
Giek Far Chan
Nanoporous Materials for Biological Applications Research Group, Sustainability Research Alliance, Universiti Teknologi Malaysia, 81310 UTM Johor Bahru, Johor, Malaysia
Noor Aini Abdul Rashid
Nanoporous Materials for Biological Applications Research Group, Sustainability Research Alliance, Universiti Teknologi Malaysia, 81310 UTM Johor Bahru, Johor, Malaysia
Abdull Rahim Mohd Yusoff
Department of Chemistry, Faculty of Science, Universiti Teknologi Malaysia, 81310 UTM Johor Bahru, Johor, Malaysia
Lee Suan Chua
Institute of Bioproduct Development, Universiti Teknologi Malaysia, 81310 UTM Johor Bahru, Johor, Malaysia
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 2 | Page No.: 91-97







ABSTRACT
In this study, we attempted to understand possible mechanism associated with bacterial azo dye decolourisation via mass spectral analysis and shotgun cloning of genomic DNA from C. freundii A1. The presence of S-ribosylhomocysteine, a precursor in biosynthesis of autoinducer-2 after complete decolourisation of Amaranth shed light on quorum sensing as the possible related mechanism. Screening of recombinants from E. coli strain ABLE C library on plates supplemented with Amaranth dye resulted in the isolation of a positive recombinant clone, designated ABLE C-pTrueBlue-ropA12 (or in short AC-A12) which showed enhanced Amaranth decolourisation activity. Molecular identification of the cloned DNA fragment revealed an autoinducer-2 synthase (luxS) gene that codes for the final enzyme in the biosynthetic pathway for production of autoinducer-2, which involves in bacterial quorum sensing mechanism. This is the first time the biosynthesis of autoinducer is reported to be related to azo dye decolourisation.
PDF Abstract XML References Citation
Received: November 22, 2011;
Accepted: January 04, 2012;
Published: March 13, 2012
How to cite this article
Giek Far Chan, Noor Aini Abdul Rashid, Abdull Rahim Mohd Yusoff and Lee Suan Chua, 2012. Biosynthesis of Autoinducer-2 as the Possible Mechanism to Enhance Decolourisation of Azo Dye by Citrobacter freundii A1. Journal of Biological Sciences, 12: 91-97.
DOI: 10.3923/jbs.2012.91.97
URL: https://scialert.net/abstract/?doi=jbs.2012.91.97
DOI: 10.3923/jbs.2012.91.97
URL: https://scialert.net/abstract/?doi=jbs.2012.91.97
INTRODUCTION
Bioremediation of azo dyes remains as a critical industrial and environmental issue in the 21st century. Azo dyes, characterized by the presence of azo bonds (-N = N-), remain as the largest group, 60-70% of commercially produced synthetic dyes that do not occur naturally (Stolz, 2001; Omar, 2008). The resistance of azo dyes to biological or chemical degradation is due to the azo linkages, the aromatic moieties and sulfonated substitutions that resulted in colour fastness and structural stability (Saratale et al., 2011). The lack of proper degradation of dyes presents other environmental hazard as the dyes and their metabolites may leach into the water bodies and pose health hazard (Srikanlayanukul et al., 2008).
Generally, mechanisms involved in microbial decolourisation of azo dyes include adsorption, transformation or metabolism of dye compounds. Over the decades of research, bacterial decolourisation of azo dyes has gained progress as treatment method, though the study on mechanism of bacterial decolourisation remains lacking. Numerous studies mainly focused on the enzymatic aspect of bacterial decolourisation. For instances, Hong et al. (2009) presented evidence of dissimilatory azoreduction at the subcell level from Shewanella decolorationis S12. Russ et al. (2000) suggested that the reduction of sulfonated azo dyes by reduced flavins formed by cytosolic flavin-dependent azoreductases. This was observed both in vitro and in vivo and was evident with the recombinant strain Sphingomonas sp. BN6(pRJR34), where the in vitro reaction showed significant increase in azoreductase activities as compared to the wild type strain (Russ et al., 2000). As reviewed by Saratale et al. (2011), significant research were done on both oxidative and reductive enzymes and published at the microbiological, protein and molecular level that contributed significantly to the understanding of decolourisation mechanism.
