shorea wood decayed by monokaryons. Interestingly, by a white rot fungus (Phanerochaete velutina) cellulose and hemicelluloses were degraded at the initial stage of wood degradation, i.e., 30 days after incubation. In contrast, Stropharia rugosoannulata attacked cellulose but never exceeded the loss of hemicelluloses. However, these fungi possibly produced predominant enzymes namely, cellulase and manganese peroxidase. In general, lignin is a major natural polyphenolic compound in wood, comprising about 20-30% of the dry mass. It is mostly deposited in wood cell corners (Boerjan et al., 2003; Gierlinger et al., 2004). In this study, FTIR analysis revealed that the lignin, cellulose and hemicelluloses were depolymerised due to changes in FTIR peaks compared to the FTIR reference bands (Parker, 1983; Faix et al., 1993; Bjarmestad and Dahlman, 2002; Ferraz et al., 2000; Pandey and Pitman, 2003, 2004). During the period when the hyphae attacked the wood, different types of hydrolytic enzymes could be liberated and alter the complex cell wall components to become fungal energy sources. Pandey and Pitman (2003) studied the chemical changes in beech wood decayed by C. versicolor using FTIR and found that it decreased the intensities of lignin and carbohydrate peaks, preferentially hemicelluloses (Pandey and Pitman, 2003, 2004) (Fig. 2).
Observation of wood structural changes by SEM: Shorea wood blocks decayed by monokaryons and dikaryons were examined for wood structural changes by SEM. Each Shorea wood block sample was longitudinal and cross sectioned and the results showed that undecayed Shorea wood blocks were not changed and the Shorea cell walls were still intact. The outer layer of Shorea wood blocks remained smooth, some wood pits were present and wood cell structures were complete. Shorea wood block samples that were decayed by monokaryons showed that the wood layers were changed. The longitudinal and cross section views (Fig. 3c, d) showed that inside of the wood tracheids there were numerous hyphae-deposited and the tracheid structures were changed when compared to the undecayed wood tracheids (Fig. 3b). Also found that Shorea cell wall structures were decayed. According to several studies, white rot fungi usually attack all the wood components simultaneously (Fackler et al., 2007). In fact, white rot fungi preferentially remove lignin from the entire cell wall causing the separation of cells, especially when the middle lamella is extensively attacked. It has been shown that white rot fungi were colonized on wood quickly and the ray parenchyma cells are the first to be colonized. This observation revealed that fungal hyphae penetrate from cell to cell via pit structures or bore holes through the cell walls. Moreover, several factors influenced the wood degradation pattern, for example, wood nitrogen content, temperature and humidity. However, the present study also showed the changes in cell wall structures of wood that were decayed by both monokaryons and dikaryons (Fig. 3). Unfortunately, the differences were not very obvious in this study. For further study, the biodegradation time could be extended in order to differentiate between undecayed and decayed wood structures.
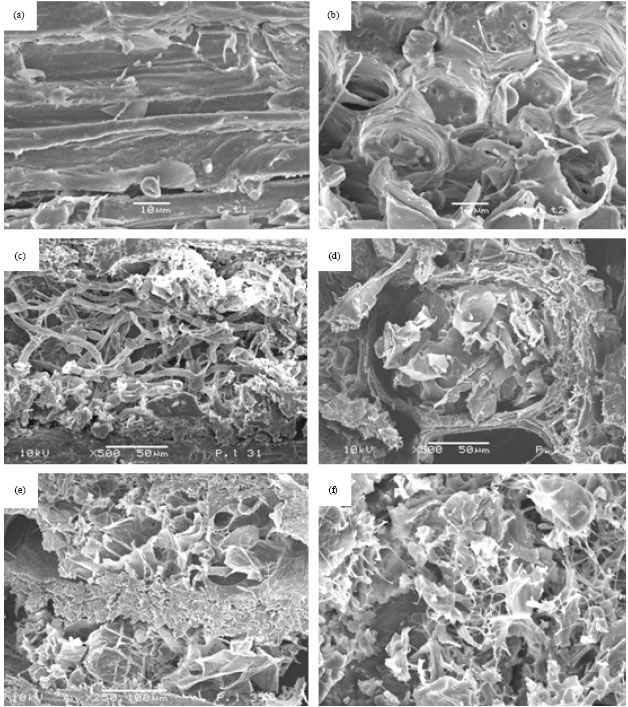 | |
| Fig. 3(a-f): | Undecayed Shorea wood structure was not changed and the wood layer was smooth (a-b), whereas the hyphae penetrated the decayed Shorea wood blocks and wood layers and changed their structures (c) and (d) show monokaryotic mycelium penetrating tracheids. (e) and (f) show dikaryotic mycelium penetrating tracheids and decaying Shorea cell wall structures |
CONCLUSION
Cambodian P. linteus is a white rot fungus and used for the examination of Shorea wood degradation. This fungus secreted hydrolytic enzymes to decay wood components such as lignin, cellulose and hemicellulose. Monokaryons and dikaryons have slightly different patterns in Shorea wood degradation. However, the differences were not statistically significant. Cambodian P. linteus monokaryons were a more appropriate mycelium for wood degradation, some paper industrial processes and enzyme technology, due to its slowly growing and easy management. Moreover, Cambodian P. linteus monokaryons did not produced dark pigments from the fruiting body.
ACKNOWLEDGMENTS
This study was supported by a grant from Mahasarakham University, Thailand. Gracious thanks is extended to Ananyapron Prommettha for laboratory assistance and Department of Biology for laboratory space and special thanks to Dr. Jolyon Dodgson for the proof editing of English manuscript.
REFERENCES
- Addleman, K. and F. Archibald, 1993. Kraft pulp bleaching and delignification by dikaryons and monokaryons of Trametes versicolor. Applied Environ. Microbiol., 59: 266-273.