In this study, the presence of a possible mechanism which could enhance decolourisation of azo dyes by C. freundii A1 was investigated. The strain was formerly isolated from the oxidation pond in the vicinity of Universiti Teknologi Malaysia, Johor, Malaysia and is capable of decolourising azo dyes (Rashid et al., 1999). Ina previous study done. C. freundii A1 was shown to possess flavin reductase which catalyzed reductive cleavage of Amaranth. Hence, in this study, mass spectral analysis and shotgun cloning were carried out to gain further insight on other mechanisms related to decolourisation. The presence of autoinducer-2 precursor, as well as, the molecular identification and decolourisation activity of recombinant containing autoinducer-2 synthase gene which showed enhanced azo dye decolourisation is reported here. Quorum sensing was hypothesized as the possible mechanism involved in decolourisation of azo dyes by C. freundii A1. In silico bioinformatic analyses revealed some novel understanding of the possible interaction of autoinducer-2 synthase in quorum sensing and biodegradation metabolic pathway.
MATERIALS AND METHODS
Microorganism: C. freundii A1 was obtained from the culture collection of the Laboratory of Molecular Biology and Microbiology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia. The bacterial strain was maintained on nutrient agar at 4°C or as a 15% (v/v) glycerol stock for long term-storage at -80°C.
Decolourisation of Amaranth by C. freundii A1: Amaranth was chosen as the model azo dye and used at a final concentration of 0.1 g L-1 for decolourisation study. P5 medium was prepared and decolourisation of Amaranth by C. freundii A1 was carried out under microaerophilic condition according to the procedures described by Chan et al. (2011). Complete decolourisation was achieved within 24 h followed by centrifugation at 5,000 rpm for 15 min. The supernatant was collected and transferred to a separating funnel. Extraction was done with equal volume of ethyl acetate. The organic layer was separated and concentrated prior to LC-MS/MS.
LC-MS/MS analysis of autoinducer-2 precursors: Acquity analytical UPLC (Waters, USA) equipped with a C18 reserved phase Acquity column (150 x 4.6 x 1.7 μm; Waters, USA) was used. The mobile phase was a binary solvent system consisting of 0.1% formic acid and 100% acetonitrile. The eluent flow rate was set at 0.25 mL min-1 and the injection volume was 5 μL. The acetonitrile concentration was started at 10% for 5 min and ramped up to 90% over 10 min and held constant for 5 min. The acetonitrile concentration was ramped down to 10% in the next 5 min, followed by 5 min of final washing and re-conditioning of the column. An Electrospray Ionization (ESI) and a triple-quadrupole-linear ion trap of tandem mass spectrometer, 4000 Q TRAP (Applied Biosystems, Canada) in positive ion mode with a mass range up to m/z 700 was used for mass detection. The scan rate was set at 1000 amu s-1. The source temperature was set at 400°C. Nitrogen was used as ion source gas for nebulizing, 40 psi; drying solvent, 40 psi; curtain gas, 10 psi; collision gas, high; declustering potential, -40 V and collision exit energy, -10 V. Data acquisition and processing was done using Analyst 1.4.2, followed by prediction of autoinducer-2 precursors which was carried out using MS Fragmenter 12.0 (ACD/Labs, Canada).
Shotgun cloning of genomic DNA from C. freundii A1: Genomic DNA isolation was performed using Wizard Genomic DNA Purification Kit (Promega) according to manufacturer’s instruction. The isolated genomic DNA was viewed under UV transillumination in 1% (w/v) agarose. Genomic DNA from C. freundii A1 was partially digested for 1 h using restriction endonuclease BglII (Fermentas, Lithuania) and the reaction was stopped by adding EDTA (6 μL; 0.25 M). The DNA fragments of sizes between 3 kb to 10 kb were carefully excised from 1% (w/v) agarose gel and extracted using PURE-GENE Gel Extraction Kit (BST Techlab, Malaysia). The DNA fragments were cloned into pTrueBlue-rop vector (Genomics One, Canada) which was digested using restriction endonuclease BglII and dephosphorylated with calf intestine alkaline phosphatase (Fermentas, Lithuania). The linearized plasmid was also extracted from 1% (w/v) agarose gel. Ligation was carried out at the vector: insert ratio of 1:2. The ligation reaction was performed overnight at 8-10°C using T4 DNA ligase (Fermentas, Lithuania). The resulting ligation mixture was used for bacterial transformation into competent Escherichia coli ABLE C; lac(LacZω-) [Kanr McrA- McrCB- McrF- Mrr- HsdR (rK- mK-)] [F’ proAB lacIqZΔM15 Tn10 (Tetr)] (Stratagene, USA). The cells were plated out for blue-white screening. The recombinant clones were isolated and maintained on Luria-Bertani (LB) agar containing 100 μg mL-1 of ampicillin.
Screening for dye-decolourising recombinants: LB agar plates containing Amaranth azo dye (0.1 g L-1), IPTG (0.1 mM) and ampicillin (100 μg mL-1) were prepared. Recombinant clones were streaked on the LB plates and incubated at 37°C for 3 to 4 days. Colonies that could decolourise were selected and screening was repeated in BHI broth (Oxoid, Australia). When necessary, ampicillin (100 μg mL-1), IPTG (0.5 mM) and Amaranth azo dye (0.1 g L-1) were added to the media. Inoculum of E. coli recombinant AC-A12 was prepared in LB broth containing ampicillin (100 μg mL-1) and optical density was determined at 550 nm. Similar amount of E. coli ABLE C containing pTrueBlue-rop was used as control. The inoculum was added into BHI broth containing ampicillin (100 μg mL-1) and grown aerobically at 37°C for 18 h, followed by static incubation to enable decolourisation of azo dye under reducing environment.
Analyses of dye-decolourising recombinant: Recombinant plasmid from dye-decolourising clone was isolated and digested with restriction endonuclease BglII. An aliquot was electrophoresed through 1% (w/v) agarose gel. Sequencing of the unknown insert was attempted using T7 universal primers. The sequence obtained was compared with the known DNA and amino acid sequences in GenBank database at NCBI (http://www.ncbi.nlm.nih.gov/) using BLASTn and BLASTx (Altschul et al., 1997). Multiple sequence alignment with related sequences was performed using ClustalW. MEGA version 4.1 (Beta 3) was used for construction of Neighbor-Joining phylogenetic tree with bootstrap values calculated based on 1000 replicates (Tamura et al., 2007). In silico analysis was carried out at ExPASy server (http://expasy.org/tools/) and prediction of protein interaction was done using STRING 9.0 at http://string-db.org (Gasteiger et al., 2005; Szklarczyk et al., 2011).
RESULTS
After complete decolourisation of Amaranth within 24 h, the decolourised sample was extracted and analyzed by LC-MS/MS. Figure 1 shows the mass spectra of S-ribosylhomocysteine (SRH) with m/z ratio of 266.2 detected at retention time of 13.2 min. The analysis revealed the presence of precursor for biosynthesis of autoinducer-2. This discovery led to the belief that quorum sensing could be related to bacterial decolourisation.
Hence, genomic library was constructed and the library produced 2.5 x 106 clones. Screening of 500 clones on LB plates containing 0.1 g L-1 Amaranth allowed the selection of 12 yellowish-coloured colonies which may indicate the cells’ capacity to decolourise the red colour Amaranth adsorbed from its surrounding. The clones were further screened for decolourisation activity in BHI broth. Figure 2 shows the decolourisation capability of one of the E. coli ABLE C recombinant, designated AC-A12 which exhibited the best decolourisation activity. The clone was able to decolourise Amaranth at faster rate compared to the control used at similar amount of cells.