Direct Link - Akio, E., T. Hiromi and F. Goro, 1988. Degradation of lignin-related compounds, pure cellulose, and wood components by white-rot and brown-rot Fungi. Holzforschung, 42: 85-93.
Direct Link - Berben, S.A., J.P. Rademacher, L.O. Sell and D.B. Easty, 1987. Estimation of lignin in wood pulp by diffuse reflectance Fourier-transform infrared spectrometry. Taipe J., 70: 129-140.
Direct Link - Blanchette, R.A., L. Otjen, M.J. Effland and W.E. Eslyn, 1985. Changes in structural and chemical components of wood delignified by fungi. Wood Sci. Technol., 19: 35-46.
CrossRef - Bjarmestad, S. and O. Dahlman, 2002. Chemical composition of hard wood and softwood pulps employing photoacoustic fourier transform infrared spectroscopy in combination with partial least squares analysis. Anal. Chem., 74: 5851-5858.
PubMedDirect Link - Breen, A. and F.L. Singleton, 1999. Fungi in lignocellulose breakdown and biopulping. Curr. Opin. Biotechnol., 10: 252-258.
CrossRefDirect Link - Boerjan, W., J. Ralph and M. Baucher, 2003. Lignin biosynthesis. Ann. Rev. Plant Biol., 54: 519-546.
CrossRefDirect Link - Fackler, K., M. Schwanninger, C. Gradinge, B. Hinterstoisser and K. Messne, 2007. Qualitative and quantitative changes of beech wood degraded by wood-rotting basidiomycetes monitored by Fourier transform infrared spectroscopic methods and multivariate data analysis. FEMS Microbiol. Lett., 271: 162-169.
CrossRefDirect Link - Faix, O., J. Bremer, O. Schmidt and S.J. Tatjana, 1993. Monitoring of chemical changes in white-rot degraded beech wood by pyrolysis-gas chromatography and Fourier-transform infrared spectroscopy. J. Anal. Applied Pyrol., 21: 147-162.
CrossRef - Fischer, M and M. Binder, 2004. Species recognition, geographic distribution and host-pathogen relationships: A case study in a group of lignicolous basidiomycetes, Phellinus s.l. Mycologia, 96: 799-811.
Direct Link - Friese, M.A and S. Banerjee, 1992. Lignin determination by FT-IR. Applied Spectrosc., 46: 246-248.
Direct Link - Ferraz, A., J. Baeza, J. Rodriguez and J. Freer, 2000. Estimating the chemical composition of biodegraded pine and eucalyptus wood by DRIFT spectroscopy and multivariate analysis. Bioresour. Technol., 74: 201-212.
CrossRef - Genestar, C. and J. Palou, 2006. SEM-FTIR spectroscopic evaluation of deterioration in an historic coffered ceiling. Anal. Bioanal. Chem., 384: 987-993.
CrossRef - Gierlinger, N., D. Jacques, M. Schwanninger, R. Wimmer and L.E. Paques, 2004. Heartwood extractives and lignin content of different larch species (Larix sp.) and relationships to brown-rot decay-resistance. Trees, 18: 230-236.
CrossRef - Kacurakova, M., P. Capek, V. Sasinkova, N. Wellner and A Ebringerova, 2000. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym., 43: 195-203.
CrossRef - Kothe, E., 2001. Mating-type genes for basidiomycete strain improvement in mushroom farming. Applied Microbiol. Biotechnol., 56: 602-612.
CrossRef - Kues, U., 2000. Life history and developmental processes in the basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev., 64: 316-353.
PubMedDirect Link - Pandey, K.K., 1999. A study of chemical structure of soft and hardwood and wood polymers by FTIR spectroscopy. J. Applied Polym. Sci., 71: 1969-1975.
CrossRefDirect Link - Pandey, K.K. and A.J. Pitman, 2003. FTIR studies of the changes in wood chemistry following Decay by brown-rot and white-rot fungi. Int. Biodeterior. Biodegrad., 52: 151-160.
CrossRef - Pandey, K.K. and A.J. Pitman, 2004. Examination of the lignin content in a softwood and a hardwood decayed by a brown-rot fungus with the acetyl bromide method and Fourier transform infrared spectroscopy. J. Polym. Sci. A, 42: 2340-2346.
CrossRef - Pasikatan, M.C., J.L. Steele, C.K. Spillman and E. Haque, 2001. Near infrared reflectance spectroscopy for online particle size analysis of powders and ground materials. J. Near Infrared Spectrosc., 9: 153-164.
Direct Link - Raberg, U. and G. Daniel, 2009. Brown rot decay of copper-chromated-phosphorus impregnated fence poles: Characterization by molecular analyses and microscopy. Int. Biodeterior. Biodegrad., 63: 906-912.
CrossRef - Rana, R., R. Langgenfeld-Heyser, R. Finkeldey and A. Polle, 2010. FTIR spectroscopy, chemical and histochemical characterisation of wood and lignin of five tropical timber wood species of the family of Dipterocarpaceae. Wood Sci. Technol., 44: 225-242.
CrossRef - Rodrigues, J., O. Faix and H. Pereira, 1998. Deterimation of lignin content of Eucalytus globules wood using FTIR spectrometry. Holzforschung, 52: 46-50.
Direct Link - Schwanninger, M., J. Rodrigues, H. Pereira abnd B. Hinterstoisser, 2004. Effects of short-time vibratory ball milling on the shape of FT-IR spectra of wood and cellulose. Vib Spetrosc., 36: 23-40.
CrossRef - Worrall, J.J., S.E. Anagnost and R.A. Zabel, 1997. Comparison of wood decay among diverse Lignicolous fungi. Mycologia, 89: 199-219.
Direct Link