The recombinant plasmid from AC-A12 was isolated and digested with BglII to release the DNA insert (Fig. 3). After restriction, two fragments giving a total size of 7.5 kb were observed; 3 and 4.5 kb. Therefore, the gene cassette possessed an internal restriction site for BglII. The 7.5-kb fragment was further sequenced and analyzed. The fragment was found to possess the full sequence of an autoinducer-2 synthase (luxS) gene. The 456-bp gene sequence was deposited into GenBank database under accession number AY646883.1.
The luxS gene from C. freundii A1 showed 97% identity with autoinducer-2 synthase gene from Clostridium difficile.
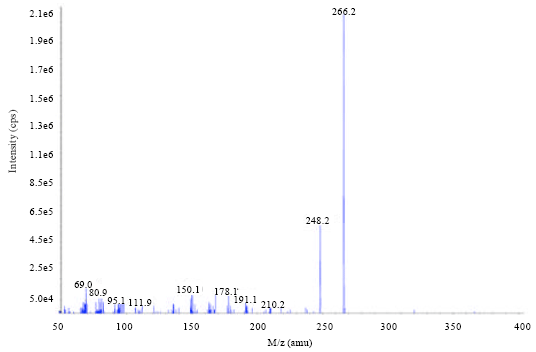 | |
| Fig. 1: | Mass spectral analysis of S-ribosylhomocysteine (m/z: 266.2) detected after 24 h of Amaranth decolourisation by C. freundii A1 |
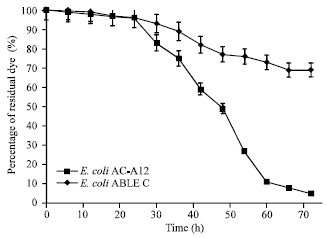 | |
| Fig. 2: | Decolourisation of Amaranth (0.1 g L-1) by E. coli AC-A12 clone in BHI broth. E. coli ABLE C transformed with pTrueBlue-rop plasmid was used as control |
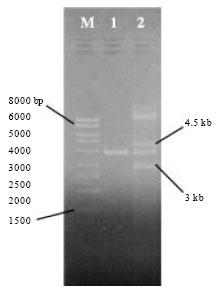 | |
| Fig. 3: | Recombinant plasmid from E. coli ABLE C recombinant (AC-A12 clone) after digestion with BglII. Lane 2 shows the 7.5-kb gene cassette (fragments of 4.5 kb and 3 kb in size) from recombinant plasmid of AC-A12, which is compared to Lane 1 of the pTrueBlue-rop vector (3.757 kb). Lane M is the Mass Ruler™ DNA ladder (Fermentas) |
From codon usage analysis for this autoinducer-2 synthase, there is an inclination for codon selection, for example UUA for leucine, AGA for arginine, GGA for glycine, GUA for valine, ACA for threonine, AAA for lysine, UUU for phenylalanine, GAA for glutamic acid and GAU for aspartic acid. The gene contained a total number of nucleotides of 163 Adenine, 55 Cytosine, 98 Guanine and 137 Thymine. The polypeptide contains 151 amino acid residues. It also revealed 96% homology to the amino acid sequence of S-ribosylhomocysteinase from Clostridium difficile. LuxS from C. freundii A1 had an estimated molecular weight (MW) of 16.8 k Da and a predicted pI of 5.66. The multiple sequence alignment of LuxS amino acid sequences from C. freundii A1 and other bacteria obtained from GenBank was performed and a phylogenetic tree was generated to determine that homology of autoinducer-2 synthase that exists among different bacteria (Fig. 4). The phylogram reveals the homologous relationship of the autoinducer-2 synthase from C. freundii A1 with other bacteria, particularly to Clostridium spp. The protein appeared to be quite conserved across species of bacteria. The autoinducer-2 synthase from C. freundii A1 belongs to LuxS superfamily comprising of the LuxS protein involved in synthesis of autoinducer AI-2 and its hypothetical relatives.
In addition, in silico analysis was carried out to predict the function and interaction of autoinducer-2-synthase by comparing with a related Citrobacter sp. Fig. 5 shows that autoinducer-2-synthase mainly interacts with nucleoside phosphorylase, cystathionine beta-lyases/cystathionine gamma-synthases, cysteine synthase, methionine synthase II (cobalamin-independent), 5,10-methylenetetrahydrofolate reductase and HAD (haloacid dehalogenase) superfamily sugar phosphatases. The protein-protein interactions with STRING 9.0 were provided with confidence score of 0.899 to 0.999.
DISCUSSION
In this study, the presence of precursor S-ribosylhomocysteine (SRH) for biosynthesis of autoinducer-2 (AI-2) was elucidated in decolourised sample of C. freundii A1. Autoinducer-2 has been suggested to act as ‘universal’ signal molecules for interspecies communication and encodes information specific to cell number (Xavier and Bassler, 2003). In the biosynthesis of AI-2, autoinducer-2-synthase (LuxS) catalyzes the transformation of S-ribosylhomocysteine (SRH) into 4, 5-dihydroxy-2, 3-pentanedione (DPD) and homocysteine. DPD undergoes thermodynamically favorable cyclization into autoinducer molecules of 2, 4-dihydroxy-2-methyldihydro-3-furanone (DHMF) (De Keersmaecker et al., 2006). Winzer et al. (2002) reported the presence of LuxS as the N-ribosylhomocysteinase (cleavage enzyme) in C. freundii and C. intermedius by sequence alignment with various bacteria possessing pathway for the synthesis of AI-2.
 | |
| Fig. 4: | Phylogram generated based on the amino acid sequence alignment of autoinducer-2 synthase from C. freundii A1 with other bacteria. The GenBank assession numbers of the autoinducer-2 synthase amino acid sequences used are indicated in the parentheses. |
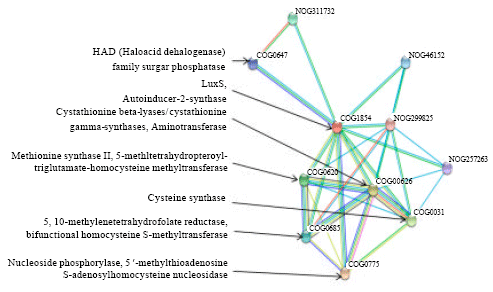 | |
| Fig. 5: | Predictive protein-protein interaction of autoinducer-2-synthase (LuxS protein; COG1854) from related C. koseri with its functional protein partners by STRING 9.0. (COG0775: Nucleoside phosphorylase; COG0626: cystathionine beta-lyases/cystathionine gamma-synthases; COG0031: cysteine synthase; COG0620: methionine synthase II (cobalamin-independent); COG0685: 5,10-methylenetetrahydrofolate reductase; COG0647: predicted sugar phosphatases of the HAD superfamily; NOG311732, NOG46152, NOG257263 and NOG299825: non supervised orthologous group) |
In this study, C. freundii A1 was shown to possess the gene coding for autoinducer-2-synthase which could function in the catalytic transformation of SRH to AI-2. To the best of our knowledge, this is for the first time that biosynthesis of AI-2 was associated with decolourisation of azo dye. The physiological roles of AI-2 in various microorganisms could be varied and study in this area remains limited to biofilm formation via quorum sensing mechanism (Komiya-Ito et al., 2006). The isolation of recombinant E. coli AC-A12 with enhanced dye-decolourizing capability suggests a relationship between decolourisation and AI-2 synthesis. The induction of this gene in recombinant E. coli AC-A12 resulted in faster decolourisation of azo dye. The recombinant clone or C. freundii A1 may utilize quorum sensing mechanism to regulate expression of genes under specific conditions, such as under nutrient-limited condition, toxic surrounding and for biodegradation of azo reduction products. DeLisa et al. (2001) attempted on DNA microarray based identification to investigate the global changes in transcription and mRNA abundance elicited by the AI-2 signaling molecule in E. coli. The study suggested a significant induction of genes related to quorum sensing (DeLisa et al., 2001). Barrios et al. (2006) further elucidated the AI-2 induced biofilm formation in E. coli through the motility quorum-sensing regulator gene.
The in silico analysis revealed the functional interaction of autoinducer-2-synthase with enzymes related to quorum sensing and biodegradation. The aminotransferase classes of enzymes are hypothesized to be involved in deamination of amine compounds, which are dye intermediates formed after reduction of azo bonds during decolourisation of azo dyes. Deamination by aminotransferase could be the intial step to the biotransformation of amine compounds, followed by desulfonation pathway. Magony (2004) found that an aminotransferase type and ring hydroxylating dioxygenase enzymes, probably participated in the biotransformation of sulfanilic acid (or p-amino-benzenesulfonate, which is a typical representative of aromatic sulfonated amines), to sulfocatechol by Sphingomonas subarctica (Magony, 2004). This interaction could be further explored to understand the function of aminotransferase in biotransformation of aromatic amines resulted from azo dye decolourisation and its relations in microbial quorum sensing. Besides that, autoinducer-2-synthase (LuxS) was predicted to interact with HAD (haloacid dehalogenase) family sugar phosphatase (Fig. 5). The HAD superfamily functions in microbial degradation of chlorinated pollutants. This superfamily consists of more than 3000 members in both prokaryotic and eukaryotic organisms which includes dehalogenases, phosphoesterases, ATPases, phosphonatases and sugar phosphomutases (Huang et al., 2011; Lu et al., 2005). However, the vast majority of HAD superfamily members are thought to function as phosphatases (phosphohydrolases) (Nguyen et al., 2010). This finding is interesting as there is no report that relates quorum sensing mechanism with dehalogenation in bioremediation of chlorinated pollutants in soil. The overall mechanism of LuxS related to quorum sensing and biodegradation, as well as transcriptional changes in microbes during decolourisation of azo dye, would be an interesting aspect for future study of transcriptome profiling.
CONCLUSIONS
In this study, the presence of a precursor for the biosynthesis of autoinducer-2 and autoinducer-2 synthase gene in C. freundii A1 was elucidated. The isolation of recombinant AC-A12 which showed dye removal capability points to the possible involvement of autoinducer-2 synthesis in decolourisation of azo dyes. In silico analyses revealed the possible interactions of this enzyme in quorum sensing and biodegradation pathways.
ACKNOWLEDGMENTS
We would like to thank the Ministry of Science, Technology and Innovation (MOSTI), Malaysia, for financial support to carry out this research.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Barrios, A.F.G., R. Zuo, Y. Hashimoto, L. Yang, W.E. Bentley and T.K. Wood, 2006. Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022). J. Bacteriol., 188: 305-316.
CrossRefDirect Link - Chan, G.F., N.A.A. Rashid, L.L. Koay, S.Y. Chang and W.L. Tan, 2011. Identification and optimization of novel NAR-1 bacterial consortium for the biodegradation of Orange II Insight Biotechnol., 1: 7-16.
CrossRef - De Keersmaecker, S.C.J., K. Sonck and J. Vanderleyden, 2006. . Let LuxS speak up in AI-2 signaling. Trends Microbiol., 14: 114-119.
CrossRef - DeLisa, M.P., C-F. Wu, L. Wang, J.J. Valdes and W.E. Bentley, 2001. DNA microarray-based identification of genes controlled by autoinducer 2-stimulated quorum sensing in Escherichia coli. J. Bacteriol., 183: 5239-5247.
CrossRef - Gasteiger, E., C. Hoogland, A. Gattiker, S. Duvaud, M.R. Wilkins, R.D. Appel and A. Bairoch, 2005. Protein Identification and Analysis Tools on the ExPASy Server. In: The Proteomics Protocols Handbook, Walker, J.M. (Ed.). 1st Edn., Humana Press, New Jersey, USA., ISBN-13: 978-1588295934, pp: 571-607.
- Hong, Y.G., J. Guo and G.P. Sun, 2009. Energy generation coupled to azoreduction by membranous vesicles from Shewanella decolorationis S12. J. Microbiol. Biotechnol., 19: 37-41.
PubMedDirect Link - Huang, J., Y. Xin and W. Zhang, 2011. Isolation, characterization and identification of a Paracoccus sp. 2-haloacid-degrading bacterium from the marine sponge Hymeniacidon perlevis. J. Basic Microbiol., 51: 318-324.
CrossRef - Lu, Z., D. Dunaway-Mariano and K.N. Allen, 2005. HAD Superfamily phosphotransferase substrate diversification: Structure and function analysis of HAD subclass IIB sugar phosphatase BT4131. Biochemistry, 44: 8684-8696.
CrossRef - Magony, M., 2004. Characterization of the biodegradation pathways of sulfanilic acid and catechol derivatives in Sphingomonas subarctica. Acta Biologica Szegediensis, 48: 65-65.
Direct Link - Nguyen, H., L. Wang, H. Huang, E. Peisach, D. Dunaway-Mariano and K.N. Allen, 2010. Structural determinants of substrate recognition in the HAD superfamily member D-glycero-D-manno-heptose 1,7- bisphosphate phosphatase, GmhB. Biochemistry, 49: 1082-1092.
CrossRef - Russ, R., J. Rau and A. Stolz, 2000. The function of cytoplasmic flavin reductases in the reduction of azo dyes by bacteria. Applied Environ. Microbiol., 66: 1429-1434.
PubMedDirect Link - Saratale, R.G., G.D. Saratale, J.S. Chang and S.P. Govindwar, 2011. Bacterial decolorization and degradation of azo dyes: A review. J. Taiwan Inst. Chem. Eng., 42: 138-157.
CrossRefDirect Link - Stolz, A., 2001. Basic and applied aspects in the microbial degradation of azo dyes. Applied Microbiol. Biotechnol., 56: 69-80.
CrossRefDirect Link - Szklarczyk, D., A. Franceschini, M. Kuhn, M. Simonovic and A. Roth et al., 2011. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res., 39: D561-D568.
CrossRef - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Winzer, K., K.R. Hardie, N. Burgess, N. Doherty and D. Kirke et al., 2002. LuxS: its role in central metabolism and the In vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology, 148: 909-922.
Direct Link - Xavier, K.B. and B.L. Bassler, 2003. LuxS quorum sensing: More than just a numbers game. Curr. Opin. Microbiol., 6: 191-197.
CrossRefDirect Link - Omar, H.H., 2008. Algal decolorization and degradation of monoazo and diazo dyes. Pak. J. Biol. Sci., 11: 1310-1316.
CrossRefPubMedDirect Link - Srikanlayanukul, M., K. Wirut, W. Takashi and K. Chartchai, 2008. Decolorization of orange ii by immobilized thermotolerant white rot fungus Coriolus versicolor RC3 in packed-bed bioreactor. Biotechnology, 7: 280-286.
CrossRefDirect Link - Komiya-Ito, A., T. Ito, A. Yamanaka, K. Okuda, S. Yamada and T. Kato, 2006. N-tetradecanoyl homoserine lactone, signaling compound for quorum sensing, inhibits Porphyromonas gingivalis growth. Res. J. Microbiol., 1: 353-359.
CrossRefDirect Link